欧报春二型花柱的繁育特性研究
2023-10-17李金凤张晓曼
刘 柳,刘 艳,李金凤,张 璐,张晓曼
(河北农业大学 园林与旅游学院,河北保定 071000)
二型花柱是由基因控制的花的多态性,根据柱头与花药位置的不同分为长花柱和短花柱,是一种特殊的雌雄蕊异位方式[1],有利于花粉的准确传递,促进异交, 避免自花授粉和和形态内授粉。自1583年在耳报春(Primulaauricular)和粉报春(Primulafarinose)中报道二型花柱现象以来,至少在28个由动物授粉的开花植物中发现过这种特殊的花部结构[2-5],这是植物长期繁育和对环境适应性进化的必然结果。二型花柱植物通常具有自交不亲和性,但在不同植物中所表现的可育性不同。此外,不同花柱之间还常伴有花粉数量、花药大小、颜色、柱头的形状、突起高度、密度、分布的差异性[6-8],二型花柱的研究是生态学的热点之一。
欧报春(Primulavulgaris)是报春花科(Primulaceae)报春花属(PrimulaL.)多年生草本花卉。欧报春全株具香气,叶面皱,呈长椭圆形,株高10~15 cm,是报春花属中花色丰富度最高、品种相对最多,且应用最为广泛的植物,无论是在观赏价值还是抗性方面都优于其他报春,具有良好的开发前景。目前,对欧报春的研究较少,且已有的研究主要集中在花色形成机理与遗传[9-10]、栽培育种[11-14]、组培体系的建立[15-16]等方面,缺乏对欧报春二型花柱的花部形态特征、繁育特性等基础性研究。本文探究了欧报春二型花柱花部特征差异和繁育特性,为二型花柱植物自交不亲和机制的研究奠定基础,对欧报春今后育种提供依据。
1 材料和方法
1.1 试验材料
研究供试材料于2021年5月栽培于河北农业大学温室中(温度维持在20~25 ℃)。2021年11月-2022年5月选取生长良好的欧报春植株进行试验。
1.2 试验方法
1.2.1 花部形态特征观测
选择处于盛花期的欧报春长花柱和短花柱各30株,每株选择1个花序用游标卡尺分别测量花冠直径、花冠筒高度、柱头高度(从子房基部到柱头顶部距离)、花药高度(从子房基部到花药顶部距离)、花筒中部直径(花冠筒的最窄处)等形态特征, 并将测量的数据分别详细记录,单位精确到0.01 mm。
1.2.2 花粉、柱头形态扫描电镜观察
选取处于盛花期的长花柱和短花柱各10朵,分别将花粉用导电胶粘在金属台上,喷金,镀膜,于Prisma E型扫描电镜中拍照,观察和测量观察花粉赤道面观、极面观、表面纹饰和网眼形状,测量花粉极轴长(P)、赤道轴长(E)。同样步骤观察花柱,测量花柱直径和柱头乳突细胞长度,拍照。
1.2.3 花粉/胚珠比、花粉活力和柱头可授性测定
选取处于花蕾期的(雌雄蕊成熟但未散粉)长花柱和短花柱各20朵。参照Cruden[17]的方法,摘取所有花药,用1.0 mol/L HC1在60 ℃条件下水解去除花药壁,制成2.5 mL的悬浮液。先吸取0.01 mL,制片,重复记数直至观察完所有悬浮液,分别统计欧报春长花柱和短花柱的花粉量;在解剖镜下用解剖针挑破子房壁统计胚珠数量。
花粉/胚珠比=该花的花粉总量/胚珠数量。
于上午9:00,分别取开花1,3,5,7,9,11,13,15 d的长花柱和短花柱花粉,用 TTC染色法来测定花粉活力[18]。
花粉活力=具有花粉活力花粉数/观察花粉总数×100%。
将上述检测花朵的柱头用镊子取下,用联苯胺-过氧化氢反应液来检验欧报春柱头的可授性[19]。参考孙建等[20]的分级标准,1级:少量气泡,可授性低,标记为+/-;2级:气泡较多,可授性中等,标记为+;3级:有大量气泡,可授性高,标记为+/+。
1.2.4 杂交指数估测和杂交亲和性测定
依Dafni[21]的方法,通过对欧报春成熟情况,性器官空间位置和花序直径来估算欧报春杂交指数。
对欧报春的栽培种群进行人工授粉,设置6种授粉组合:(1) 同型自花授粉,每日上午8:00,分别选择当天开放长花柱和短花柱的花朵,在未开放前套袋,7 d后揭开袋子,用洁净的刷子蘸取同一朵花的花粉进行授粉并套袋,标记具体的授粉时间。(2)异株同型授粉,每日上午8:00,分别选择当天开放长花柱和短花柱的花朵,未散粉前对母本去雄并套袋。父本花粉,选择不同株的同型植株,未开花前进行套袋,待开花7 d后收集花粉用洁净的刷子对母本进行授粉并套袋,标记具体的授粉时间和父母本。(3)异型花间授粉,每日上午8:00,分别选择当天开放长花柱和短花柱的花朵,未散粉前对母本去雄并套袋。父本花粉,选择异型植株,未开花前进行套袋,待开花7 d后收集用洁净的刷子对母本进行授粉并套袋,标记具体的授粉时间和父母本。同株长花柱记作P,短花柱记作T,不同株长花柱记作P1,短花柱记作T1。每种处理做50个重复,结实后在解剖镜下统计饱满种子数。
为了观察6种授粉组合授粉后花粉的萌发、花粉管的生长和进入子房的情况,于授粉后1 h,2 h, 4 h, 6 h,8 h,12 h,18 h,24 h,48 h,72 h取下各组合的雌蕊,装入FAA溶液的离心管中处理24 h,放入70%的乙醇中进行储存备用。取出雌蕊,用蒸馏水冲洗3~4次,将长花柱、短花柱雌蕊分别放入盛有2 mol/L、4 mol/L的NaOH溶液的离心管中,分别软化60 min,30 min后取出,用蒸馏水冲洗,放入0.1%水溶性苯胺蓝溶液中进行染色。采用常规制片法制片[22],用荧光显微镜观察,测量花粉管长度和花柱长度,计算相对位置。
1.3 数据处理
试验数据利用 SPSS 22.0进行数据处理分析,用Duncan法比较不同处理间差异,采用Excel汇总数据和制作图表。
2 结果与分析
2.1 花部形态特征观测
通过观察花部结构的发现,欧报春为典型的二型花柱植物。长花柱柱头高于花药,在花冠口处可见(图1,A、B);短花柱柱头低于花药,雄蕊在花冠口处可见(图1,C、D)。
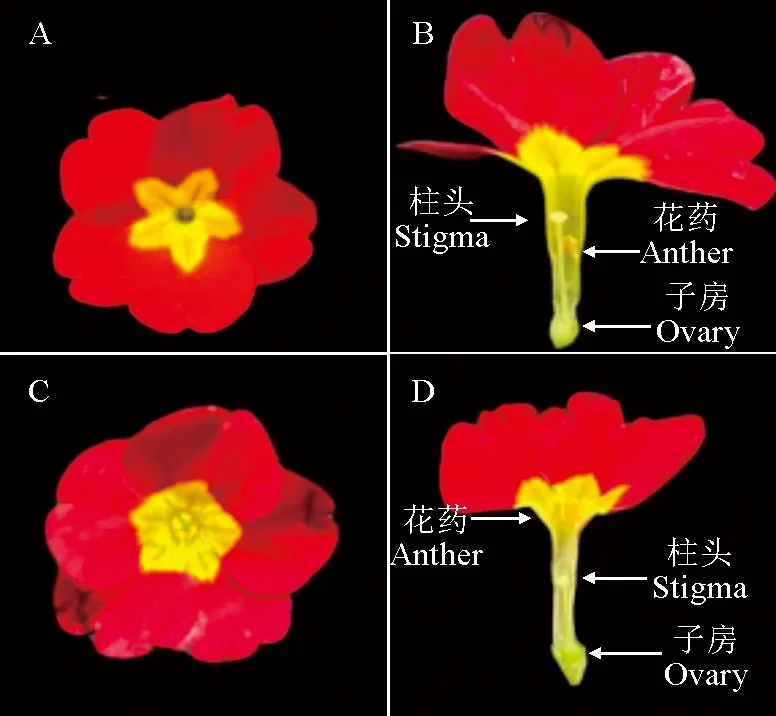
A. 长花柱花部形态;B.长花柱剖开图;C.短花柱花部形态;D.短花柱剖开图
由表1可知,长花柱的花冠直径为3.86 cm,短花柱为3.74 cm, 长花柱大于短花柱0.12 cm, 差异不显著;长花柱的花冠筒长度为1.43 cm,短花柱为1.55 cm,长花柱小于短花柱0.12 cm,差异显著;长花柱柱头高度为1.28 cm,短花柱为0.79 cm,长花柱大于短花柱0.49 cm,差异极显著;长花柱花药高度为0.83 cm,短花柱为1.35 cm, 长花柱小于短花柱0.52 cm,差异极显著;长花柱花筒中部直径为0.37 cm,短花柱为0.36 cm, 长花柱大于短花柱0.01 cm,差异显著。综上所述,长花柱和短花柱的相同花部之间存在差异。
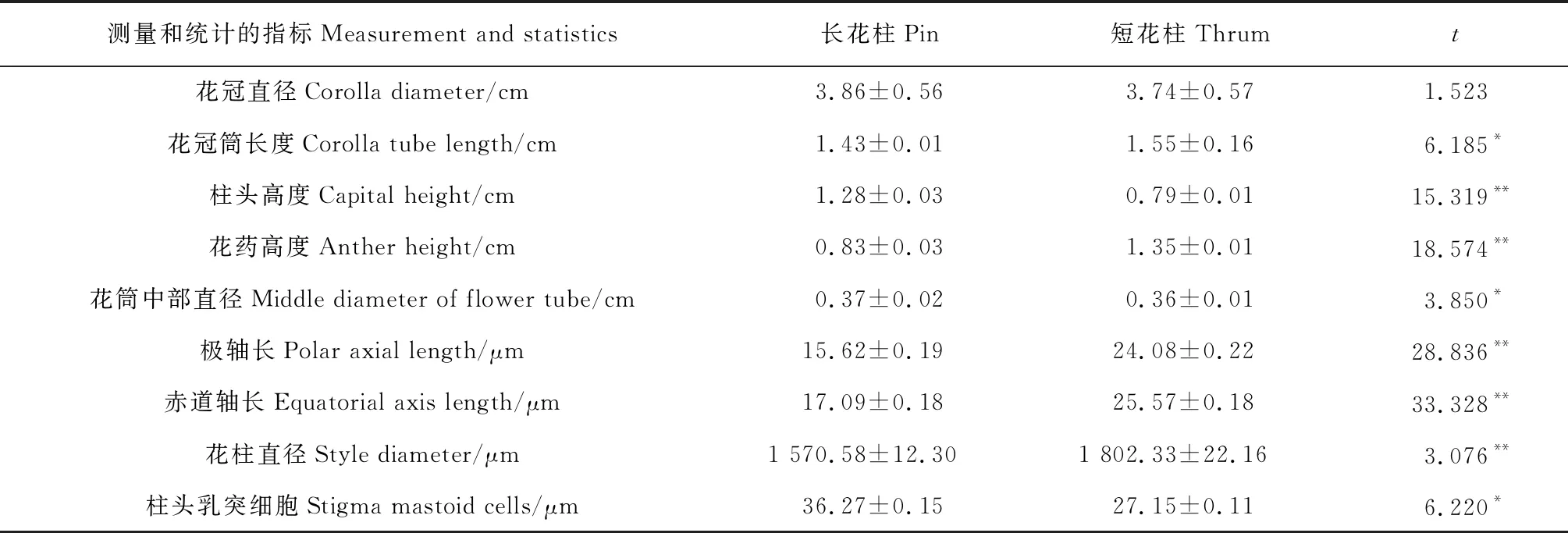
表1 长花柱和短花柱的花部形态特征
2.2 花粉、柱头观察
欧报春的长花柱和短花柱花粉粒较小,并都以单粒的形式存在,形状均为超长球形,因而不易从外形上将2种花粉区别开;花粉赤道面观均为近椭圆形(图2,A、C);极面观均为近圆形(图2,B、D);外壁纹饰为网状雕纹;网眼形状为密网纹;具有萌发沟;长花柱的极轴长和赤道轴长分别为15.62,17.09 μm,短花柱相对应各部分别为24.08,25.57 μm,长花柱均小于短花柱,呈极显著差异(表1)。

A. 长花柱花粉, 标尺=5 μm;B.长花柱花粉, 标尺=10 μm;C.短花柱花粉, 标尺=10 μm;D.短花柱花粉, 标尺=10 μm;E.长花柱, 标尺=500 μm;F.长花柱, 标尺=200 μm;G. 短花柱, 标尺=500 μm;H.短花柱,标尺=200 μm。
长花柱的花柱直径小于短花柱,呈极显著差异(表1);欧报春柱头表面未发现黏性物质,且分泌物较少,不易捕获花粉(图2,E、G);柱头与花柱连接孔处较光滑,长花柱外壁为细微的网状结构,短花柱为粗糙的网状结构;柱头表面均有乳突,长花柱柱头的乳突细胞长度显著大于短花柱(表1),因而容易固定短花柱所产生的较大花粉,短花柱柱头的乳突较短(图2,F、H),从而更容易固定长花柱花所产生的较小花粉。
2.3 花粉/胚珠比、花粉活力和柱头可授性测定
欧报春长花柱单花花粉总量为18 425粒,短花柱为14 918粒,花粉数量差异不显著(表2);长花柱胚珠数量为48.00,短花柱为44.08,胚珠数量差异不显著;长花柱P/O比为384.20,短花柱为369.70,按Cruden的标准,欧报春的繁育系统属于兼性异交类型。
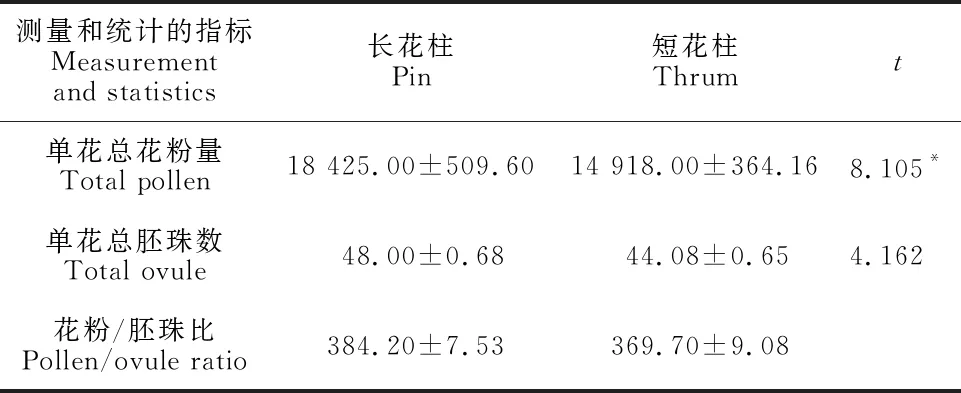
表2 花粉-胚珠比值
由表3可知,不同开花时期长花柱和短花柱的花粉活力存在一定差异,花粉活力随时间的变化相似。欧报春长、短花柱的花粉活力开花后均逐渐升高,在第7天时达到最高,分别为73.06%和79.90%,然后再降低,第15天时花粉失去活力。不同开花时期长、短花柱的柱头可授性也存在一定差异,总体呈先升后降的趋势。开花第1天,长花柱已具备中等程度的可授性,而短花柱可授性低;开花第7~11天时,长、短花柱均具备高可授性;开花第15天,长、短花柱的可授性均由高降为低。综上所述,长花柱和短花柱的花粉活力和柱头可授性能在较长时间内维持较高活力。

表3 欧报春花粉活力与柱头可授性检测结果
2.4 杂交指数估测和杂交亲和性测定
根据Dafni 对杂交指数估算标准(表4),长花柱和短花柱的单花直径都远大于6 mm,赋值为3;花药开裂与柱头具有可授性之间不同步,赋值为1; 长花柱的花药与柱头在空间上有隔离,不能接触,赋值为1,杂交指数值为5; 短花柱的花药与柱头虽然在空间上有隔离,但柱头位于花药下方,所以花药与柱头可能会有接触,赋值为0,杂交指数值为4,因此根据Dafni的标准,欧报春的繁育系统类型可定为异交,部分自交亲和,需要传粉者。

表4 欧报春杂交指数估测
由表5可知,不同授粉组合均可结实,但结实数之间存在差异,具体表现为短花柱为母本的异型花间授粉>长花柱为母本的异型花间授粉>长花柱自花授粉和长花柱异株同型授粉>短花柱自花授粉和短花柱异株同型授粉>长花柱和短花柱的对照。综上所述,异型花间授粉的结实数明显高于自花授粉和异株同型授粉,短花柱为母本的异型花间授粉组合亲和性优于其他组合,由对照组可知,在无传粉昆虫的温室中欧报春能少量结实。

表5 人工授粉结实数
花粉管观察试验结果表明,长花柱、短花柱异花授粉无论时间长短花粉管都顺利到达子房,长花柱为母本的异型花花间授粉花粉管在72 h内到达子房(图3,A),短花柱为母本的异型花间授粉花粉管在48 h内到达子房(图4),并与胚珠结合(图3,B),花粉管的生长速度最快;长花柱自花授粉中部分花粉管72 h可到达子房(图3,C),长花柱异株同型授粉仅有少量花粉管到达花柱与子房的连接处;短花柱自花花粉异株和同型花粉萌发情况较差(图3,D),但花粉萌发后的生长速度与异型花粉授粉无显著差异。综上所述,欧报春存在自交不亲和,长花柱的自交不亲和性低于短花柱。
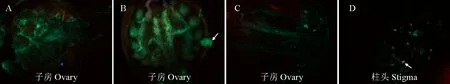
B图中箭头所指位置为胚珠;D图中箭头所示为花粉萌发。
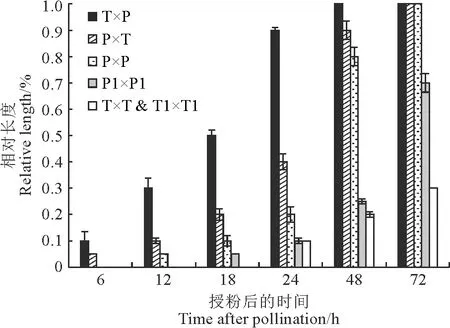
T×P.短花柱×长花柱;P×T.长花柱×短花柱;P×P.长花柱×同株长花柱;P1×P1.长花柱×不同株长花柱;T×T & T1×T1.短花柱×同株短花柱&短花柱×不同株短花柱。
3 讨 论
二型花柱植物在形态上的差异除了雌雄蕊高度外还有花部特征的多样性[23]。欧报春长花柱的花冠直径和花筒中部直径大于短花柱,这通常被认为是一种机制,以补偿花粉粒到达长花柱的偏差,迫使长花柱比短花柱吸引更多的传粉者[24]。欧报春长花柱的花柱高度与短花柱的花药高度相近,与买金梅对刺叶彩花(Acantholimonalatavicum)的研究结果[25]一致,究其原因,应是避免雌雄蕊的相互干扰,同时促进异型花间的授粉。研究发现欧报春长花柱的花粉和花柱形态与短花柱不同,长花柱柱头的乳突较短花柱的长,容易固定短花柱产生的较大的花粉,短花柱柱头的乳突较短,从而更容易固定长花柱花产生的较小的花粉,与冯建菊等人对新疆紫草(Arnebiaeuchroma)和卢婉佩对小报春(Primulaforbesii)的研究结果[26-27]一致。花粉形态和柱头乳突的差异叶会导致花粉粘附能力的不同,从而影响植物的繁殖特性,如对二型花柱植物白花丹(Plumbagozeylanica)的花粉外壁与柱头角质层形态研究发现,其会影响物理接触和化学过程[28]。此外,有研究表明不亲和反应与花粉和柱头结构和化学成分有关[29]。
传粉在对植物的繁育中有重要意义,花粉活力和柱头可授性是影响传粉成功的重要因素[30]。TTC染色法具有快速、准确、灵敏度高等特点,联苯胺-过氧化氢法测定柱头可授性在报春中被广泛应用[31]。柱头是接受花粉的部位,其活性影响花粉的附着、萌发和花粉管伸长。当花粉活力与柱头可授性都处于高活力时,是收集花粉的最佳时期,也是最佳的授粉时段,有利于植物顺利进行授粉受精过程。根据花粉/胚珠比、杂交指数估测结果来看,欧报春的繁育系统为异交和自交都有。虽有研究表明二型花柱植物往往遵循的严格自交不亲和繁育系统[32-34],与本试验结果不同。但也存在二型花柱的植物具有部分自交亲和[23,35],如紫草科的一些品种通常具有自交亲和性[36],四季报春(Primulaobconica)[31],翠南报春(Primulapatens)[37]也存在自交亲和性。同一属中花柱异型的植物的繁育特性可能也不同,与所处环境有关,如在水仙属中比利牛斯山黄水仙(Narcissusalbimarginatus)和密花兜被(N.calcicola),是自交不亲和的, 而非洲之恋水仙(Narcissusrupicola)是自交亲和的,是因为该物种是分布于地中海的高海拔地区,环境恶劣, 传粉昆虫甚少, 在此环境下植物失去自交不亲和系统,来确保繁殖成功[38]。欧报春这种自交亲和的繁育系统可能是与其生活史相适应的[39],是对自身的一种生殖保障。
杂交授粉试验表明长花柱的自交不亲和性显著低于短花柱,这与Massinga等人的研究结果,存在部分报春花属植物的长花柱具有一定的自交亲和性,而短花柱表现为严格的自交不亲的一致[8],其原因还需进一步探究。荧光显微观察表明,无论是自花花粉、异型花粉还是异花花粉均能在长花柱的柱头上萌发,只是异型花粉的萌发速度快于其他类型,而短花柱为母本的植株无论自交还是同型异株授粉更多的表现为自交不亲和,花粉无法在柱头上萌发,与邵剑文等人对安徽羽叶报春的研究结果[40]一致。这是因为长花柱依靠控制花粉管的生长对花粉进行选择和识别,短花柱于花粉在柱头表面萌发的阶段对花粉进行识别。Tremayne对报春花属植物研究发现自交阻碍发生在花柱中的类型比自交阻碍发生在柱头表面的类型更容易产生种子[41],本试验结果进一步验证了这一结论。
