草地贪夜蛾高致病力绿僵菌菌株的筛选与鉴定
2023-10-20赵建伟丁雪玲赵金凤姚凤銮卢学松何玉仙
赵建伟,丁雪玲,郑 宇,赵金凤,姚凤銮,卢学松,何玉仙*
(1. 福建省农业科学院植物保护研究所,福建省作物有害生物监测与治理重点实验室,福建省作物有害生物绿色防控工程研究中心,福州 350013;2. 福建农林大学植物保护学院,福州 350002)
草地贪夜蛾Spodopterafrugiperda起源于美洲,对农业生产危害极大,被联合国粮农组织列为全球重大迁飞性农业害虫之一(Nagoshietal., 2017;Guietal., 2020),可危害玉米、高粱、大豆等200多种重要农作物(Harrisonetal., 2019;彭国雄等,2019)。草地贪夜蛾于2019年初入侵我国云南地区,现已扩散至全国大部分地区,给我国农业及粮食安全生产造成严重威胁(潘兴鲁等,2020;占军平等,2020)。福建地处亚热带,气候适宜常年种植玉米,为草地贪夜蛾越冬提供了良好的寄主条件,是草地贪夜蛾周年繁殖区,防控任务艰巨(邱良妙等,2020)。
针对草地贪夜蛾在我国的迁飞路径和危害特点,国内植保工作者已提出多种监测防治手段并提出了杀虫剂田间应用策略(刘妤玲等,2019),但过度依赖农药可能导致草地贪夜蛾产生抗药性问题。在草地贪夜蛾的起源地美国、巴西等国家,由于长期依靠化学防治,导致其对传统杀虫剂产生了高水平抗性(Al-Sararetal., 2006;Zhangetal., 2020)。因此,生物防治相关方法也已被提出,如发掘和利用昆虫病原真菌(徐毓笛等,2020)、捕食性天敌(孙贝贝等,2020)、寄生性天敌(汤印等,2020)等生物资源进行草地贪夜蛾防治。其中,利用昆虫病原微生物实施“以菌治虫”具有低毒、低残留、无污染、不易产生抗药性等优点,是害虫生物防治的关键措施,也是害虫绿色防控体系发展的趋势(宋晓兵等,2016)。
绿僵菌作为一类重要而常见的虫生真菌,可寄生多种害虫,在农、林及卫生害虫的防治中应用广泛(Donald and Dymond, 2004;张亚波等,2020)。国外在利用绿僵菌防治草地贪夜蛾方面已有较多报道,墨西哥、巴西等国家很多地区的玉米种植区内均发现大量被莱氏绿僵菌感染的草地贪夜蛾幼虫(Ruiz-Nájeraetal., 2013;Ordóez-Garcíaetal., 2015);Akutse等(2019)测定了多种昆虫病原真菌对草地贪夜蛾幼虫的毒力,其中金龟子绿僵菌Metarhiziumanisopliae对2龄幼虫的致死率低于10%。鉴于草地贪夜蛾在我国的入侵时间较短,国内关于绿僵菌防治草地贪夜蛾的研究刚刚起步。彭国雄等(2019)在室内条件下测定了金龟子绿僵菌CQMa421对草地贪夜蛾不同龄期幼虫、蛹、成虫和卵的杀虫活性,发现该菌株对草地贪夜蛾低龄幼虫具有杀虫活性并引起蛹黑化。2019年7~8月,郑亚强等(2019)在云南省曲靖市玉米田调查发现,田间草地贪夜蛾的莱氏绿僵菌感染率在2.52%~29.83%之间;雷妍圆等(2020)从广州市玉米田染菌草地贪夜蛾幼虫上获得一株莱氏绿僵菌,高浓度孢子悬浮液处理下草地贪夜蛾2龄幼虫的死亡率达100%。本研究从福建省不同地区收集罹病草地贪夜蛾幼虫僵虫以及其他鳞翅目和半翅目幼虫僵虫,对病原真菌进行常规分离及产孢培养,采用浸渍法测试其对草地贪夜蛾2龄幼虫和蛹的致病力,并对高致病力菌株进行鉴定,以期为草地贪夜蛾的生物防治提供优良菌株。
1 材料与方法
1.1 材料
供试菌株:采自福建省三明永安、漳州长泰、龙岩上杭、南平建瓯、莆田仙游、福州闽侯和永泰地区等田间罹病的草地贪夜蛾幼虫僵虫以及其他罹病鳞翅目和半翅目幼虫僵虫,在超净台内挑取带菌虫体少许孢子,采用划线法接种于萨氏麦芽糖琼脂酵母(SMAY)培养基(蛋白胨10 g、麦芽糖40 g、酵母浸粉2 g、琼脂20 g、蒸馏水1 L),置于光照培养箱(温度26±1℃, 相对湿度70%±5%,光周期L∶D=14 h∶10 h,)中培养。6~8 d后挑取孢子转接到新的SMAY培养基中进行单孢培养,10~15 d后刮取分生孢子置于高压灭菌处理的30%甘油中,震荡混匀后于-80℃冰箱贮存备用。供试菌株详细信息见表1。
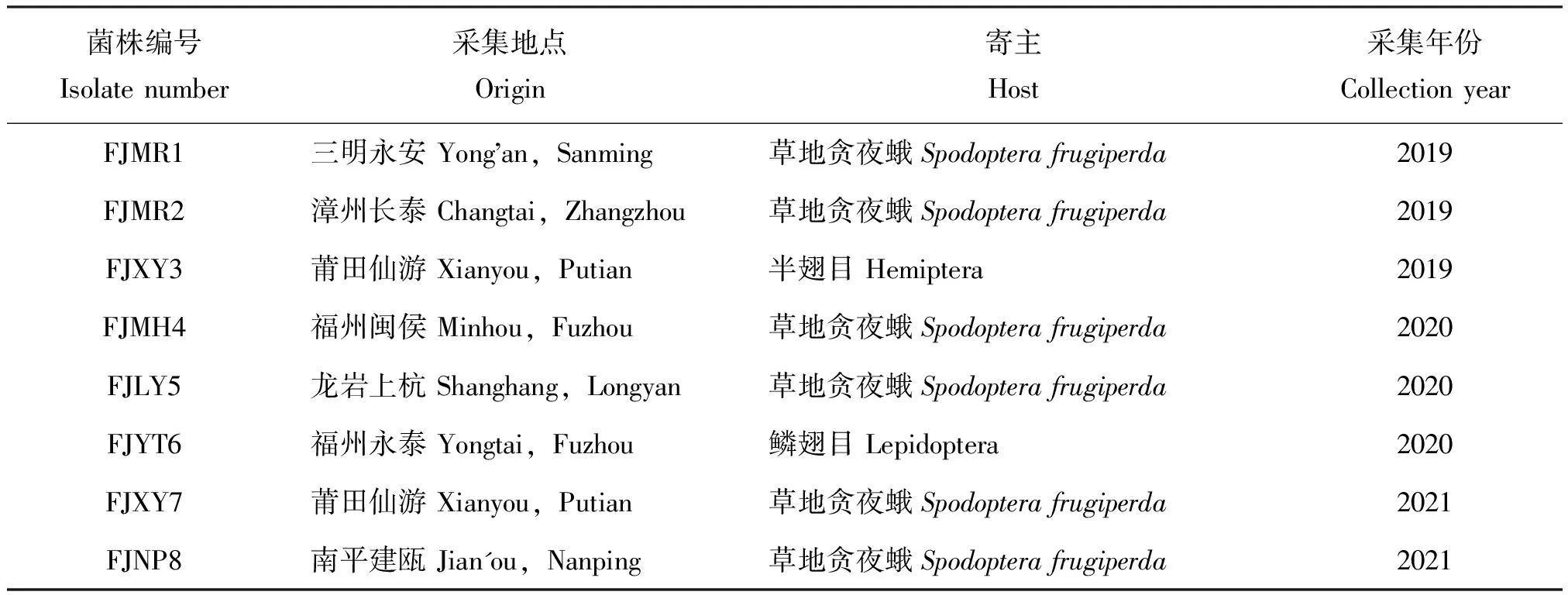
表1 供试绿僵菌菌株基本信息
供试虫源:供试草地贪夜蛾于2019年6月采自福建省建瓯市东峰镇玉米地,经室内人工饲料连续饲养的实验室种群,室内饲养条件为温度 26±1℃、相对湿度60%~70%、光周期L∶D=14 h∶10 h。选取2龄幼虫和1日龄蛹进行致病力测定。
人工饲料:草地贪夜蛾人工饲料参考李子园等(2019)方法配制。
1.2 孢子悬浮液的制备
将供试菌株接种于SMAY固体培养基,置于光照培养箱(温度26±1℃,相对湿度70%±5%,光周期L∶D=14 h∶10 h)中培养15 d,收集孢子,加入无菌0.05% Tween-80水溶液,充分震荡后用6层无菌纱布滤掉菌丝体获得孢子悬浮液,采用血球计数板计数,最终将孢子浓度调整为5×107个/mL备用。
1.3 菌株对草地贪夜蛾2龄幼虫的致死率测定
挑取大小一致、健康的草地贪夜蛾2龄幼虫在浓度为5×107个/mL的供试菌株孢子悬浮液中浸渍5 s后取出晾干,然后将其移置人工气候箱内(温度26±1℃,相对湿度65%±5%)用塑料养虫盒(直径为3 cm,高为3 cm)单头饲养,养虫盒内放入人工饲料供其取食。设3次重复,每重复20头幼虫,以0.05% Tween-80无菌水处理的2龄幼虫为对照。每日定期记录试虫的死亡数量,持续观察8 d。死亡试虫及时挑出保湿培养,若体表长出白色菌丝并产绿色孢子,则视为染菌死亡。
1.4 菌株对草地贪夜蛾蛹的致死率测定
选取大小一致的1日龄蛹,放入配制好的浓度为5×107个/mL的供试菌株孢子悬浮液浸渍5 s后取出晾干,然后将其移置人工气候箱内(温度26±1℃,相对湿度65%±5%)用塑料养虫盒(直径为6 cm,高为5 cm)饲养,养虫盒内放入棉球保湿,每个养虫盒中放入5头蛹。设3次重复,每重复20头蛹,以0.05% Tween-80无菌水处理的草地贪夜蛾蛹作为对照。每日定期记录蛹的死亡情况(蛹体多次触碰均无反应则视为死亡),持续观察8 d。死亡蛹体及时挑出保湿培养,若体表长出白色菌丝并产绿色孢子,则视为染菌死亡。
1.5 菌株鉴定
菌株形态学观察:将高致力菌株FJMR2和FJXY7接种于SMAY培养基,于温度26±1℃、相对湿度70%±5%、光周期L∶D=14 h∶10 h的人工气候箱中培养7 d,记录菌落培养特征,并在光学显微镜下观察菌丝和分生孢子形态以及产孢结构。
分子鉴定:提取纯化菌株FJMR2和FJXY7的基因组DNA,利用真菌通用引物IT1(5′-TCCGT AGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGC TTATTGATATGC-3′)对菌株rDNA-ITS序列进行普通PCR扩增。50 μL PCR反应体系:DNA模板2 μL(20 ng/μL),上、下游引物各2 μL(10 μmol/L),2×EsTaq PCR Master Mix 25 μL,ddH2O补足至50 μL。反应条件:94℃预变性5 min;94℃ 40 s,56℃ 30 s,72℃ 30 s,35个循环;72℃延伸 5 min。PCR产物经1.0%琼脂糖凝胶电泳检测后送生工生物工程(上海)股份有限公司测序。测序结果在GenBank基因数据库中比对后选择相近序列,采用MEGA 7.0软件的邻接法重复1 000次构建系统发育树。
1.6 统计分析
试验数据通过Excel 2013和SPSS 17.0软件进行统计分析,采用Duncan氏新复极差法检验,分析比较不同菌株间的致病力差异,并利用Probit方法计算半数致死时间(LT50)。
死亡率(%)=死虫数/试虫总数×100。
校正死亡率(%)=(处理组死亡率-对照组死亡率)/(100-对照组死亡率)×100。
2 结果与分析
2.1 供试绿僵菌菌株对草地贪夜蛾2龄幼虫的致病力
用5×107个/mL孢子浓度的悬浮液处理8 d后,8株绿僵菌菌株对草地贪夜蛾2龄幼虫的致病力差异显著,其中菌株FJMR2和FJXY7对草地贪夜蛾2龄幼虫的致死率最高,分别为88.76%和82.13%;其次为菌株FJNP8和FJMR1,致死率分别为77.78%和76.19%;菌株FJMH4、FJYT6和FJLY5的致死率在52.33%~60.70%之间;菌株FJXY3的致死率最低,仅为28.25%。菌株FJMR2和FJXY7对草地贪夜蛾2龄幼虫的致死速率最快,半数致死时间(LT50)分别为4.81 d和4.93 d;其次为菌株FJNP8和FJMR1,LT50分别为5.37 d和5.53 d;菌株FJYT6、FJLY5和FJMH4的LT50在6.94 d~7.14 d之间;菌株FJXY3的致死率低于50%,故无法计算LT50(表2)。
2.2 供试绿僵菌菌株对草地贪夜蛾蛹的致病力
用5×107个/mL孢子浓度的悬浮液处理8 d后,8株绿僵菌菌株对草地贪夜蛾蛹的致病力差异显著,其中菌株FJMR2和FJXY7对蛹的致死率最高,分别为86.57%和84.00%;其次为菌株FJNP8、FJMR1和FJMH4,致死率在72.74%~75.39%之间;菌株FJLY5的致死率为56.55%;菌株FJXY3和FJYT6的致死率均低于50%。菌株FJMR1、FJMR2、FJMH4和FJXY7对蛹的半数致死时间(LT50)为4.83~5.86 d,差异不显著;菌株FJLY5和FJNP8对蛹的LT50分别为6.63 d 和6.29 d;菌株FJXY3和FJYT6对蛹的致死率均低于50%,故无法计算LT50(表3)。
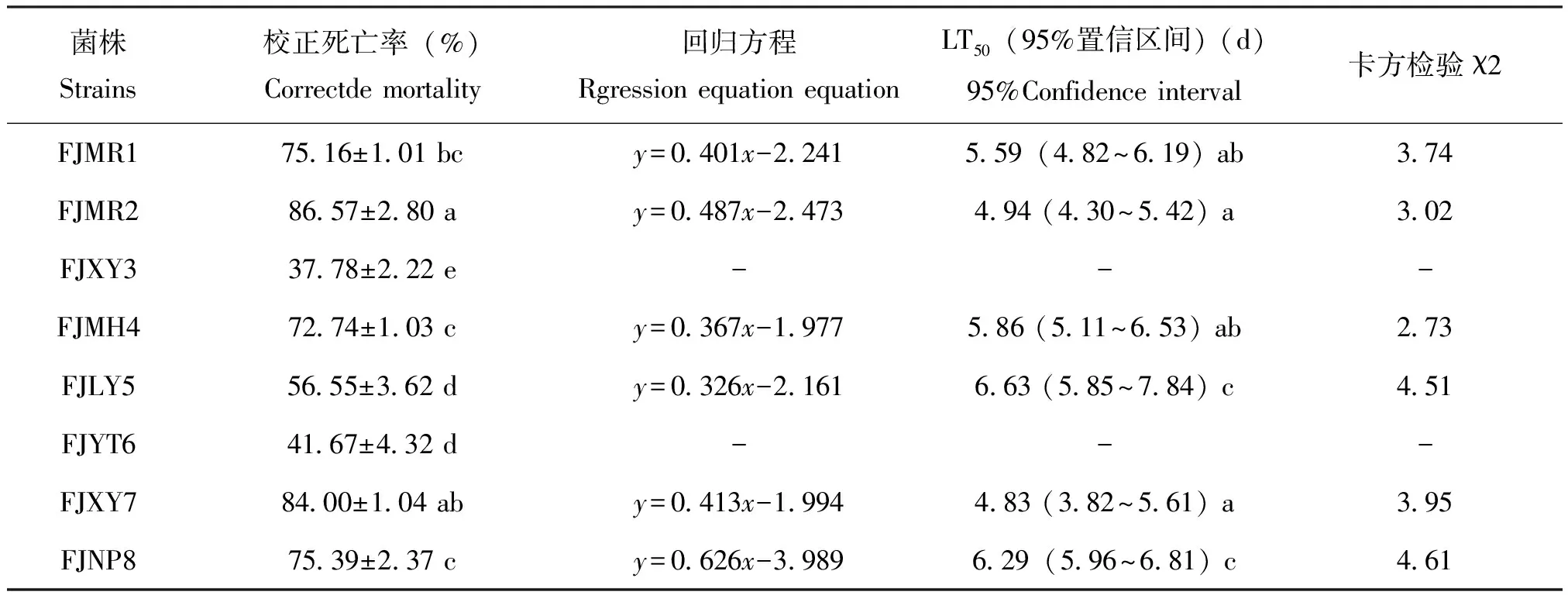
表3 8株绿僵菌菌株对草地贪夜蛾蛹的毒力测定
总体上,菌株FJMR2和FJXY7对草地贪夜蛾2龄幼虫和蛹的致死率最高、半数致死时间最短。
2.3 菌株FJMR2和FJXY7的种类鉴定
菌株FJMR2和FJXY7在SMAY培养基上的菌落形态相似。培养7 d后,菌落直径6~9 mm,质地短绒状凸起,正面乳白色,反面土黄色,无分泌物;菌丝表面光滑透明,具分隔,直径1.1~4.2 μm;分生孢子梗着生于气生菌丝,圆柱形,近基部明显膨大,大小2.5~5.5×1.5~3.0 μm,瓶梗呈轮生状,每轮3~5个;分生孢子链生,卵圆形或椭圆形,大小2.5~4.0×1.5~2.5 μm,表面光滑(图1-C)。初步鉴定菌株FJMR2和FJXY7为莱氏绿僵菌Metarhiziumrileyi。

图1 莱氏绿僵菌形态特征及草地贪夜蛾2龄幼虫和蛹感染莱氏绿僵菌的症状(以FJMR2为例)Fig.1 Morphological characteristics of Metarhizium rileyi and symptoms of infected Spodoptera frugiperdaexternal symptoms of larvae, pupa of Spodoptera frugiperda after infection Metarhizium riley (Take FJMR2 as an example)注:A,菌株生长初期;B,菌株生长后期;C,分生孢子和菌丝形态;D,田间被绿僵菌感染的僵虫;E,2龄幼虫染病初期;F,2龄幼虫染病后期;G,蛹染病后期;H,蛹染病前期。Note: A, Initial stage of colony growth; B, Late stage of colony growth; C, Mycelia and branched conidiophores; D, Symptoms of S.frugiperda infected by M.rileyi in field; E, Initial stage of the 2nd instar larvae infected by M.rileyi; F, Late stage of the 2nd instar larvae infected by M.rileyi; G, Late stage of the pupae infected by M.rileyi; H, Initial stage of the pupae infected by M.rileyi.
以ITS1和ITS4为引物,从菌株FJMR2和FJXY7基因组DNA分别扩增出1条约550 bp的序列。将获得序列在NCBI数据库中进行BLAST相似性比对,发现该2株菌株与MetarhiziumrileyiCBS 806.71(GenBank登录号MH872111)的同源性高达99.9%以上,位于系统发育树最小进化分支,亲缘关系最近(图2)。综合菌株培养特征、形态特征和分子生物学方法鉴定,最终确定菌株FJMR2和FJXY7均为莱氏绿僵菌Metarhiziumrileyi。
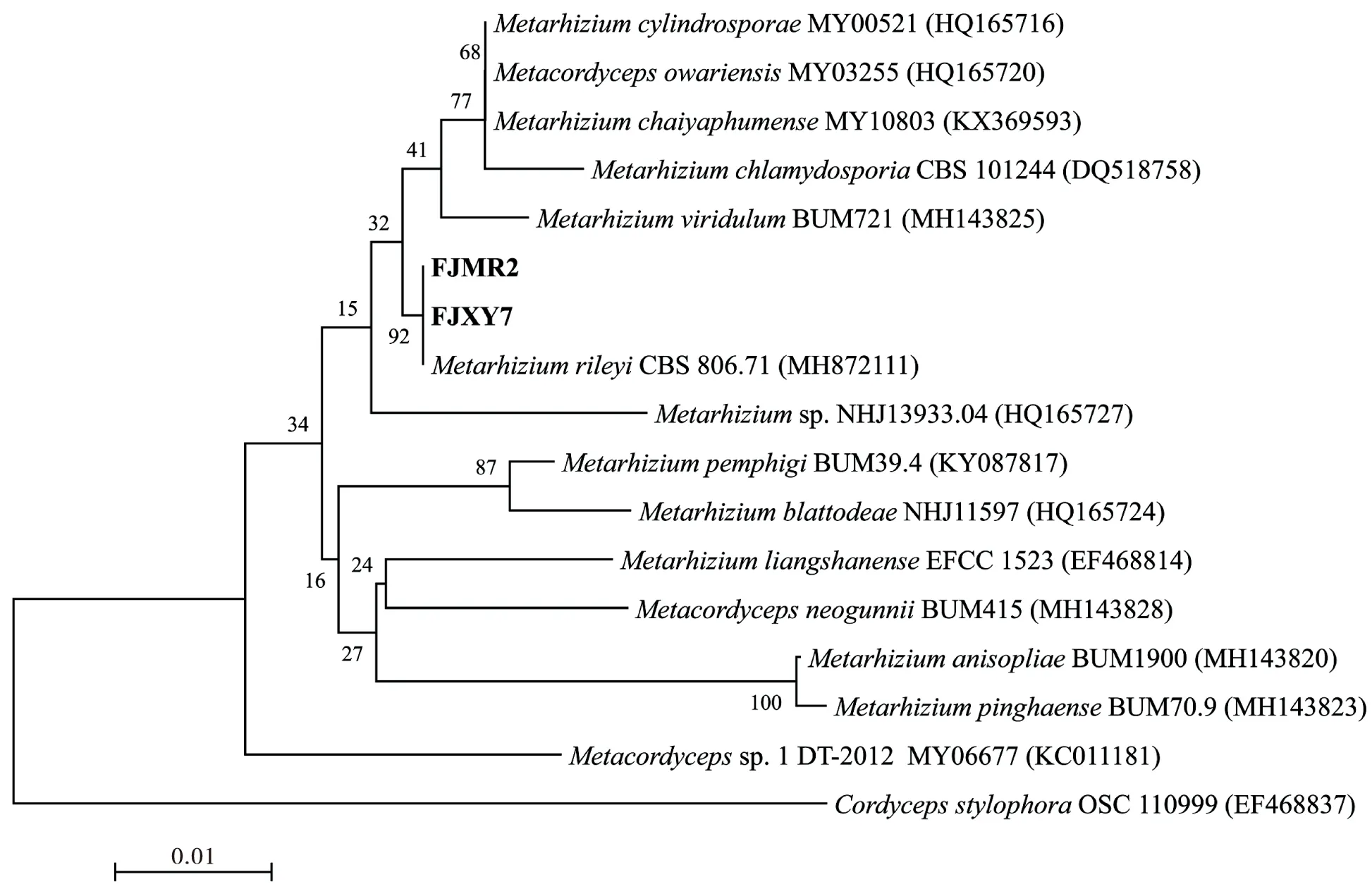
图2 基于rDNA-ITS序列构建的菌株FJMR2和FJXY7与相关种的系统发育进化树Fig.2 Phylogenetic tree of the strain FJMR2 and FJXY7 with others related species based on rDNA-ITS sequences
3 结论与讨论
绿僵菌作为重要的害虫生防真菌,国内外在其物种资源、致病力研究及生防产品开发中已取得重要成果(蒲蛰龙和李增智,1996)。鉴于草地贪夜蛾在我国的入侵时间较短,有关草地贪夜蛾绿僵菌资源的研究主要集中在金龟子绿僵菌和莱氏绿僵菌(彭国雄等,2019;雷妍圆等,2020)。致病力是评价菌株应用潜力的核心指标,也是筛选优良菌株的基本依据,为进一步挖掘防治草地贪夜蛾的优良菌株,本研究测定了分离自福建省不同地区不同寄主的8株绿僵菌菌株对草地贪夜蛾2龄幼虫和蛹的致病力,发现8株供试菌株对草地贪夜蛾均有不同程度的致病力,致死率为28.25%~88.76%,LT50为4.81~7.14d。从中筛选出2株对草地贪夜蛾具较强致病力的菌株FJMR2和FJXY7,根据菌株形态特征和rDNA-ITS序列对比分析,鉴定2株菌株均为莱氏绿僵菌Metarhiziumrileyi。
莱氏绿僵菌又名莱氏野村菌Nomuraearileyi,可侵染多种鳞翅目害虫,尤其针对夜蛾科多种为害较大的害虫,如斜纹夜蛾SpodopteralituraFabricius、银纹夜蛾Argyrogrammaagnata(Staudinger)、甜菜夜蛾SpodopteraexiguaHübner等,是一种生防潜力极高的虫生真菌,在适宜的自然环境下能够引发害虫的病害流行,显著降低害虫的种群密度(Srisukchayakuletal.,2005;罗卿权等,2018)。周立峰等(2012)测定了5株莱氏绿僵菌对斜纹夜蛾的毒力,第10天的侵染率在60%~95%之间。胥志民等(2019)研究发现,莱氏绿僵菌Nr 5772在5×109个/mL孢子浓度下对甜菜夜蛾3、4、5龄幼虫的LT50分别为4.44 d、4.48 d和4.85 d。雷妍圆等(2020)用孢子浓度为1×109个/mL的莱氏绿僵菌GZSF-1处理草地贪夜蛾2龄幼虫,7 d后幼虫的死亡率达100%。本研究结果显示,莱氏绿僵菌FJMR2和FJXY7对草地贪夜蛾2龄幼虫具有较高致病力,在5×107个/mL孢子浓度下对2龄幼虫的致死率均达80%以上,与龙秀珍等(2021)的研究报道结果一致。另研究发现,FJMR2和FJXY7能侵染草地贪夜蛾蛹,在5×107个/mL浓度下对蛹的致死率达84%以上,大大降低了蛹的羽化率;而龙秀珍等(2021)研究发现,莱氏绿僵菌CDTLJ1对草地贪夜蛾蛹的致死率仅有20%,可能是菌株地理来源不同,致病性差异较大造成的。土壤具有更好的保湿保温能力,又可避免紫外线的照射,为孢子的萌发创造了有利条件,更适合绿僵菌等杀虫真菌对土壤中栖息害虫的防治(Maetal.,2019),而且在土壤中施加菌剂以防治地下害虫被认为是一种安全有效且可持续的害虫防控措施(Roggeetal.,2017)。鉴于草地贪夜蛾大部分在土壤中化蛹,可以考虑在土壤中添加菌剂,达到对草地贪夜蛾持续控制的目的。
综上所述,文中筛选到2株对草地贪夜蛾2龄幼虫及蛹有较高致病力的莱氏绿僵菌FJMR2和FJXY7,具有研究价值和应用潜力,但本文研究结果是基于稳定的室内环境条件下取得的,如何让该菌株在复杂的自然环境中发挥对草地贪夜蛾的持续控制作用仍有待进一步研究。
