不同水位条件下鄱阳湖越冬白鹤的移动模式与栖息地利用*
2023-10-19江红星陈丽霞王艺璇
王 莹 江红星 陈丽霞 王艺璇 高 彤
(中国林业科学研究院森林生态环境与自然保护研究所 生物多样性保护国家林业和草原局重点实验室 北京 100091)
白鹤(Leucogeranus leucogeranus)是世界上15 种鹤类中特定依赖湿地生存的3 种鹤类之一(Meineet al.,1996),是我国I 级重点保护野生动物,世界自然保护联盟红皮书将其列为“极危”等级。白鹤全球种群分为西部、中部和东部3 个种群,它们均在俄罗斯北部繁殖,迁徙数千公里,分别在伊朗、印度和中国越冬。目前,白鹤全球种群数量估计为3 500~4 000 只,几乎全部为东部种群(http://www.birdlife.org)。白鹤东部种群繁殖于俄罗斯西伯利亚东北部的苔原地带,迁徙途径我国松嫩平原、辽河平原和华北平原,98%的种群在江西鄱阳湖越冬(Kanaiet al.,2002;马克·巴特等,2005;Houet al.,2020)。因此,鄱阳湖湿地生态环境的稳定性对于全球白鹤种群的稳定与发展至关重要。
近些年来,鄱阳湖水位的年内和年际间变化,导致白鹤在鄱阳湖的空间分布格局和栖息地利用模式发生了显著变化(Houet al.,2020)。历史上,鄱阳湖越冬白鹤主要以湖区浅水沼泽和泥滩中的沉水植物苦草(Vallisneriaspp.)块茎为主要食物(曾南京等,2002;孙志勇等,2010;吴建东等,2013;Burnhamdet al.,2017)。鄱阳湖4—10 月为沉水植物苦草的生长期,其中7—10 月为冬芽形成期(万文豪等,2002),水位过深或过浅都会抑制苦草光合作用,影响苦草生长和冬芽形成(Dobberfuhl,2007;Wanget al.,2008),进而影响越冬期白鹤觅食栖息地的选择(Wuet al.,2009;黎磊等,2015)。
2016 年越冬期以来,在鄱阳湖水位变化加剧,越冬白鹤对周边农田栖息地的利用率显著增加,特别是藕塘和稻田(王文娟等,2019;Houet al.,2020),但有关越冬白鹤的移动模式和栖息地利用尚缺乏定量对比研究。本文基于2016—2022 年13 只卫星跟踪白鹤在鄱阳湖越冬期的定位数据,利用 ArcGIS 10.7 空间分析功能和鄱阳湖区土地利用类型数据,对比分析丰水年和平水年越冬白鹤的移动距离、活动区面积大小、栖息地利用情况,以便为鄱阳湖越冬白鹤种群保护和栖息地适应性管理提供科学依据。
1 研究区概况
鄱阳湖(115°49′—116°46′E,28°11′—29°51′N)位于江西省北部和长江以南,是中国最大的淡水湖。鄱阳湖是与长江连通的典型季节性吞吐湖泊,受亚热带季风性气候、长江和鄱阳湖五河注入的影响,水位年变化幅度大。每年4—9 月为丰水期,10 月—翌年3 月为枯水期,丰水和枯水季节湖面水位变化最大可达9~14 m,有“高水是湖,低水似河”的独特景观(张本,1988)。在枯水季节随着鄱阳湖水位下降,鄱阳湖周边不同高程的浅水洼地相继与主湖分开,形成子湖泊。这些子湖泊地势平缓,多呈浅碟型,其生境呈“草滩—泥滩—浅水水域”的环带状分布,这种复杂多样的湿地生境为不同生态环境需求的越冬水鸟提供了重要的食物资源和栖息场所(马克·巴特等,2005;金斌松等,2016)。
2 研究方法
2.1 白鹤卫星跟踪
2016—2020 年,在我国东北白鹤中途停歇地对救助恢复后的13 只白鹤佩戴了背负式卫星跟踪器。跟踪器由湖南环球信士科技有限公司生产,型号为HQBG3621S 和HQBG5037S,质量分别为24 g 和40 g,符合跟踪器质量不超过研究对象体质量3%~5%的国际标准(Higuchiet al.,1996;1998)。本研究仅对定位精度包括A、B、C 3 个等级的数据进行分析,定位误差分别为5、10、20 m。
2.2 数据分割
以鄱阳湖的北端29.51°N 为标志,确定13 只白鹤跟踪个体每年迁到与迁离鄱阳湖的日期,并统计不同年份的越冬天数与有效位点数(表1)。

表1 2016—2021 年越冬期鄱阳湖白鹤卫星跟踪信息Tab. 1 Satellite tracking information of Siberian Cranes in winters from 2016 to 2021 in Poyang Lake
不同水位年组界定,主要依据鄱阳湖水文站水位数据以及湖区白鹤自然食物苦草块茎生物量。自2003 年以来,三峡大坝的运行对鄱阳湖水位变化产生了重要影响(图1a)。对比2003—2022 年间鄱阳湖星子站4—10 月平均水位与2016—2021 年度4—10 月平均水位,2016、2017、2020 和2021 年度4—10 月平均水位均高于2003—2022 年间平均水位,而2018 和2019 年度4—10 月平均水位均低于2003—2022 年间平均水位(图1b)。结合郝能祖等(2019)和余定坤等(2020)监测结果,2018 年和2019 年湖区的苦草密度均高于其他4 个年份。鉴于此,本研究将6 个研究年份划分为3 个水位年组:2016 年和2017 年越冬期为W1 丰水年组、2018 年和2019 年越冬期为W2 平水年组、2020 年和2021 年越冬期为W3 丰水年组。丰水年组内和组间的4—10 月水位差异不显著,平水年组内的4—10 月水位差异不显著。
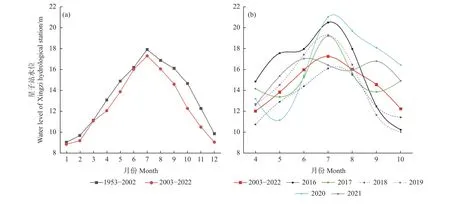
图1 星子水文站月平均水位Fig. 1 The monthly average water levels of Xingzi Hydrological Station
根据历年白鹤的越冬习性,将白鹤迁到鄱阳湖至12 月中旬定义为越冬前期;12 月下旬至翌年2 月中旬定义为越冬中期,白鹤种群数量与分布相对稳定;翌年2 月下旬至迁离鄱阳湖定义为越冬后期(夏少霞等,2010;胡振鹏等,2012)。根据杨秀林(2019)通过计算白鹤每天每小时移动距离,将6:00—17:59 期间的定位数据界定为白鹤的日间活动位点,将18:00—翌日5:59 期间的定位数据界定为白鹤的夜间活动位点。
2.3 移动距离与活动区大小
不同越冬阶段的白鹤移动距离为相应越冬阶段连续2 个位点间欧式距离之和。越冬期白鹤移动总距离为整个越冬期连续2 个位点间的欧式距离之和。
活动区(home range)面积在一定程度能反映栖息地质量,目前多使用核密度估计法(Kernel Density Estimator,KDE)(Worton,1995;Powell,2000)。为剔除白鹤处于飞行状态的位点,本研究筛选出速度小于3 km·h-1的位点用于活动区面积计算。使用R 4.2.0“adehabitatHR”包生成每只白鹤个体不同越冬阶段的90%KDE 和50%KDE 范围,其中90%KDE 代表活动区大小,50%KDE 代表核域大小,将生成的shp 文件导入ArcGIS 10.7 分别计算90%KDE 和50%KDE 面积。
2.4 土地利用类型分类
从中国科学院资源环境科学数据中心(http://www.resdc.cn/)下载研究区2015、2018 和2020 年的土地利用类型数据。该数据包括6 个一级地类和25 个二级地类。考虑到白鹤主要利用稻田、藕塘、自然湿地和草地生境(Jiaet al.,2013;Burnhamdet al.,2017;王文娟等,2019;Houet al.,2020),使用ArcGIS 10.7 的重分类功能,提取研究区的农田(水田、旱地)、湿地(湖泊、沼泽地、滩地)、草地和其他类型(林地、城乡、工矿、居民用地和未利用土地)等4 种土地利用类型。利用Spatial Analyst 工具中的多值提取至点功能,提取跟踪白鹤个体日间和夜间卫星跟踪位点所对应的土地类型。不同越冬阶段各土地类型的利用率为对应阶段该土地类型位点数除以总位点数(Mintonet al.,2003;王昱熙等,2020)。
2.5 统计分析
所获得的数据从两方面进行统计分析:1)3 个水位年组(W1、W2、W3)之间的对比分析;2)丰水年组(W1 和W3 合并)与平水年组(W2)的对比分析。
首先进行独立样本Kolmogorov-Smirnov Test 检验数据是否符合正态性。若符合正态分布,采用单因素方差分析(One-way ANOVA)分析和2 个独立样本t检验。若不符合正态分布,采用非参数检验方法中的多个独立样本Kruskal-Wallis H 检验和2 个独立样本Mann-Whitney U 检验。统计数据均表示为算术平均值±标准差(Mean ± SD),统计分析在SPSS 22.0 中进行。
3 结果与分析
3.1 越冬期不同阶段的移动距离
越冬期白鹤移动总距离在丰水年W1 组、W3 组以及合并组(W1+W3)分别为[(1 368 ± 446) km,n=7]、[(1 544 ± 240) km,n=6]和[(1 398 ± 398) km,n=13],平水年W2 组为[(760 ± 420) km,n=9]。其中,丰水年3 组的白鹤移动总距离显著大于平水年W2 组(W1/W2 组:P= 0.006;W3/W2 组:P= 0.001;合并组/W2 组:P=0.001);丰水年组之间的白鹤移动总距离无显著差异(P> 0.05)。
从越冬期不同阶段的白鹤移动距离来看,越冬前期丰水年合并组的白鹤移动距离显著大于平水年W2组(合并组/W2 组:P= 0.034),其他组间无显著差异;越冬中期,丰水年3 组的白鹤移动距离显著大于平水年W2 组(W1/W2 组:P= 0.000;W3/W2 组:P= 0.001;合并组/W2 组:P= 0.000);越冬后期,各组间无显著差异(P> 0.05)(图2)。
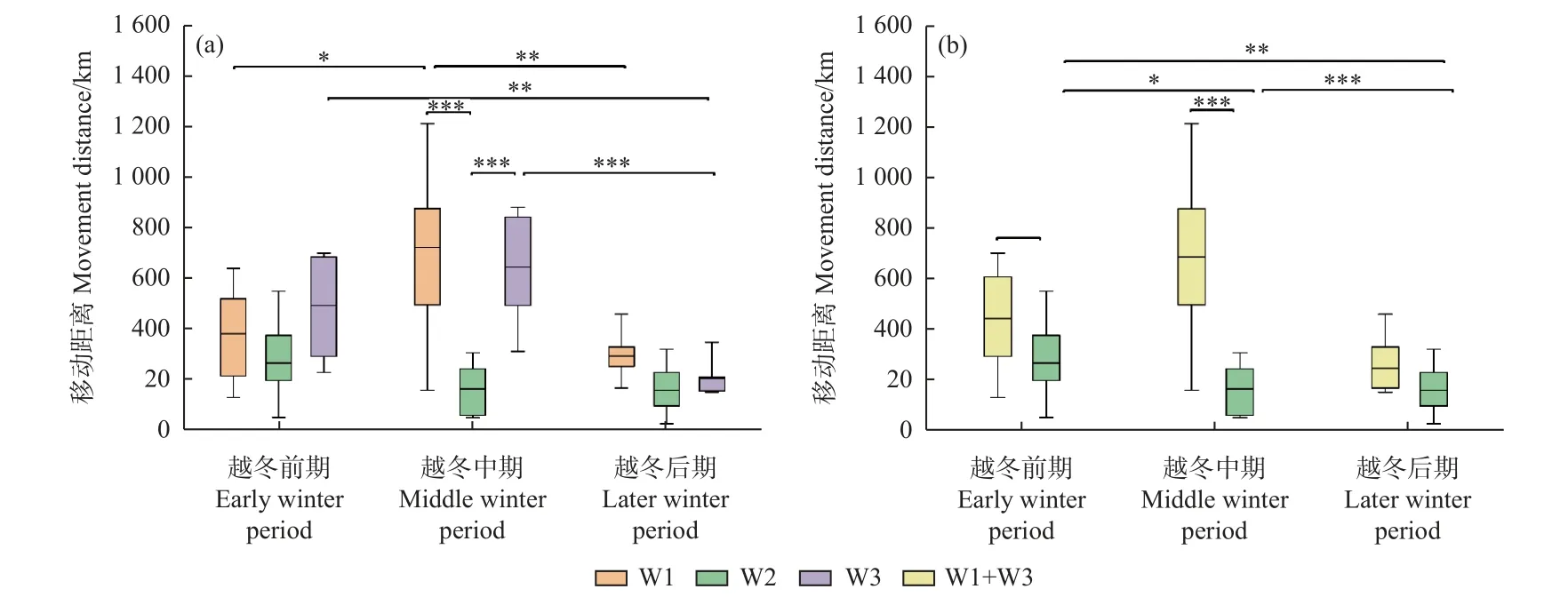
图2 越冬期不同阶段的白鹤移动距离Fig. 2 Movement distances of Siberian Cranes in different winter stages
从丰水年和平水年越冬期不同阶段的移动距离可知,丰水年3 组白鹤移动距离为越冬中期 > 前期 >后期,除W1 组越冬前期和后期间的移动距离及W3组越冬前期和中期间的移动距离差异不显著外,其余均差异显著(P< 0.05)。平水年W2 组不同越冬阶段的白鹤移动距离均差异不显著(P> 0.05)(图2)。
3.2 越冬期不同阶段的活动区面积
从越冬期白鹤活动区大小来看,丰水年3 组白鹤90%KDE 活动区面积分别为[(3 771 ± 3 043) hm2,n=7]、[(2 192 ± 352) hm2,n=6]和[(2 935 ± 2 052) hm2,n=13],50%KDE 活动区面积分别为(467 ± 377) hm2、(307 ± 117) hm2和(402 ± 298) hm2。平水年W2 组白鹤的90%KDE 和50%KDE 活动区面积分别为[(1 898 ±1 056) hm2,n=9] 和[(328 ± 220) hm2,n=9],各水位年组间均无显著差异(P> 0.05)。
从越冬期不同阶段的白鹤活动区大小来看,越冬前期,丰水年合并组90%KDE 活动区面积显著大于平水年W2 组(P= 0.027),其余各组间均无显著差异;越冬中期,丰水年W1 组的90%和50%KDE 活动面积均显著大于平水年W2 组和丰水年W3 组(W1/W2 组:P=0.000;P=0.002;W1 组/W3 组:P=0.003;P=0.007),丰水年合并组90%和50%KDE 活动区面积均显著大于平水年W2 组(P=0.036;P=0.007);越冬后期,丰水年合并组90%KDE 活动区面积显著大于平水年W2 组(P=0.025)(图3)。
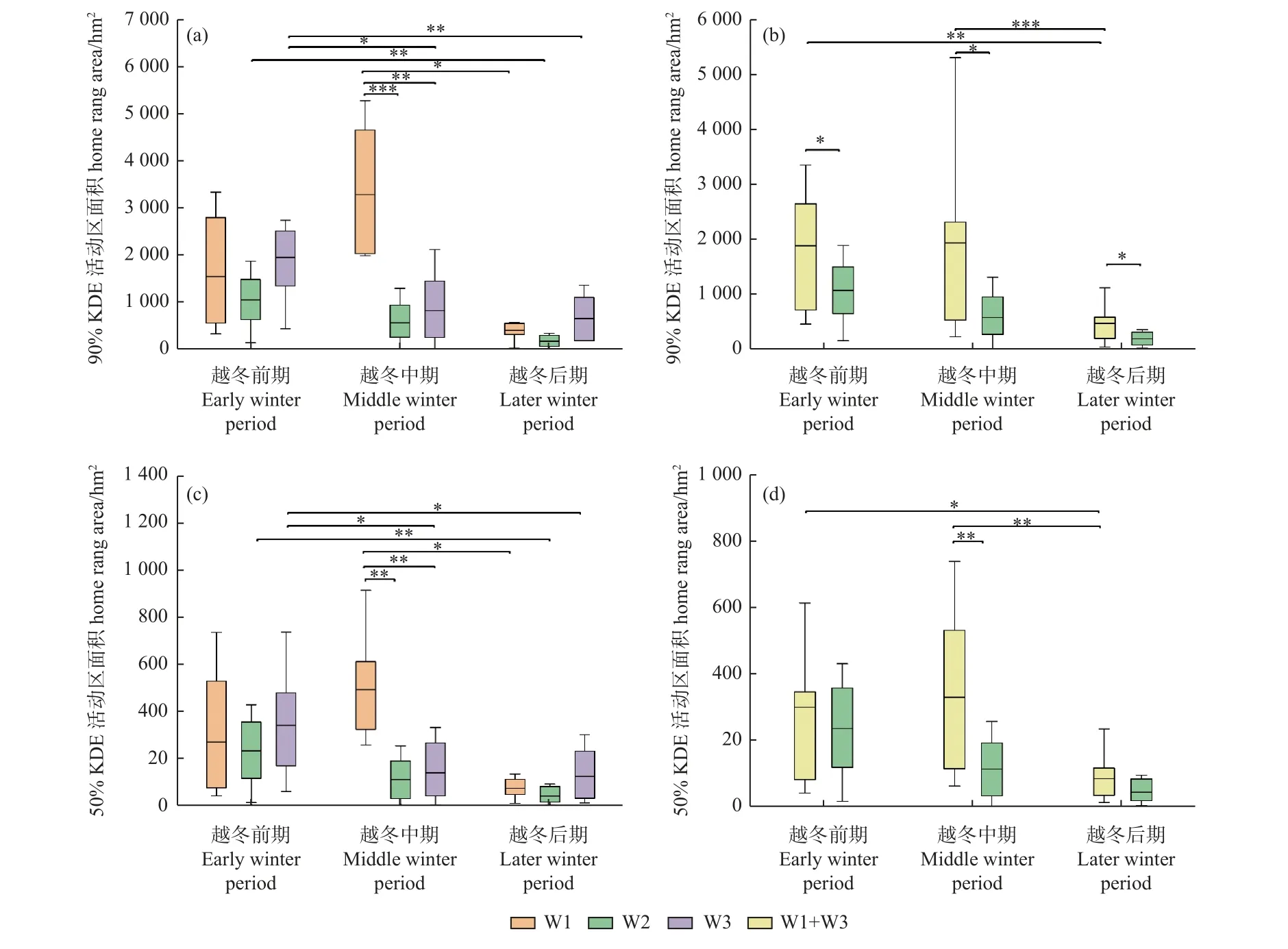
图3 越冬期不同阶段的白鹤活动区面积Fig. 3 Home ranges of Siberian Cranes in different winter stages
从丰水年和平水年越冬期不同阶段的活动区面积可知,丰水年W3 组和合并组白鹤的90%和50%KDE活动区面积为越冬前期 > 中期 > 后期,除W3 组越冬中期和后期,以及合并组越冬前期与中期活动区面积差异不显著,其他阶段均差异显著;W1 组为越冬中期 >前期 > 后期,仅越冬中期和后期差异显著(P= 0.014;P= 0.025)。平水年W2 组90%KDE 和50%KDE 为越冬前期 > 中期 > 后期,其中仅越冬前期和后期差异显著(P= 0.004;P= 0.009)(图3)。
3.3 越冬期不同阶段的栖息地利用率
3.3.1 越冬期不同阶段的日间栖息地利用率 从白鹤越冬期日间栖息地利用率来看,丰水年3 组白鹤对农田利用率分别为52.2% ±22.7%(n=7)、44.7% ± 20.1%(n=6)和48.2% ± 20.9%(n=13),对湿地利用率分别为44.9% ± 21.1%、55.2% ± 20.0%和50.4% ± 20.5%。平水年组W2 白鹤对农田利用率为11.0% ± 12.3%(n=9),对湿地利用率为(89.0% ± 12.3%)。丰水年3 组白鹤对农田利用率显著高于平水年组(W1/W2 组:P=0.006;W3/W2 组:P= 0.016;合并组/W2 组:P= 0.000),对湿地利用率显著低于平水年组(W1/W2 组:P=0.001;W3/W2 组:P= 0.019;合并组/W2 组:P= 0.000)。丰水年组间白鹤对农田和湿地利用率无显著差异(P> 0.05)。
从越冬期不同阶段的白鹤日间栖息地利用率来看,越冬前期和中期,丰水年3 组白鹤对农田利用率均显著高于平水年组(P< 0.05),对湿地利用率均显著低于平水年组(P< 0.05)。越冬后期,各组间农田利用和湿地利用率差异不显著(P> 0.05)(图4)。
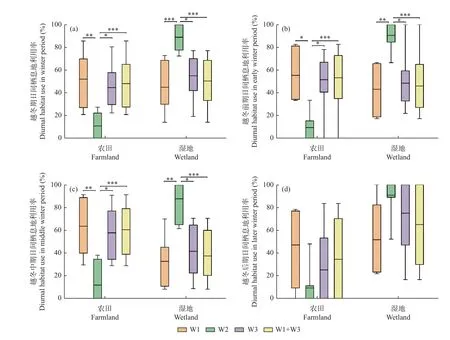
图4 不同水位年组白鹤的日间栖息地利用比例Fig. 4 Diurnal habitat use ratio of Siberian Cranes in different water year groups
从丰水年和平水年越冬期不同阶段的日间栖息地利用率来看,丰水年3 组和平水年组白鹤3 个越冬阶段的农田和湿地利用率均无显著差异(P> 0.05)(图4)。
3.3.2 越冬期不同阶段的夜间栖息地利用率 从白鹤越冬期夜间栖息地利用率来看,丰水年3 组白鹤农田利用率分别为15.7 ± 22.2%(n=7)、15.1% ± 16.1%(n=6)和15.4% ± 18.4%(n=13),湿地利用率分别为84.1% ± 22.1%、84.9% ± 16.0%和84.5% ± 18.4%,平水年组W2 的白鹤农田利用率为5.1% ± 8.7%(n=9),湿地利用率为94.9% ± 8.7%。丰水年组与平水年组的白鹤夜间农田和湿地利用率均无显著差异(P> 0.05)。
从越冬期不同阶段的白鹤夜间栖息地利用率来看,越冬中期,丰水年W1 组和合并组的白鹤农田利用率均显著高于平水年组(P< 0.05),湿地利用率均显著低于平水年组(P< 0.05)(图5)。
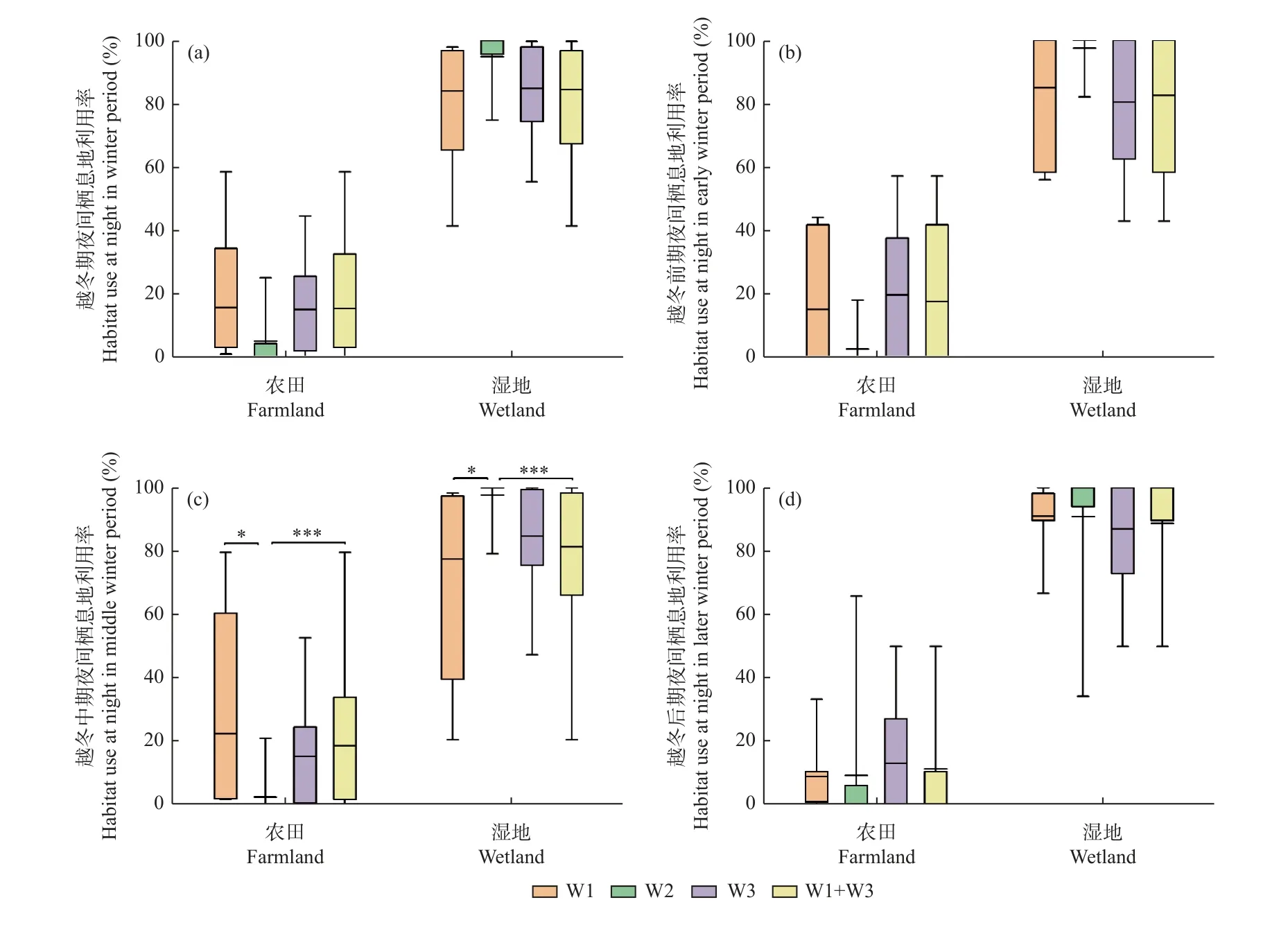
图5 白鹤在不同水位年组的夜间栖息地利用比例Fig. 5 Habitat use ratio of Siberian Cranes at night in different water year groups
从丰水年和平水年越冬期不同阶段的夜间栖息地利用率来看,丰水年3 组和平水年组的白鹤3 个越冬阶段的农田和湿地利用率均无显著差异(P> 0.05)(图5)。
4 讨论
4.1 不同水位年白鹤越冬期的移动模式
避难所理论(refuging theory)认为鸟类每天的移动距离是对栖息地周边食物资源可获得性的一种适应性行为,也是增加自身适合度的一种觅食策略(Hamiltonet al.,1970)。随着时间推移,由于栖息地附近的食物资源枯竭,聚集的鸟类个体将增加栖息地和觅食地之间的移动距离,来满足能量补给需求;相应地,个体付出代价和风险也随之增加。每天的移动距离会影响个体的能量消耗和能量平衡,水鸟更多倾向于选择较近区域觅食(Gauthieret al.,1984;MacArthuret al.,1996;Van der Graafet al.,2005)。当栖息地食物资源缺乏时,鸟类会增加移动距离即去较远区域觅食,其每日能量摄入与消耗之间的平衡可能会受到移动模式的影响(Pearseet al.,2010;Bavecoet al.,2011)。
研究发现平水年白鹤越冬期移动距离显著小于丰水年,平水年越冬3 个阶段的90%KDE 活动区面积也均显著小于丰水年合并组的相应越冬阶段,这表明平水年白鹤喜食的苦草等自然食物资源充足,白鹤很容易从栖息地及其周围得到能量补给,无需增加移动成本去较远区域觅食。这与吴海峰等(2018)对鄱阳湖越冬白枕鹤(Grus vipio)活动区面积随着水位升高而增大的研究结果一致。平水年白鹤移动距离随着越冬时间推移而逐渐减小,与越冬中期和后期鄱阳湖水位下降,洲滩裸露,自然食物苦草冬芽的可获得性增加有关(孙志勇等,2010)。
相反,丰水年苦草等沉水植物的生长受高水位影响,越冬期自然食物资源短缺,白鹤被迫增加移动距离去较远区域觅食,以满足自身能量摄入需求。丰水年移动距离随着越冬时间推移出现先增后减现象,与自然食物短缺和种间竞争有关,导致白鹤越冬前期和中期大量进入藕塘和稻田等农田栖息地觅食,越冬后期白鹤在自然湿地集群以待北迁(王文娟等,2019;Houet al.,2020)。刘成林等(2011)研究表明鄱阳湖水位越高,草滩和浅水区越冬候鸟栖息地越小,鸟类生存的自然栖息空间进一步缩小。枯水期鄱阳湖水位14.18 m(星子站,黄海高程)以上时, 水陆过渡带将缩小乃至消失,鄱阳湖湿地作为候鸟栖息地的功能将丧失。
4.2 不同水位年白鹤越冬期的觅食策略
食物资源的丰富度与可获得性是影响越冬水鸟觅食栖息地利用的重要因素(Luotoet al.,2007;Saygiliet al.,2011)。食物资源对动物活动区大小产生重要影响,丰富的食物资源使得个体活动区较小,贫乏的食物资源则使得个体活动区较大(Börgeret al.,2008;Corrialeet al.,2013)。水位通过影响食物资源分布和可获得性,间接影响越冬候鸟活动区大小(刘成林等,2011;Pickens et al.,2017;吴海峰等,2018)。近些年来,由于鄱阳湖水文情势发生变化,白鹤利用农田栖息地的比例呈现不同程度增加,这得到鸟类学家的广泛关注(钟毅峰,2020;Hou et al.,2021;底明晓等,2022),但这些调查研究主要基于某个越冬阶段或某个越冬区域的调查,可能会因时间或空间局限导致对白鹤整个越冬期栖息地利用模式产生偏差,这也是与本研究结果有所不同的主要原因。
平水年,白鹤自然食物资源的可获得性是其觅食策略的主要影响因素,与平水年白鹤越冬期日间利用湿地生境比例达到89.0% ± 12.3%(n = 9)直接相关。研究发现平水年白鹤活动区面积在越冬前期显著大于中期和后期,与不同越冬阶段各水位年组移动距离变化趋势基本保持一致。Ivey 等(2015)对越冬期沙丘鹤(Grus canadensis)的移动距离和活动区大小研究发现,飞行移动距离越小,活动区面积也越小。平水年苦草等沉水植物正常生长,张笑辰等(2014)对鄱阳湖越冬食性相似的白鹤和小天鹅(Cygnus columbianus)的研究发现,越冬前期苦草冬芽分布的水深大于中期,由于白鹤形态学上的限制,导致其无法在水位较深的区域取食,而小天鹅可以在更深区域选择食物资源更丰富的觅食地。丰水年,在自然食物资源匮乏条件下,白鹤会从自然湿地转移到周边藕塘和稻田(王文娟等,2019;底明晓等,2022)。丰水年越冬期日间白鹤利用农田栖息地的比例达到50%左右,其中越冬前期和中期对农田栖息地的利用比例高于自然湿地,寻找适宜觅食地和种间竞争是影响其觅食策略的主要因素。丰水年日间白鹤提高对农田的利用率,与薛琳(2019)对不同越冬期鄱阳湖越冬白鹤日间生境选择研究结果一致。自然湿地中的苦草冬芽易消化(Foxet al.,2011),白鹤容易获取;而在藕塘中莲藕埋藏深且体积大,不易获取;稻田中稻谷易获取,但纤维素含量高,不易消化(植毅进等,2019)。尽管 如此,自然湿地中白鹤觅食时间显著高于农田生境,警戒时间也显著要低(袁芳凯等,2014;邵明勤等,2018)。因此,丰水年白鹤转移到农田生境觅食,是增加自身适合度的一种适应性行为,但仍不会放弃自然湿地觅食栖息地。
不论丰水年还是平水年,白鹤夜间主要利用自然湿地,这与胡振鹏等(2012)的观察结果一致。在丰水年的越冬中期,由于自然食物资源短缺与种间竞争,白鹤可能会增加农田觅食时间,较晚返回自然湿地,导致其夜间农田利用率显著高于平水年组。有关白鹤的食性改变是否影响其生理健康状况等还需开展相关研究,以期为白鹤越冬栖息地管理提供科学依据和支撑。
5 结论
丰水年白鹤通过增加移动距离、活动区面积以及农田栖息地利用比例来应对水位变化造成的食物资源短缺,满足能量需求来提高自身适合度。丰水年白鹤日间对农田利用率仅50%左右,夜间仍主要利用自然湿地;平水年白鹤的主要觅食地和夜宿地仍是自然湿地。因此,应加强鄱阳湖区自然湿地适应性管理,以提高不同水文条件下的湿地功能和恢复力,确保越冬白鹤的健康稳定与越冬安全。
