酵母培养物和活性酵母对湖羊血清生化指标、瘤胃发酵及瘤胃细菌结构与组成的影响
2023-10-17武奥林蔡大亮彭清洁谢绿绿戴晋军
武奥林, 蔡大亮,, 彭清洁, 周 迪,谢绿绿,, 张 彦,, 戴晋军,
(1.农业微生物资源发掘与利用全国重点实验室,湖北宜昌 443003;2.酵母功能湖北省重点实验室,湖北宜昌 443003)
酵母源物质已被证实对反刍动物具有积极作用, 成为替代抗生素的优选添加剂(Mezzetti 等,2022;Brewer 等,2014)。 酵母源物质主要包括酵母菌、 酵母培养物、 酵母水解物和酵母微量元素等,随着研究的深入,很多研究开始聚焦于酵母源物质对瘤胃的发酵特性、 菌群结构(Centeno 等,2023;Ferraretto 等,2012) 以及组学关联性等(Wang 等,2023)。 研究表明, 酵母源物质可以对纤维分解菌 (Pinloche 等,2013)、 淀粉利用菌(Jiang 等,2017)和乳酸盐利用菌(Rossi 等,2004)等瘤胃细菌产生影响(Rossi 等,2004),且不同的添加剂量会导致不同的菌群结构, 进而产生不同效果(Jiang 等,2017)。
反刍动物的生长依赖宿主与瘤胃微生物的共生作用, 微生物可以为宿主提供能量、 蛋白质、维生素等多种营养。 高达90%到达小肠的蛋白质和50%的宿主能量需求是由网状瘤胃微生物细胞提供的(Boyd 等,1991)。 在日粮中,添加一些酵母菌或酵母培养物, 可以通过增加饲料摄入量和饲料消化率来提高动物的生产性能(何文娟等,2023;饶时庭等,2020)。 在某些条件下,它们还可能改变瘤胃发酵模式,降低乙酸/丙酸比例, 提高氨态氮和TVFA 含量 (Wang 等,2023; 任胜男等,2020), 提高血液抗氧化能力(王晓芳等,2022;Jia 等,2018), 增加纤维分解菌、淀粉利用菌和乳酸盐利用菌的总活菌数,且不同的添加剂量会导致不同的菌群结构, 进而产生不同效果(Jiang 等,2017;Pinloche 等,2013;Rossi 等,2004)。 本试验旨在研究酵母培养物和活性酵母对湖羊血清生化指标瘤胃发酵及瘤胃细菌结构与组成的影响。
1 材料与方法
1.1 试验设计 试验选取遗传背景一致,体重相近[(25±2.5)kg]的湖羊公羔72 只,随机分为3 组,每组4 个重复,每个重复6 只羊,每组分别饲喂3个不同配比的日粮,即对照组(基础日粮,CON),酵母培养物A 组 (基础日粮+1%酵母培养物A,YCA);酵母培养物B 组(基础日粮+1%酵母培养物A+1.0×109CFU/g 活性酵母,YCB)。 预饲期10 d,正饲期60 d,正饲期后每组随机选取6 只羊进行指标检测。日粮配制根据肉羊营养需要量(NYT816-2021)配制,日粮组成及营养水平见表1。
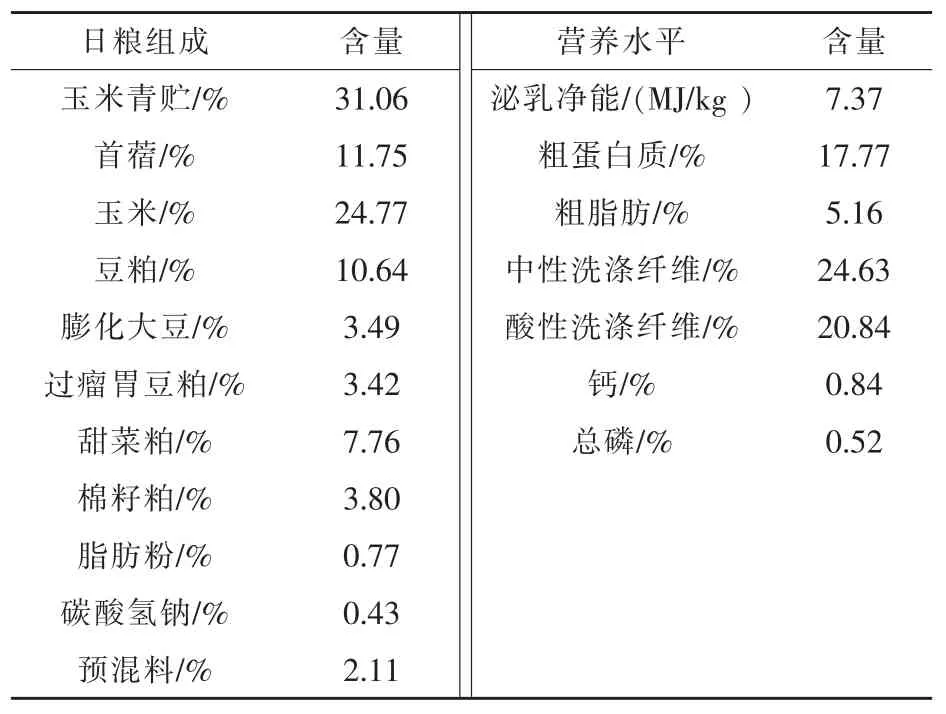
表1 日粮组成及营养水平(干物质基础)
酵母培养物A 与活性酵母均购自湖北安琪酵母股份有限公司, 酵母培养物A: 粗蛋白质≥15%,甘露聚糖≥2%,粗灰分≤10%,水分≤10%;活性干酵母:活菌含量≥2×1010CFU/g。
1.2 试验样品采集与处理 晨饲后2 h, 通过湖羊口腔采集瘤胃液,4 层纱布过滤并分装至5 mL离心管中,置于-80 ℃冻存备用。 通过比色法测定瘤胃液氨态氮含量(冯宗慈等,2013),采用气相色谱法(Yue 等,2009)测定总挥发性脂肪酸和各挥发性脂肪酸含量。
1.3 测定指标与方法
1.3.1 血清生化指标和生殖激素 试验结束后,湖羊颈静脉采血,3500 r/min 离心10 min 取上清, 采用全自动血清生化分析仪测定血清中总蛋白(TP)、白蛋白(ALB)、谷丙转氨酶(ALT)、谷草转氨酶(AST)、尿素氮(BUN)、甘油三酯(TG)、总胆固醇(T-CHO)含量。
1.3.2 瘤胃细菌测序 提取样品总DNA 后,根据保守区设计得到引物,在引物末端加上测序接头,进行PCR 扩增并对其产物进行纯化、定量和均一化形成测序文库,建好的文库先进行文库质检,质检合格的文库用Illumina NovaSeq 6000 进行测序。 高通量测序(如Illumina NovaSeq 测序平台)得到原始图像数据文件,经碱基识别(Base Calling)分析转化为原始测序序列(SequencedReads),其中包含测序序列(Reads)的序列信息以及其对应的测序质量信息。 首先使用Trimmomatic v0.33软件,对测序得到的Raw Reads 进行过滤;然后使用cutadapt 1.9.1 软件进行引物序列的识别与去除,得到不包含引物序列的Clean Reads,其次使用QIIME2 中的dada2 方法进行去噪, 双端序列拼接并去除嵌合体序列,97%聚类后获得操作分类单元(OTUs)。
1.4 数据统计分析 用Excel 2010 对数据进行初步整理,采用Minitab 21.3.1.0 统计软件中的单因素方差分析对所测指标进行统计分析。 利用百迈客云平台和origin 对瘤胃微生物多样性、 物种组成及丰富度、 物种差异以及主成分进行分析及作图。用P ≤0.05 表示差异显著,P ≤0.01 表示差异极显著。
2 结果与分析
2.1 酵母培养物和活性酵母对湖羊瘤胃挥发酸指标的影响 由表2 可见, 与对照组相比,YCB组瘤胃戊酸比例极显著升高(P <0.01),氨态氮浓度显著升高(P <0.05),其余瘤胃挥发酸含量各组间差异不显著(P >0.05)。
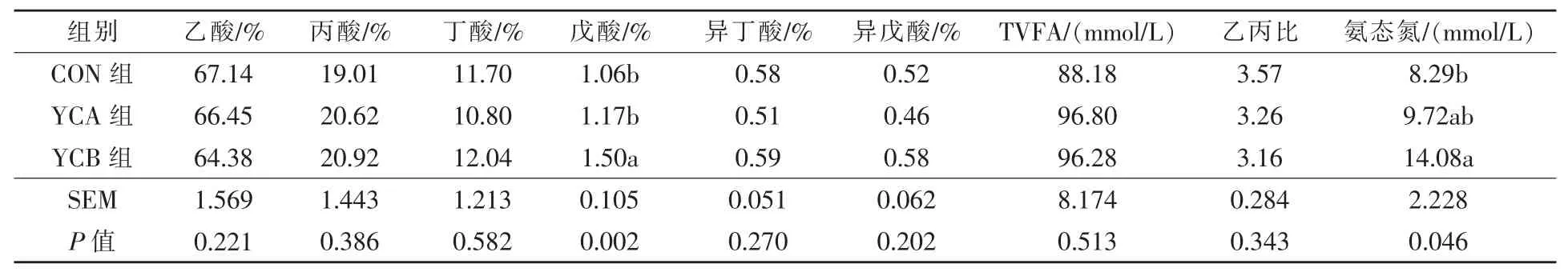
表2 酵母培养物和活性酵母对湖羊瘤胃挥发酸指标的影响
2.2 酵母培养物对湖羊血清生化指标的影响由表3 可见, 各组间血清生化指标差异均不显著(P >0.05)。

表3 酵母培养物和活性酵母对湖羊血清生化指标的影响
2.3 酵母培养物对湖羊瘤胃微生物区系的影响
2.3.1 测序结果及合理性 基于OTU 的Venn 图可统计多组或多个样本中所共有和独有的物种数目。 如图1 所示,以97%的一致性原则将序列聚类成OTU,共得到1996 个OTU,其中对照组OTU数为1471 个,YCA 组1350 个,YCB 组1467 个,共享823 个OTU,占总OTU 数的41.23%。
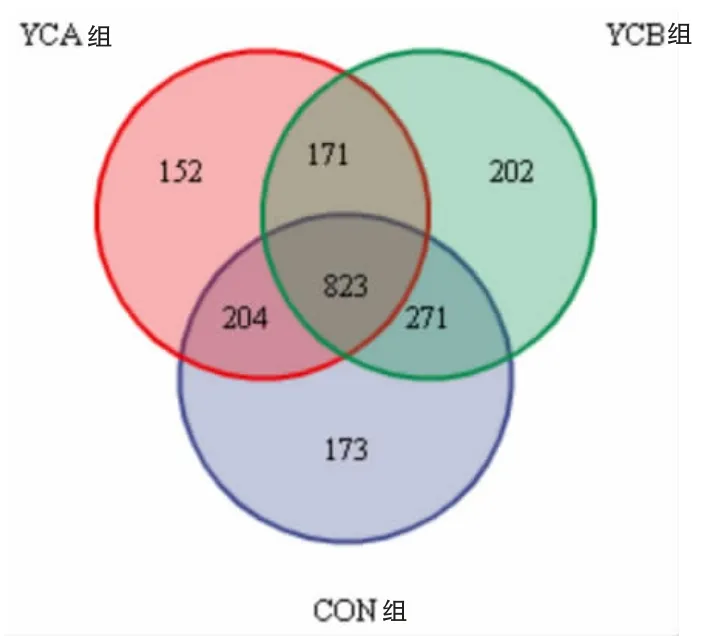
图1 Veen 图(基于OTU)
2.3.2 稀释曲线 瘤胃细菌稀释曲线如图2,稀释曲线呈现大幅度上升后趋于平缓, 说明深度覆盖到样本中的所有物种, 足以代表样本中的物种丰富度。
2.3.3 α-多样性 coverage 表示各样品文库的覆盖率,各组样品覆盖率均为99%以上。 如表4 所示,Chao、Ace、Shannon 和Simpson 指数均无显著的组间差异(P >0.05)。
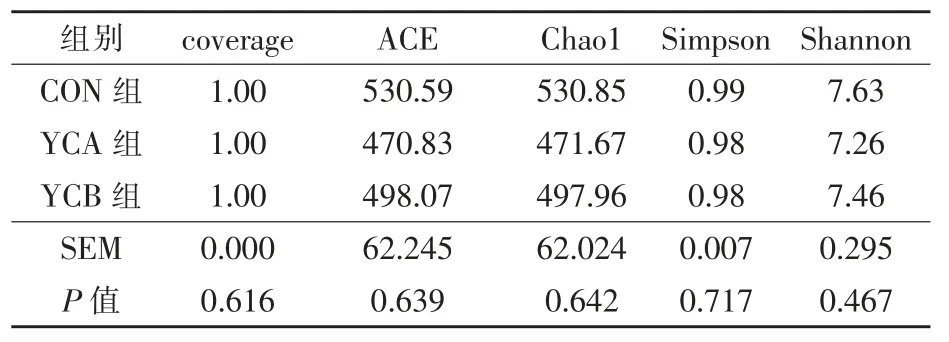
表4 酵母培养物对湖羊α-多样性指数的影响(OTU 水平)
2.3.4 β-多样性 分别在OTU 和属水平进行PCA 分析。 如图3 所示,在OTU 水平,PC1 和PC2对样品差异的贡献率分别为20.00%和16.92%。在属水平,PC1 和PC2 对样本差异的贡献率分别为41.60%和19.25%。 对照组、YCA 组和YCB 组瘤胃细菌在OTU 和属水平均聚类于不同的坐标位置,具有显著的组间差异。

图3 PCA 分析(OTU 和属水平
2.3.5 物种组成及丰富度
2.3.5.1 门水平 分析三组瘤胃细菌菌群在门水平的区别,共检测到相对丰度大于0.1%的细菌菌门11 个。由表5 可知,在门水平上,湖羊瘤胃优势细菌菌门(丰度>1%)主要分布在拟杆菌门、厚壁菌门和变形菌门。
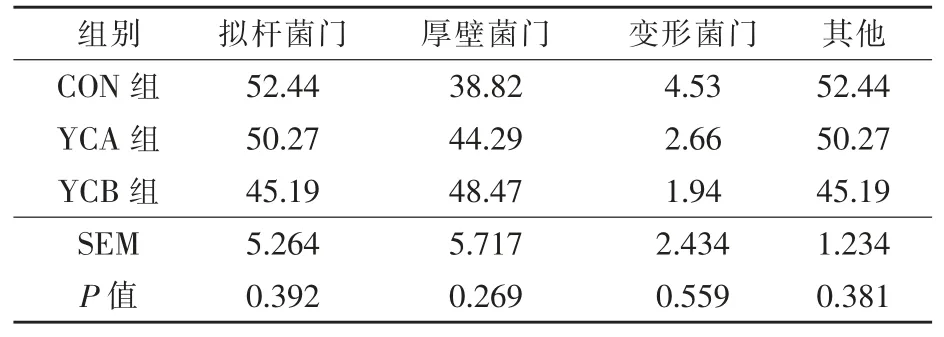
表5 酵母培养物对湖羊瘤胃细菌组成的影响(门水平>1%) %
2.3.5.2 属水平 分析三组瘤胃细菌菌群在属水平的区别, 共检测到相对丰度大于0.1%的细菌菌属66 个。 由表6 可知,在属水平上,共有优势菌属为普雷沃氏菌属,其余占比前10 的菌属分别为未分类的普雷沃氏菌科、 未分类的月形单细胞菌科、瘤胃球菌属、理研菌科RC9 群、未分类瘤胃细菌、解琥珀酸菌属、未分类的拟杆菌科RF16 群和普雷沃氏菌科UGG_003 群。 与对照组相比,YCA 和YCB 组的解琥珀酸菌属相对丰度显著降低(P <0.05)。
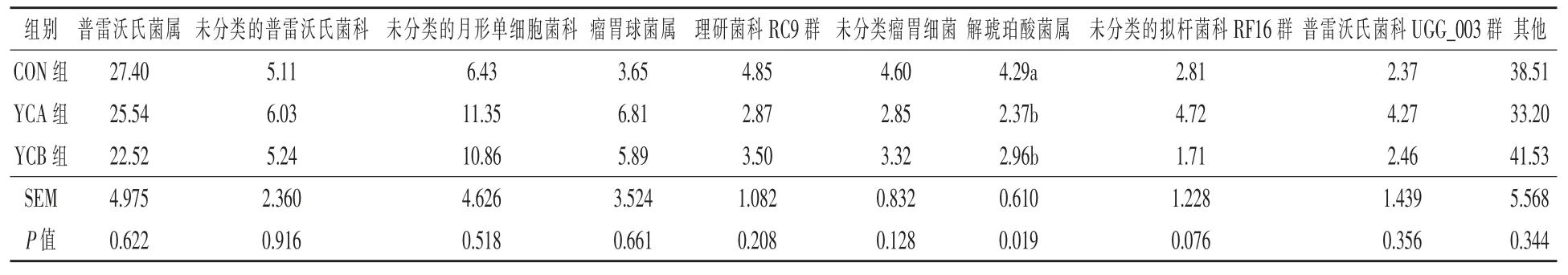
表6 酵母培养物对湖羊瘤胃细菌组成的影响(属水平TOP10) %
2.3.6 瘤胃细菌组成与发酵参数及血清指标相关性分析 图4 展示了湖羊瘤胃细菌属水平相对丰度(排名前10 的菌属)与瘤胃发酵参数及血清生化指标的相关关系。 其中普雷沃氏菌属相对丰度与血清总蛋白 (R=-0.502,P ≤0.05)、 总胆固醇(R=-0.568,P ≤0.05)、戊酸(R=-0.519,P ≤0.05)浓度呈显著负相关, 未分类的月形单细胞菌科与血清总蛋白(R=0.506,P ≤0.05)浓度呈显著正相关,与血清谷丙转氨酶(R=0.604,P ≤0.01)浓度呈极显著正相关, 瘤胃球菌属与血清谷丙转氨酶(R=0.540,P ≤0.05)呈显著正相关,未分类的拟杆菌科RF16 群与血清总蛋白 (R=-0.479,P ≤0.05)呈显著负相关。
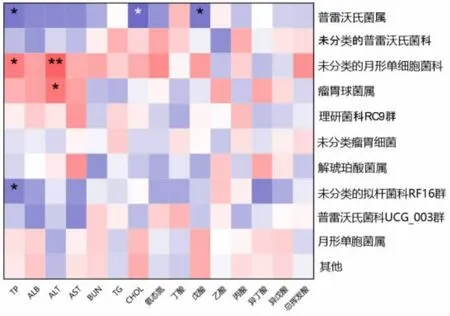
图4 相关性分析
3 讨论
3.1 饲喂酵母培养物和活性酵母对瘤胃发酵参数的影响 瘤胃发酵参数是影响瘤胃消化的重要因素, 其代表了瘤胃的微生物生长状况及瘤胃消化能力。 研究表明,酵母源物质可以影响瘤胃发酵参数, 提高丙酸的表达量, 降低乙丙比(张雷雷等,2023; 任胜男等,2020;Chaucheyras等;1996;Mutsvangwa 等,1992),这是由于酵母菌和乳酸菌等可竞争性利用发酵液中的糖类,刺激埃氏巨型球菌的生长, 后者可将乳酸降解为丙酸和丁酸(Amin 等,2021)。 但Desnoyers 等(2009)研究发现,活性酵母可以提高瘤胃挥发性脂肪酸浓度(平均+2.17 mM)并降低瘤胃乳酸浓度(平均-0.9 mM),对乙丙比没有影响。 在本研究中,1%酵母培养物与1.0×109CFU/g 活性酵母不会显著影响丙酸比例, 但添加后丙酸比例有升高趋势,乙丙比有降低趋势。 只添加酵母培养物不会显著影响戊酸比例, 同时添加酵母培养物和活性酵母,戊酸比例出现了显著增加,这与Opsi(2012)等研究一致。 Liu 等(2022)研究发现,在育肥牛日粮中添加4 g/头·d 活性酵母,可以显著提高瘤胃中丙酸比例, 但对乙酸比例及乙丙比无显著影响。 曾钰等(2020)也发现,在舍饲牦牛日粮中添加1%和1.5%的酵母培养物,可以显著提高瘤胃丙酸比例,降低乙丙比。 与活性酵母相比,酵母培养物中的活菌含量低,并不依赖活的酵母细胞起作用(Poppy 等,2012),酵母培养物中的有机酸、酶、小肽等均会对瘤胃发酵参数产生影响(姜奥宇等,2022;王小虎,2017;Vyas 等,2015; 马宁等,2009;Castillo 等,2004),而活性酵母更依赖于酵母菌的呼吸作用(Newbold 等,1996)。说明酵母培养物和活性酵母均可在一定程度上提高瘤胃丙酸比例, 降低乙酸比例,但功能性物质不同,可能需要更多的酵母培养物有效成分或更多的有效酵母活菌数才能达到乙酸和丙酸比例的显著改变。
瘤胃内氨态氮浓度通常与饲料蛋白的降解及微生物蛋白的合成有关, 还与瘤胃的尿素循环有关,6 ~30 g/dL 的氨态氮浓度不会产生不良的影响(Firkins 等,2007)。 在本研究中,瘤胃氨态氮浓度在此范围内, 添加1%的酵母培养物和1.0×109CFU/g 的活性酵母会显著提高瘤胃氨态氮浓度,只添加1%的酵母培养物氨态氮浓度有升高趋势, 说明酵母培养物会在一定程度促进瘤胃氨态氮生成, 活性酵母会进一步增加瘤胃氨态氮的生成能力,这与蔡大亮等(2022)的研究结果一致。此外,当瘤胃中氨态氮浓度偏低时,会有大量在循环尿素进入瘤胃,进而降低血清BUN 浓度,但在本研究中,YCA 组与YCB 组的血清BUN 浓度均有升高趋势, 说明酵母培养物和活性酵母可以改变瘤胃内环境, 促进微生物蛋白的合成与蛋白饲料的利用。
3.2 饲喂酵母培养物和活性酵母对血清生化指标的影响 血清生化指标不仅反映了营养物质的体内代谢情况, 还是衡量机体健康程度的重要标准。 血清TP 和ALB 可以反映日粮中蛋白质的消化吸收情况,而BUN 是蛋白质代谢的终产物,TP和BUN 的不足均会导致机体抵抗力降低,过多则会导致氮的浪费 (张学峰等,2016), 绵羊的TP、ALB 和BUN 参考范围分别为53 ~89 g/L、21 ~46 g/L 和2.86 ~ 14.28 mmol/L (Uyanik,2001;Pernthaner 等,1993;Kaneko,1980)。 在本研究中,TP、ALB 和BUN 均在参考范围内,且各组间无显著差异,这与Hassan 等(2016)研究一致。 血清TG、T-CHO 与脂质代谢密切相关,T-CHO 是指血液中所有脂蛋白所含有的胆固醇含量, 包括游离胆固醇和胆固醇酯,AST 与ALT 代表了肝脏的代谢情况。 本研究表明, 添加1%的酵母培养物和1.0×109CFU/g 的活性酵母不会影响脂质代谢及肝脏代谢。
3.3 饲喂酵母培养物和活性酵母对湖羊瘤胃细菌菌群的影响 本研究结果显示,CON 组的OTU数最多,其次为YCB 组和YCA 组,总体差别不大。 样本覆盖率高于99%,可以真实反映湖羊瘤胃细菌的结构和多样性。 α-多样性中Ace、Chao和Shannon 指数越大,多样性越高,Simpson 指数越小,多样性越高,在本研究中,各指数均无显著差异, 说明添加1%的酵母培养物与1.0×109CFU/g 的活性酵母不会影响瘤胃菌群多样性,这与Wang 等(2023)研究一致。 主成分分析结果表明,在OTU 水平,与对照组相比,YCA 组和YCB组的瘤胃细菌的菌群多样性有逐渐升高趋势,添加活性酵母后OTU 水平的瘤胃细菌菌群多样性更高;在属水平,第一主成分和第二主成分对样品的贡献值更高,CON 组与YCA 组的细菌菌群组成更接近,YCA 组与YCB 组的置信椭圆面积更大, 说明添加酵母培养物与活性酵母均可增加并改变属水平瘤胃细菌菌群多样性。 总而言之,添加1%的酵母培养物与1.0*10^9 的活性酵母均可增加并改变湖羊瘤胃细菌多样性,且在添加酵母培养物的基础上添加活性酵母,瘤胃细菌菌群的变异度更大。
瘤胃细菌的优势菌门通常为拟杆菌门和厚壁菌门(Auffret 等,2017;Klevenhusen 等,2017),在本研究中,瘤胃的优势菌门为拟杆菌门、厚壁菌门,这与前人研究结果一致。 Gao 等(2022)研究发现,在肉牛日粮中添加0.8 g/头·d 的活性酵母显著降低了瘤胃中变形菌门的相对丰度,但添加50 g/头·d 的酵母培养物对变形菌门相对丰度没有影响。 在本研究中,添加1%的酵母培养物和1.0×109CFU/g 的活性酵母不会对变形菌门相对丰度产生显著影响, 但与对照组相比有降低趋势, 这可能与酵母培养物和活性酵母的添加量较低有关。 郝凌魁等(2021)研究发现,在羯羊日粮中添加13.37%的酵母培养物后,拟杆菌门的相对丰度增加, 厚壁菌门的相对丰度降低,但与对照组无显著差异,这与本研究趋势正好相反,由于各组间拟杆菌门并无显著差异,本文不做进一步讨论。
普雷沃氏菌属是瘤胃中最多的拟杆菌门菌属,主要参与淀粉、木聚糖等能量物质的降解。本研究结果显示,在属水平上,各处理组的瘤胃优势菌群均为普雷沃氏菌属, 且各组间无显著差异,这与前人研究结果一致(孙美杰等,2022;曾钰等,2020)。 郭婷等(2022)研究发现,在围产前期奶牛日粮中添加120 g/d 酵母培养物,瘤胃细菌中解琥珀酸菌属及月形单胞菌属的相对丰度极显著降低。Jiang 等(2017)研究发现,在奶牛日粮中添加5.7×107CFU/头·d 的活性酵母可以提高纤维分解菌瘤胃球菌、 瘤胃杆菌和反刍兽新月单胞菌的浓度, 而添加6.8×108 CFU/头·d的活性酵母则可以提高瘤胃杆菌和反刍兽新月单胞菌的浓度, 可见不同的活性酵母添加剂量会对瘤胃属水平细菌菌群产生不同影响。 在本研究中,添加1%的酵母培养物和1.0×109CFU/g的活性酵母会显著降低接琥珀酸菌属的浓度,解琥珀酸菌属可将琥珀酸代谢生成丙酸盐,进而分解为丙酸,由此可以解释TVFA 中丙酸比例的上升原因。
瘤胃菌群通常与瘤胃挥发酸和血清生化指标密切相关, 微生物通过发酵产生挥发酸等小分子物质被瘤胃吸收, 并进入血液供机体生长代谢(郭延俭,2015)。 本研究发现,未分类的月形单胞菌科与血清AST 浓度呈极显著正相关,而瘤胃球菌属也与AST 浓度有显著正相关性。AST 主要分布在肝脏等组织中, 当组织细胞受损,AST 就会释放入血, 而肝脏还可以吸收氨合成尿素(冯仰廉,2004),进而参与瘤胃的尿素循环,因此,瘤胃球菌属和未分类的月形单胞菌科与AST 的相关性,可能与瘤胃的尿素循环有关。金红岩等(2020)在牦牛日粮中添加主成分为酵母菌的微生态制剂后, 也发现血清中AST 浓度有增加趋势, 且月形单胞菌属与瘤胃球菌属也有增加趋势,本研究与其一致。 普雷沃氏菌属的发酵产物主要包括乙酸、琥珀酸和丙酸,而栖瘤胃普雷沃氏菌是瘤胃中主要的蛋白降解菌之一(冯仰廉,2004),在本研究中,普雷沃氏菌属与血清TP 和CHOL 具有显著负相关性,与戊酸具有显著正相关,但与乙酸和丙酸无显著相关性,这可能由于普雷沃氏菌参与了蛋白的降解,但酵母培养物和酵母菌的添加量不足, 导致普雷沃氏菌属与乙酸和丙酸无相关性。
4 小结
本试验结果表明,1%的酵母培养物可在一定程度上提高瘤胃细菌多样性,改变瘤胃菌群组成,降低瘤胃解琥珀酸菌属比例。1%的酵母培养物和1.0×109CFU/g 的活性酵母可以增强酵母培养物的效果,进一步改善瘤胃发酵,提高瘤胃戊酸比例和氨态氮浓度。
