荒漠早春开花植物囊果草大小孢子发生及雌雄配子体发育
2023-10-17黄代红吉乃提汗马木提
张 芳,黄代红,吉乃提汗·马木提,2*
(1 新疆农业大学 生命科学学院,乌鲁木齐 830052;2 新疆农业大学 草业学院, 西部干旱区草地资源与生态教育部重点实验室,乌鲁木齐 830052)
植物生殖生物学是研究植物有性繁殖过程的一门前沿学科,其涉及花芽分化、雌雄性器官发生与发育、传粉受精及胚胎发育等内容[1-3]。研究植物的生殖生物学特性,不仅对揭示其演变及进化轨迹、阐明个体发育及物种间的亲缘关系具有重要作用[4],还是开展杂交育种或系统学研究的前提和基础[5]。
小檗科(Berberidaceae)共有17属、700余种[6-8],主要分布于北温带和亚热带高山地区[7]。该科被划分为鬼臼亚科(Podophylloideae)、小檗亚科(Berberidoideae)和南天竹亚科(Nandinoideae)3个亚科[8-10]。形态解剖学和分子系统学研究证明该科为单系[10-13]。冯旻等[14]以及Liu等[15]分别对该科的南天竹(Nandinadomestica)和牡丹草(Gymnospermiummicrorrhynchum)的花器官发生及其系统学意义进行了研究,讨论了花器官的三基数祖征性状并揭示了牡丹草花器官偏离祖先模式的二基数演化特征。黄衡宇等[16]以及马绍宾等[17]对鬼臼亚科胚胎学与系统学的研究表明,鬼臼亚科为1个自然类群,确定了该亚科各属间的演化关系。张明英等[18]认为小檗亚科和南天竹亚科存在共衍征,将其与鬼臼亚科区分开来,并支持了小檗亚科与南天竹亚科的姐妹关系。这些研究为小檗科植物的生殖生物学研究积累了基础资料。
囊果草属(Leontice)隶属于小檗科南天竹亚科囊果草族(Leonticeae)[19]。该属约3~4种,主要分布于北温带地区,中国仅1种,产于新疆[7]。囊果草属通常具块状根,茎直立,草质,不分枝,全株无毛[7]。Nickol[20]认为,兰山草属(Ranzania)、二叶鲜黄连属(Jeffersonia)、山荷叶属(Diphylleia)和鬼臼属(Podophyllum)等均为囊果草属的姊妹属,且因牡丹草属(Gymnospermium)的果实类型没有可靠的形态学证据支撑,则将其归并在囊果草属中。但Liu等[15]基于牡丹草属和囊果草属花粉外壁纹饰的不同,提出牡丹草属应从囊果草属中独立出来。
囊果草(LeonticeincertaPall.)是仅分布于哈萨克斯坦和中国新疆的小檗科囊果草属多年生早春开花植物。在新疆,该物种仅带状分布于天山北坡荒漠和低山山坡[7],具有独特的观赏价值,是一种值得开发利用的野生花卉,且具有较高的饲用价值和生态价值[21-23]。
然而,由于囊果草自然种群分布区有限,分布面积狭窄,生长环境恶劣,且各种家畜均喜采食等影响而面临濒危状态[23]。在《世界自然保护联盟濒危物种红色名录》(IUCN)中,该物种被评估为近危种(NT)[7]。
有关囊果草的研究主要集中在形态特征[7]、染色体核型分析[24]、种群分布及其生态适应[23]、种子休眠与萌发特性[25]及繁殖保障机制[26]等方面,而有关该物种的生殖生物学研究鲜见报道。因此,本文以荒漠早春开花植物囊果草为研究材料,采用石蜡切片技术观察其大/小孢子发生与雌/雄配子体发育的胚胎学特征,以期明确囊果草有性生殖过程,不仅为进一步分析该物种的致危原因以及指定合理的保护策略提供理论依据,还可为小檗科植物的系统学研究积累基础资料。
1 材料和方法
1.1 取样地点与研究材料
囊果草(L.incerta)采自新疆沙湾县安集海镇南部低山山坡。该地点地理坐标为85°30′04.6″E、44°36′08.6″N,海拔600 m左右,属温带大陆性干旱荒漠气候,土壤为淡栗钙质荒漠土[27]。年均气温约6.5 ℃,年最低气温(1月)可达-28 ℃,年最高气温(7月)可达37 ℃,年均降水量为131.8 mm,其中约65%的降水发生在春季和夏季(数据来源于中国气象局国家气象信息中心, 2020-2021年)。年蒸发量为1 237~1 770 mm,年均日照时数可达2 200~3 200[27-28]。
囊果草株高5~15 cm,直立总状花序,每年3月下旬至4月上/中旬快速萌动开花,5月中旬果实成熟[7,26]。6月初,植株地上部分凋亡,属于典型的多年生早春短命植物。
1.2 研究方法
于2021年3月5日至4月16日、9月15日至11月25日,分别在自然居群中对该物种不同发育时期的花芽及开放的花朵进行采样。每7 d取1次,按照材料大小不同,分别将其固定在10 mL和50 mL FAA (70%乙醇∶甲醛∶冰乙酸 = 18∶1∶1, 体积比)固定液中,置4 ℃冰箱保存备用。
在室内将已固定的材料在解剖镜下进行解剖,分别剥离出完整的雌、雄蕊。采用石蜡切片法[29]:将材料抽气后在植物软化液中软化24 h,成熟花药用番红—固绿双重染色外,其他材料均利用Gill改良苏木精法[30]整染2 d,水洗返蓝(成熟花药用番红—固绿双重染色),按(70%、75%、80%、90%、95%、100%、100%)乙醇梯度逐级脱水,前5级均1 h,100%乙醇40 min;体积比1∶1的无水乙醇和二甲苯透明1 h,纯二甲苯透明2次,均45 min;将透明好的材料放置于体积比约1∶2的二甲苯和液体石蜡混合液中37 ℃过夜。在56~60 ℃浸蜡2~3 d,每6 h更换1次过滤干净的蜡液,最后进行包埋和修块,用Leica RM 2126RT切片机切片(蜡带厚度8~10 μm)、展片;二甲苯脱蜡、透明,树胶封片,在光学显微镜下观察并拍摄。
2 结果与分析
囊果草具2片基生叶,为总状花序,每个花序上约8~15朵小花(图1, A)。花序上的开花顺序为自下而上。其花为两性花,黄色,每朵花具6枚花萼,呈卵圆形,6枚花瓣特化为蜜腺状,6枚雄蕊离生,单心皮雌蕊,花萼明显长于花瓣(图1, B)。在花萼背面具有蓝紫色斑块(图1, C)。花朵较小,花瓣特化为蜜腺状,呈倒卵形,与雄蕊对生,雌蕊发育为囊状结构,具短花柱,柱头平截(图1, D、E)。成熟花药的开裂方式为瓣裂(图1, F)。果实呈囊泡状紧密排列在果序上,果皮不开裂,4枚种子基生。

A. 植株和总状花序; B. 单花; C. 箭头示花萼背面具蓝紫色斑块; D. 花部形态, 示雄蕊与特化花瓣; E. 侧面观, 示囊状单雌蕊; F. 箭头示花药瓣裂. b. 苞片; i. 花序; l. 叶; p. 花瓣; pi. 雌蕊; s. 花萼; st. 雄蕊; sti. 柱头; sty. 花柱. B, D-F. 标尺 = 2 mm。
2.1 小孢子发生与雄配子体发育
2.1.1 花药壁的发育
囊果草雄蕊具4个小孢子囊,两侧的小孢子囊由药隔相连。花药原基由雄蕊原基顶端分化形成。花药原基最外层为1层表皮细胞,内侧主要为分生组织,表皮内侧4个角隅处各有分裂产生的孢原细胞(图2, A)。孢原细胞经1次平周分裂形成2层细胞,向外为初生壁细胞,即周缘细胞,向内形成体积较大的初生造孢细胞。周缘细胞进行平周和垂周分裂,由外向内逐渐产生4层细胞,分别为1层药室内壁、2层中层和1层绒毡层;造孢细胞经多次有丝分裂形成小孢子母细胞,早期的绒毡层细胞为单核,胞质浓厚(图2, B-D)。花药壁分化完全,中层细胞呈扁平状,最内层的绒毡层细胞不规则,细胞体积大,原生质浓,具双核或多核(图2, E)。

A. 花药原基横切面, 示孢原细胞时期; B. 周缘细胞进行平周分裂; C. 幼小花药的横切面, 示4个小孢子囊的分化; D. 发育中的花药壁及早期小孢子母细胞; E. 箭头示双核或多核绒毡层细胞; F. 花药壁和游离小孢子; G. 绒毡层细胞开始解体; H. 纤维状加厚的药室内壁; I. 成熟花药的横切面, 示纤维状加厚的药室内壁和成熟花粉粒. aw. 花药壁; en. 药室内壁; ep. 表皮; ml. 中层; mi. 小孢子; ta. 绒毡层; fe. 纤维状加厚的药室内壁; pg. 花粉粒. A-D. 标尺=100 μm; E-H. 标尺=50 μm; I. 标尺=200 μm。
因此,囊果草的花药壁具2层中层,依据Davis[31]对被子植物花药壁发育划分标准,该物种花药壁发育为基本型。
小孢子四分体至游离小孢子时期,药室内壁逐渐加厚,中层细胞发生解体,发达的绒毡层细胞开始出现解体(图2, F)。小孢子单核靠边期,绒毡层细胞不断为小孢子提供营养,细胞界限不明显,细胞核消失,逐渐原位解体被小孢子吸收,故为腺质型绒毡层(图2, G)。随着早期花粉粒的发育,药室内壁进一步径向延长,最内1层中层细胞被吸收,绒毡层完全解体(图2, H)。成熟花药时期,花药壁仅剩纤维状加厚的药室内壁和残留的表皮细胞,绒毡层和中层细胞完全解体,表皮细胞未见明显加厚(图2, I)。
2.1.2 小孢子发生
囊果草幼小花药表皮内侧的孢原细胞经平周分裂,形成的造孢细胞体积大,近椭圆形,核仁明显,细胞质浓厚,无明显液泡(图3, A-C)。小孢子母细胞呈圆球形,细胞体积大,胞质浓厚,细胞核大而核仁明显,无明显液泡,核仁与染色质形成染色较深的圆球体(图3, D)。分化初期,圆球形的小孢子母细胞被胼胝质壁包围,进入减数分裂期。小孢子母细胞在减数第1次分裂前期,绒毡层细胞开始发生核分裂(图3, E)。随后,进入减数第1次分裂中期,此时染色体排列在赤道板附近(图3, F)。小孢子母细胞减数第2次分裂后期,4个细胞核由纺锤丝相连接(图3, G)。第1次核分裂过程不伴随胞质分裂,直到减数第2次分裂末期,4个子细胞核形成后,细胞质开始分裂,小孢子四分体呈四面体型排列(图3, H、I)。可见,囊果草小孢子母细胞减数分裂过程中胞质分裂为同时型,小孢子四分体排列为四面体型,其壁呈向心式发生。
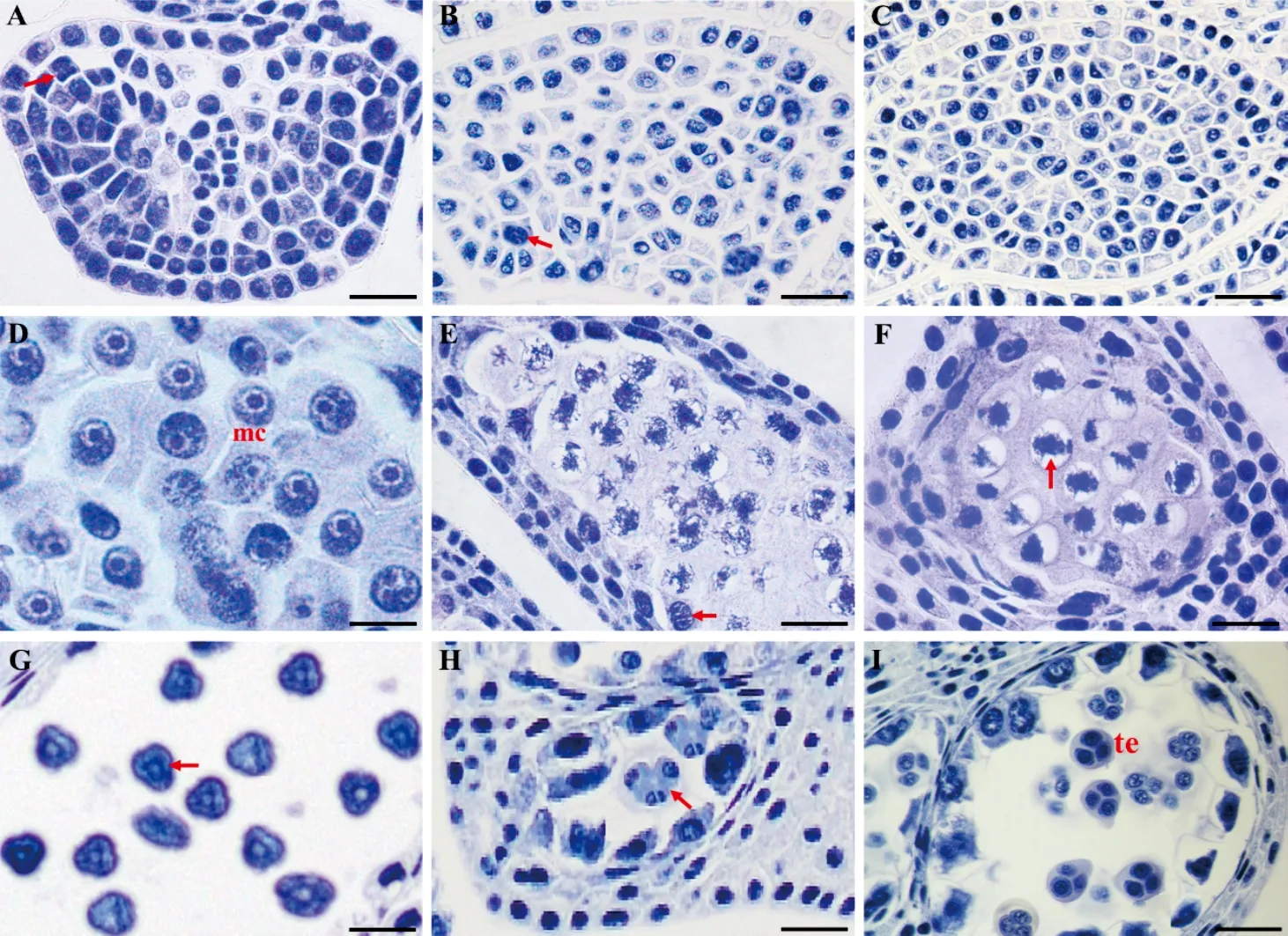
A. 孢原细胞平周分裂; B, C. 造孢细胞时期; D. 小孢子母细胞; E. 花药纵切面, 示小孢子母细胞减第Ⅰ次数分裂前期绒毡层细胞发生核分裂; F. 小孢子母细胞减数第Ⅰ次分裂中期; G. 小孢子母细胞减数第Ⅱ次分裂后期; H. 小孢子母细胞减数第Ⅱ次分裂末期; I. 四分体时期. mc. 小孢子母细胞; te. 四分体. A-C. 标尺 = 100 μm; D, G. 标尺 = 35 μm; E, F, H, I. 标尺 = 50 μm。
同一药室及同一花药的不同药室间小孢子母细胞减数分裂基本同步。在小孢子母细胞减数分裂过程中,伴随着胼胝质壁的发生,其始终包围在小孢子外。小孢子减数分裂形成的四分体亦被胼胝质壁所包围。
2.1.3 雄配子体的发育
小孢子母细胞在减数分裂完成后,小孢子四分体的胼胝质壁开始溶解,四分体小孢子逐渐被分离开,形成游离的单核小孢子(图4, A、B)。刚释放的单核小孢子又叫单核花粉粒。其细胞呈椭球形,细胞壁薄,胞质浓厚,细胞核较大且位于细胞中央(图4, C)。此后,单核花粉粒不断从绒毡层细胞中获取营养,细胞质明显液泡化,逐渐形成中央大液泡,细胞核被挤向花粉一侧,进入单核靠边期(图4, D)。

A. 胼胝质开始溶解; B. 从四分体中释放的游离小孢子; C. 单核居中期, 花粉细胞中具大液泡; D. 单核靠边期, 花粉细胞具有大液泡; E. 早期单细胞花粉粒; F. 单核花粉粒有丝分裂期; G, H. 早期2细胞花粉, 1个大的营养细胞, 箭头示1个小的靠边生殖细胞; I. 成熟花粉粒, 箭头示3细胞型花粉. A~D. 标尺 = 50 μm; E~I. 标尺 = 25 μm。
随着单核花粉粒不断生长,小孢子核在靠近花粉壁位置发生不均等有丝分裂(图4, E、F)。经有丝分裂后形成2个细胞核,贴近花粉壁的为生殖核,靠近中央液泡的为营养核(图4, G、H)。成熟花粉粒多为2细胞型,偶见3细胞型,花粉近球形(图4, I)。花药成熟后,花粉囊开裂,成熟花粉粒散出。
2.2 大孢子发生与雌配子体发育
2.2.1 大孢子发生
囊果草具单雌蕊,由1心皮组成,子房1室,为基生胎座,胚珠4枚(图5, A)。胚珠原基逐渐分裂形成指状结构,发育为珠心组织,珠心表皮细胞下出现孢原细胞,其体积大而胞质浓(图5, B)。孢原细胞经平周分裂,形成周缘细胞和造孢细胞,前者分裂形成的细胞参与珠心组织的生长发育;造孢细胞进一步发育为大孢子母细胞,其位于远离珠孔端位置,细胞呈圆球形,细胞核大且核仁明显(图5, C)。随着大孢子母细胞不断分化,内、外珠被逐渐形成并包围珠心组织。此时,直立胚珠逐渐向一侧弯曲与珠柄平行,呈倒生生长(图5, D、E)。周缘细胞数目增多,大孢子母细胞被数层周缘细胞与珠心表皮隔开(图5, F)。因此,囊果草胚珠发育为厚珠心胚珠。囊果草大孢子母细胞经减数分裂形成直线形排列的大孢子四分体(图5, G、H)。随后,四分体靠近珠孔端的3个大孢子相继退化,仅合点端1个大孢子优先分化,其体积增大,细胞核明显(图5, I)。珠孔端的3个大孢子逐渐退化消失,合点端大孢子逐渐向珠孔端移动,发育为功能大孢子(图5, J)。功能大孢子进一步发育,形成了单核胚囊(图5, K、L)。
2.2.2 雌配子体的发育
单核胚囊经1次有丝分裂,形成2核胚囊。刚形成的2核胚囊中,2个核位于胚囊稍中央位置,核两端液泡化明显,此时,外珠被分化速度加快,包裹内珠被和珠心组织(图6, A、B)。胚囊腔逐渐增大,2核逐渐向胚囊两端移动,此时,2核胚囊开始进行第2次有丝分裂,形成4核胚囊(图6, C)。胚囊腔的液泡逐渐增大,内、外珠被分化完全。随后,4核胚囊发生第3次有丝分裂,形成8核胚囊(图6, D)。同时,胚囊发生细胞质分裂,形成7细胞8核的成熟胚囊。其中,珠孔端的3个核形成卵器,2个极核靠近合点端,3个反足细胞紧密排列在合点端,其细胞核明显,细胞质浓,分化完全的成熟胚囊发育为典型的单孢蓼型(图6, E)。囊果草成熟胚囊珠孔端的卵器由1个卵细胞和2个助细胞组成,呈“品”字形排列(图6, F)。助细胞分化早期,其胞质浓厚,细胞内未见明显液泡化,细胞周围具发达的丝状器,其染色较深,呈漏斗状排列(图6, G)。随着胚囊不断发育,合点端的3个反足细胞逐渐退化消失,上下极核体积增大,逐渐融合,形成2核中央细胞(图6, H、I)。

A. 单核胚囊第Ⅰ次有丝分裂; B. 2核胚囊; C. 4核胚囊; D, E. 8核蓼型胚囊, 胚囊内合点端的反足细胞开始逐渐退化; F. “品”字形卵器, 示1个卵细胞和2个助细胞; G. 助细胞早期, 示丝状器; H, I. 反足细胞退化, 2个极核逐渐融合. tes. 2核胚囊; fes. 4核胚囊; ac. 反足细胞; ec. 卵细胞; pn. 极核; sy. 助细胞; fa. 丝状器。A-C, F-I. 标尺 = 50 μm; D, E. 标尺 = 100 μm。
3 讨 论
有性生殖过程成功进行对植物的生命延续和种群扩增具有重要意义[32]。生殖过程的任何一个环节出现异常,都将不同程度地导致生殖败育,因此,了解大/小孢子发生与雌/雄配子体发育规律对于植物有性生殖能否顺利完成具有重要的作用[33-35]。本研究结果表明,囊果草花药具4个花粉囊,花药壁由5层细胞组成,由外至内分别为1层表皮、1层药室内壁、2层中层和1层绒毡层,其药壁发育为基本型,这与Davis[31]的描述一致。其药室内壁发生纤维状加厚,腺质型绒毡层细胞具2核或多核现象。花药成熟前,同侧两个花粉囊间的隔膜退化并消失。在解剖过程中发现,花药壁的不同发育阶段均未出现异常现象。这些结果与同科植物南方山荷叶(Diphylleiasinensis)花药壁的发育过程相似[36]。
花粉是植物遗传信息的重要载体之一,具有较强的遗传稳定性[37]。开花植物的成熟花粉类型代表整个属(科)的进化特征[31, 38]。据报道,开花植物中系统发育较原始物种均具有2细胞型花粉,而3细胞型花粉演化程度较高,被认为是开花植物对不同环境的适应性演化[39-40]。在小檗科植物中2细胞型花粉出现在十大功劳属(Mahonia)[41]、山荷叶属[36]、八角莲属(Dysosma)和桃儿七属(Sinopodophyllum)[16]等大多数物种中,偶见3细胞型,且桃儿七属亦存在单细胞型花粉。在本研究中,囊果草成熟花粉多为2细胞型,偶见3细胞型。这一结果与上述已报道的属的花粉类型相吻合,反映了小檗科各属种间系统发育的一致性,进而体现了囊果草雄配子体发育过程的稳定性。
囊果草的大孢子发生与同科植物齿蕊十大功劳的发育特征表现出一致性[41],即具有倒生胚珠;胚珠具双珠被和厚珠心;1个功能大孢子发育为7细胞8核的单孢蓼型胚囊;珠孔端助细胞早期形成发达的漏斗形丝状器。研究表明,倒生胚珠、双珠被、厚珠心及蓼型胚囊均为较原始的胚胎学性状[42],且蓼型胚囊被认为是被子植物最常见的胚囊类型[43],且刚分化的助细胞丝状器发达,成熟胚囊的丝状器逐渐消失,在胚囊发育过程中,丝状器可能承担营养转输作用[44]。由此推测,囊果草的胚珠和胚囊发育过程均表现出明显的原始性状,体现了其胚胎发育的保守性。
生殖生物学特征在植物系统进化过程中属于比较保守的性状,是不同类群间亲缘关系、系统进化进程的重要判断依据[45-46]。有性繁殖过程的顺利进行则有效保证了开花植物的结实结籽率[47]。囊果草的大/小孢子发生和雌/雄配子体发育过程正常,其发生、发育过程多表现出较原始的胚胎学性状。可见,囊果草是小檗科的较原始物种。
4 结 论
在形态结构上,囊果草为两性花,雌蕊存在囊泡状膨大现象。本研究结果表明,囊果草大/小孢子发生与雌/雄配子体发育顺利完成,即花药和胚囊发育过程正常,不仅可作为授粉植株,也可作为优良的杂交母本。其发生/发育过程与小檗科已报道的其他物种具有相似性,表现出较原始的发育特征。该物种孢子发生与配子体发育的生殖生物学特征可为进一步对其进行杂交育种或人工繁育提供基础资料,还可为该物种及其近缘科属的系统学研究提供重要信息。
致谢:感谢陈可为、刘帅、杨帆和薛肖兰帮助采集试材;感谢孙博洋帮助解决试验过程中遇到的问题;感谢薛肖兰和杨帆在试验过程中的帮助。
