海参硫酸多糖与肠道菌群的相互作用及其调节糖脂代谢的研究进展
2023-10-17宋志远李双双赵前程
桑 雪,李 莹,童 瑶,于 双,宋志远,2,李双双,赵前程,*
(1.大连海洋大学食品科学与工程学院,辽宁 大连 116023;2.大连市特色海洋功效成分开发与高值化利用重点实验室,辽宁 大连 116023;3.辽宁省海洋健康食品工程研究中心,辽宁 大连 116000;4.大连工业大学,海洋食品精深加工关键技术省部共建协同创新中心,辽宁 大连 116034;5.大连民族大学生物技术与资源利用教育部重点实验室,辽宁 大连 116650;6.大连鑫玉龙海洋生物种业科技股份有限公司,辽宁 大连 116200)
随着市场上对海参食用和药用价值需求的日益增长[1],中国海参产业迅速发展,据2022年渔业年鉴报告,中国海参总产量为22.27万 t[2]。海参的保健功能一直备受关注,特别是近年来大规模疫情爆发之后更是得到广泛认可,其正逐渐从传统食材向健康机能性食品过渡。海参中含有多种功能性食品成分,海参多糖在含量上仅次于蛋白,是海参中最重要的功效活性物质之一。因此,对海参多糖的深入研究,可为海参营养和食疗价值提供科学依据,并为海参产业的健康发展提供理论依据。
硫酸多糖是多糖中单糖分子上某个或某些羟基被硫酸基团取代而形成的一类化学结构复杂、生物活性多样、构效关系鲜明的多糖类物质。海参硫酸多糖(sulfated polysaccharides from sea cucumber,SC-SPs)因其结构的高度重复性及其所蕴藏的巨大生物活性潜力,引发广大科研工作者的研究兴趣。近年来,越来越多的研究表明,SC-SPs能够调节机体能量代谢、缓解营养过剩导致的糖脂代谢疾病。
本文在综述SC-SPs结构的基础上,探讨了对肠道菌群变化与SC-SPs驱动健康益处之间的相关性,以及SC-SPs对肠道菌群和代谢产物生长的影响。
1 海参硫酸多糖的结构与性质
海参体壁是主要的食用和药用部位,而海参多糖是海参体壁的重要成分。国内外研究表明,海参体壁多糖的两种主要形式为海参岩藻聚糖硫酸酯(sea cucumber fucoidan,SC-FUC)和海参岩藻糖基硫酸软骨素(sea cucumber fucosyl chondroitin sulfate,SC-FCS),两者均为硫酸多糖并且结构特性(如分子质量、单糖组成、一级结构及链构象(高级结构))间存在显著差异。目前已经有大量的文献报道了这两类SC-SPs的结构与活性(图1和表1)。
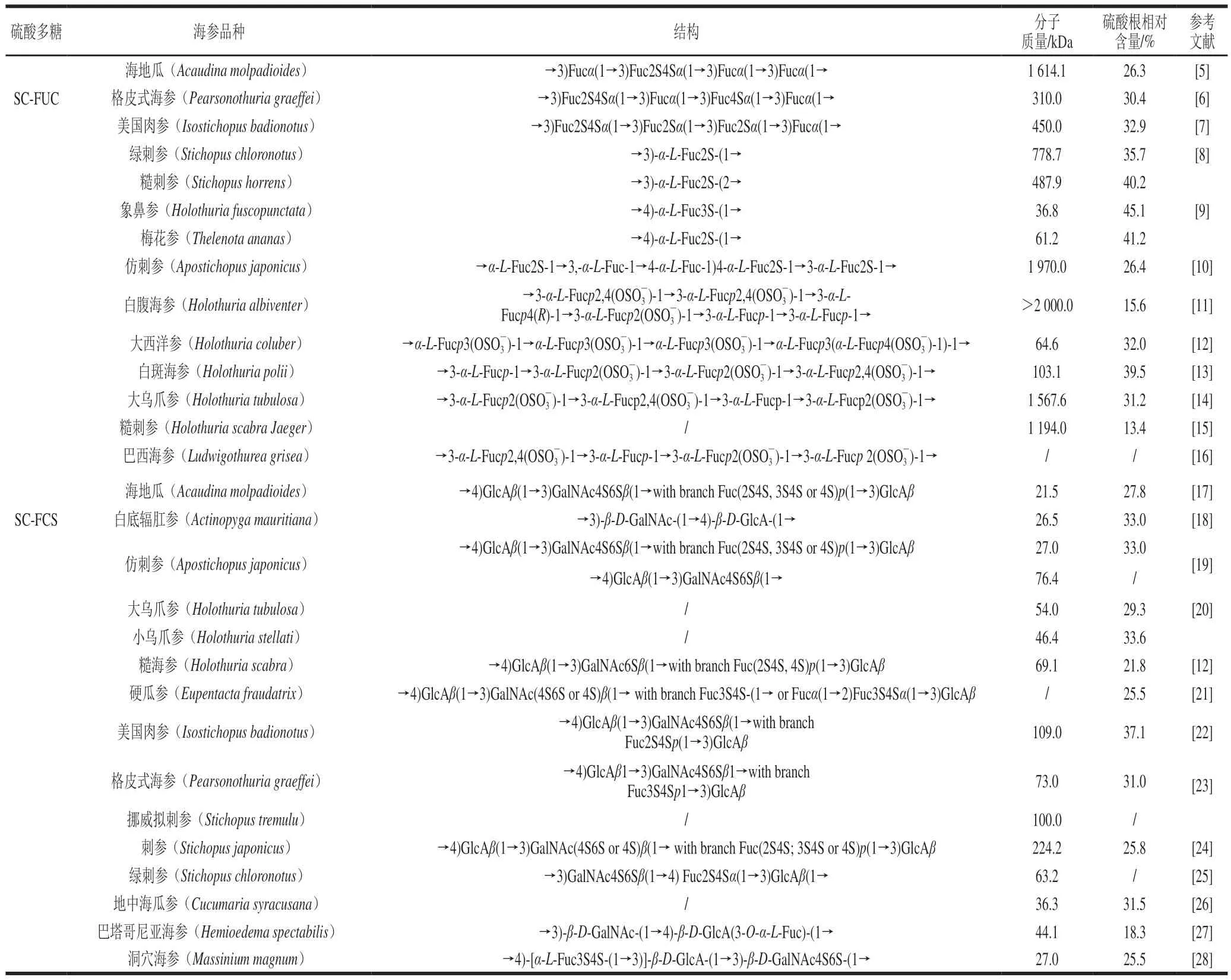
表1 海参硫酸多糖的来源及其结构、分子质量和硫酸根含量Table 1 Sources of sea cucumber sulfated polysaccharides and their structures, molecular masses and sulfate contents

图1 海参硫酸多糖的结构特性及主要生物活性[3-4]Fig.1 Structural characteristics and major biological activities of sulfated polysaccharides from sea cucumber[3-4]
FUC是一类以岩藻糖为主要单糖组成,部分羟基被硫酸根取代的酸性多糖,从藻类中提取的FUC不仅含有岩藻糖,也含有其他单糖如木糖、半乳糖、甘露糖、葡萄糖、鼠李糖等。与藻类FUC不同的是,从海洋动物中提取的FUC单糖组成较为单一,基本只含有岩藻糖[29]。从海参中提取的FUC通常具有重复的四糖结构序列,SC-FUC的结构不同之处取决于它们的硫酸根含量以及取代位点。如表1所示,从不同种类海参中提取的SC-FUC的一级结构均不同,它们具有不同的硫酸基岩藻糖重复单元、连接方式或硫酸化模式。
硫酸软骨素是一种存在于各种动物组织中的酸性黏多糖,是软骨素中硫酸基在不同位置的取代产物。硫酸软骨素是由糖醛酸(葡糖醛酸(glucuronic acid,GlcA)/艾杜糖醛酸(iduronic acid,IdoA))、N-乙酰氨基半乳糖(N-acetyl-D-galactosamine,GalNAc)及不同含量硫酸基组成。SC-FCS主链由β-D-GalNAc和β-D-GlcA构成二糖重复单元;而在SC-FCS中β-D-GlcA上的3 位支链由具有不同聚合度的L-岩藻糖构成[30],大多数N-乙酰-β-D-氨基半乳糖上出现4位和6位的双硫酸基取代结构[29]。岩藻糖支链会以1-2、1-3、1-4等键连方式与主链连接。岩藻糖支链往往存在不同硫酸根取代位点(2-O、3-O、4-O),且硫酸根取代个数也各不相同[29]。随着研究的不断深入,学者们发现不同海参来源SC-SPs的化学组成差异较大。
SPs根据单糖组成类型、糖苷键类型、分子质量以及重要的硫酸基团位置和程度而变化,结构上的差异主要是由于SPs的来源不同。然而,即使来自同一物种的FCS结构也不同。例如,Guan Ruowei等[19]测定的仿刺参(Apostichopus japonicus)FCS分子质量为76.4 kDa,而Ustyuzhanina等[18]测定相同物种的FCS分子质量仅为26.98 kDa。此外,Zhu Zhenjun等[24]提出刺参(Stichopus japonicus)的FCS有硫酸软骨素(chondroitin sulfate,CS)-E和CS-A,而Guan Ruowei等[19]发现刺参的FCS仅有CS-E,因此,结构特征的差异不仅限于SPs来源,还包括季节和栖息地的差异[23]。SC-SPs糖链组成和长度、硫酸基的数目和位置不同都会导致其生物活性不同[29],例如,FCS需要至少16%的岩藻糖基化程度才能通过激活肝素辅因子II来增强对凝血酶的抑制[31];具有3,4-O-二硫岩藻糖支链的FCS的降血脂作用强于具有2,4-O-二硫岩藻糖支链的FCS[22]。许多多糖通过硫酸化修饰可以显著改变多糖的结构特征,使其生物活性显著提高的同时,又赋予多糖新的生物活性[32-34]。多糖的侧链和硫酸根取代位点还影响着SPs的高级结构,其化学结构对其生物效应至关重要,例如,具有2,4-O-二硫化物的FCS比具有4-O-单硫酸盐和2,3-O-二硫酸盐的FCS具有更强的抗凝活性[23];再如Shang Qingsen等[35]提出,FUC的糖苷键类型能显著影响对肠道微生物群的调节。由此可见,了解特定SPs的化学结构能够避免对健康影响研究产生的误导。因此,本综述提供的有关SC-SPs结构和不同类型的信息有助于解释SC-SPs的生物活性机制,进一步促进其应用。
2 海参硫酸多糖与肠道菌群
人体胃肠道,尤其是大肠,是微生物群落中最大、最复杂的群体。胃肠道菌群包含多种微生物,包括与宿主共生的细菌、真菌和原生动物,其中细菌占肠道菌群的最大部分。人体内近80%细菌无法培养,而且这些细菌的大部分功能未知[36]。与巨大的物种多样性相比,肠道细菌群落主要由Firmicutes、Bacteroides、Proteobacteria和Actinobacteria四大门类组成,Firmicutes和Bacteroides是最主要的门。肠道微生物与宿主始终处于动态平衡的状态,这种平衡和稳定对宿主的生理功能有着深远的影响,包括免疫防疫功能的调节,以及能量和维生素的产生和合成[37-38]。
大部分多糖不能被人体直接消化吸收,却可以通过人体消化道安全到达远端肠道后与肠道菌群相互作用[39-41]。多糖可作为肠道菌群的碳源促进有益菌生长,并通过其代谢产物调节肠道菌群、改变肠道微生态多样性而发挥其益生元作用。近年来,通过使用益生元控制肠道菌群可以减轻肠道轻度炎症、改善肠道屏障的完整性,从而改善代谢平衡。多糖可以在一定程度上改变和重塑肠道菌群的多样性和组成,与此同时,多糖也可被肠道微生物菌群降解,产生代谢产物。肠道菌群代谢物(如短链脂肪酸(short-chain fatty acids,SCFAs)在预防高脂饲料喂养小鼠的肥胖、血脂异常和炎症方面发挥了重要作用。以上研究结果意味着肠道菌群的调节、有益细菌和代谢物的存在可能是实现宿主健康益处的关键策略。考虑到不同的多糖类型对肠道菌群和代谢物的影响不同,而SC-SPs作为一种独特类型的硫酸多糖,可能会产生不同的作用。本节总结了SC-SPs对肠道菌群和代谢物的调节作用(表2)。
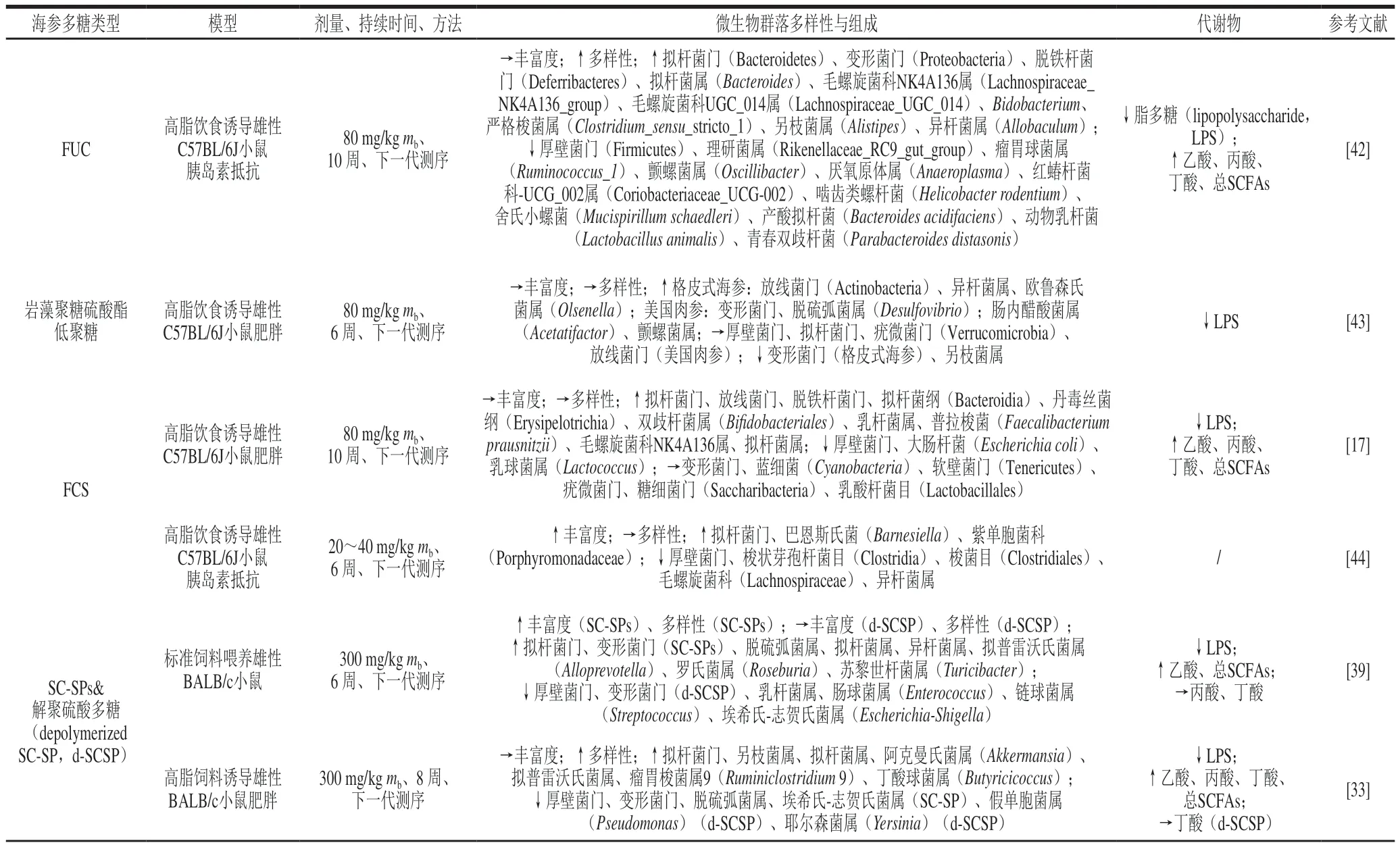
表2 海参硫酸多糖对肠道微生物群和代谢物的影响Table 2 Effects of sulfated polysaccharides from sea cucumber on gut microbiota and metabolites
2.1 海参硫酸多糖对肠道菌群及其相关代谢物的影响
肠道菌群的丰富性和多样性、微生物群落分类组成的相对丰度及其代谢物(如LPS、SCFAs和次生胆汁酸)的浓度被认为与宿主健康有关[48]。通过了解SPs对这些肠道菌群参数的影响,可以探索SPs对宿主健康产生有益作用的潜在机制。高细菌丰富度和多样性往往反映生态系统的稳定性和恢复力,并且与疾病及肠道菌群中某些物种数量减少存在一定的关联性[48]。对于SC-SPs来说,目前的研究表明,绝大多数可以提高肠道菌群的丰富度和多样性;也有一些研究表明SC-SPs对肠道菌群的丰富性或多样性的影响不大,这些差异可能归因于SC-SPs类型、分子质量、实验模型或剂量的不同(表2)。
Bacteroidetes、Firmicutes和Proteobacteria在肠道菌群中的相对丰度超过90%[49],而SPs的摄入可以调节这些菌门的相对丰度。大多数的SPs可以降低Firmicutes丰度、增加Bacteroides的丰度,表明由SPs调节的肠道菌群可能具有较低的Firmicutes/Bacteroides比率(F/B)[3]。Proteobacteria中包括许多已知的人类病原体,如沙门氏菌(Salmonella)、弧菌(Vibrio)和绿脓杆菌(Pseudomonas aeruginosa),SC-SPs对Proteobacteria的调控取决于SPs的类型、动物模型等,如两种FCS低聚糖对Proteobacteria的丰度影响不同[17,42]。用SC-SP处理的瘦小鼠的Proteobacteria丰度较高,在高脂饮食喂养的小鼠中补充SC-SP会降低其丰度,与此同时,在补充了SC-SP和d-SCSP的高脂饮食喂养的小鼠中Akkermansia丰度也有所增加[39];再如用美国肉参硫酸软骨素(fucosylated chondroitin sulfate fromIsostichopus badionotus,FCS-Ib)和美国肉参硫酸软骨素寡糖(fucosylated chondroitin sulfate’s depolymerized derivativefromIsostichopus badionotus,DFCS-Ib)均能够不同程度上恢复厚壁菌门和拟杆菌门构成比例,促进高脂膳食饲养小鼠肠道菌群的健康,其中,大分子多糖能够使F/B的比例恢复至正常水平,而DFCS-Ib作用弱于FCS-Ib[44]。有研究表明,摄入SPs可以促进有益细菌(Lactobacillus、Akkermansia、Bifidobacterium等)的增殖,从而使宿主更健康。除上述已知的有益细菌外,其他一些主要产生SCFAs的有益细菌(如Bacteroides和Alloprevotella)也可通过SC-SPs来富集。多形拟杆菌(Bacteroides thetaiotaomicron)是Bacteroides的代表物种,它包含多种糖苷水解酶和多糖裂解酶,能够参与SPs的降解[50]。在不同的实验模型中给予不同类型的SC-SPs可以提高模型体内Bacteroides的丰度[51];某些SC-SPs还可以通过抑制潜在有害细菌来调节肠道菌群,有研究表明SC-SPs可以显著抑制高脂膳食小鼠肠道中Escherichia-Shigella的丰度[33];而解聚SC-SPs可以显著抑制Pseudomonas和Yersinia的丰度[39]。
SC-SPs对潜在的LPS产生菌Desulfovibrio在不同情况下的调控存在显著差异,在瘦小鼠体内补充SC-SPs后,Desulfovibrio丰度显著增加;而高脂膳食小鼠体内的Desulfovibrio丰度却显著降低[33]。由于肠道微生态非常复杂,需要进一步的研究来阐明确切的机制,总地来说,如果SC-SPs在宿主健康方面能带来有益的效果,这可能是因为有益细菌的增加以及病原体的减少。肠道菌群与宿主相互作用的主要方式之一是通过微生物代谢中间产物或最终产物产生的小分子,SC-SPs对肠道微生物的代谢产物如LPS、SCFAs和胆汁酸(bile acids,BAs)的影响一直是研究的重点[52]。其中,LPS浓度被SC-SPs、SC-FCS、SC-FUC及SC-FUC寡糖等显著抑制;Chen Shiguo等[23]研究表明,FUC可以抑制肠道菌群中BAs含量,并将初级BAs转化为次级BAs;此外,SCFA被认为是肠道微生物群发酵多糖的主要终产物,对人体和动物等宿主产生多种有益影响,在不同的实验模型中,大多数SC-SPs对乙酸盐和总SCFAs具有显著的促进作用。基于SC-SPs结构的不同,其对肠道菌群的调节机制也不相同,Zhang Tiehua等[40]的研究也表明,天然多糖的糖苷键类型直接影响肠道菌群的分布。而由表1、2可知,SCSPs糖苷键型可能主要影响肠道菌群的多样性和代谢[35],确定肠道菌群如何利用特定多糖、肠道微生物间的互作关系及其代谢产物的特征具有重要意义。
2.2 肠道菌群参与硫酸多糖的分解代谢
人类基因组因没有编码足够的碳水化合物活性酶(CAZymes),从而限制了糖的酵解作用,而微生物基因集约为人类基因补体的150 倍[36],其中2.62%的基因被靶向分配至数十种不可消化多糖的CAZymes。因此,SC-SPs在人体内的代谢需要细菌源CAZymes的参与,Kaoutari等[53]报道了人类肠道菌群中具有代表性的CAZymes数据库和多样性,其中Bacteroidetes、Firmicutes每个基因平均分别可编码137.1 个和39.6 个CAZymes。由于SPs的水解只发生在细菌的细胞表面,这些细菌中的CAZymes必须包含输出到细胞表面的信号序列,Bacteroidetes中约81%的糖苷水解酶(glycoside hydrolases,GHs)和多糖裂解酶(polysaccharide lyases,PLs)具有信号序列,而Firmicutes中约有19%的GHs和PLs有信号序列。Bacteroides属隶属于Bacteroidetes门,研究发现其具有碳水化合物代谢途径,并能够编码不同种类负责降解SPs的降解酶,包括GHs、PLs和碳水化合物酯酶等[40],由上述推测,Bacteroidetes在菌群中主要负责SC-SPs的代谢。
硫酸多糖的降解模式还有以下两种:一是Firmicutes和Bifidobacterium仅降解少数SC-SPs的单个糖苷键;二是整个肠道菌群参与SC-SPs的代谢,发生大量代谢物互养作用[40]。目前,这一领域的研究较少且很大程度上仍处于理论研究阶段,对于肠道菌群参与SC-SPs的分解代谢是未来需要突破的一个重要研究目标。
3 肠道菌群影响宿主糖脂代谢
相对于遗传因素,膳食和药物等环境因素对肠道菌群的影响更为显著,肠道菌群的多样性和某些菌种所占的比例因宿主饮食中摄入的膳食纤维含量不同而存在明显差异[54]。宿主膳食中添加SPs后,某些特定物种微生物的丰度可增加30%以上,而肠道菌群的变化又将直接影响宿主糖脂代谢[55],因此,通过膳食干预调节宿主肠道菌群的平衡或可成为改善宿主代谢进而减少代谢性疾病发病的新途径。
多项干预性研究探索了SC-SPs对机体代谢改善的影响,结果显示SC-SPs可以降低研究对象的体质量、减少其体脂蓄积,并且上述效果在肥胖、糖尿病患者等已经患有代谢性疾病的研究对象中更为显著,表明SPs可在肥胖等代谢性疾病治疗方案中发挥重要的作用[3]。目前关于SPs改善机体代谢的研究中,大部分都集中在探讨SPs的摄入与肠道菌群及相关代谢产物的相互作用。摄入SPs等膳食纤维量较少时,肠道微生物发酵膳食纤维产生的SCFA含量也会相应降低,进而通过乙酸盐、丙酸盐和丁酸盐的作用使肠道内微生物新陈代谢由有利模式转化为产生更多有害代谢物的模式,而膳食纤维摄入较多者则能通过上述机制使机体糖脂代谢保持平衡,从而降低肥胖、2型糖尿病等代谢性疾病的发生率。
近年来,越来越多的研究表明,SC-SPs能够调节机体能量代谢,缓解营养过剩带来的代谢疾病[29]。目前,对于SC-SPs的活性研究集中在对糖脂代谢的调节方面。
3.1 肠道菌群与肥胖
肥胖被世界卫生组织定义为“异常或过量脂肪积累所造成的机体损伤”,已成为全球性的公共健康问题。身体质量指数超标已成为全球导致死亡的第5位风险因素,不仅如此,肥胖及其并发症已被证实与2019新冠病毒感染(COVID-19)的易感性和严重程度显著相关[56]。对于个体而言,肥胖通常是由于能量摄入与能量消耗不平衡所引起的体内能量过剩[57]。根据世界卫生组织的数据,2016年,超过6.5亿人被诊断患有肥胖症,预计到2030年,将有10亿人受到影响。全球23%心肌缺血和7%~41%的特定癌症都可归因于超重和肥胖[57]。肥胖及其多种并发症,如高脂血症、T2DM、某些癌症等,会导致患病者生活质量降低、死亡率增加,而且会带来巨额医疗费用负担。近年来,人们一直在探索有效、易获得以及低风险的替代疗法。
肥胖是由于长期的热量摄入超过能量消耗而导致的能量失衡,在现代环境中,这二者之间的不平衡都发生在肠道中[58]。肠道菌群在能量代谢中起着重要作用,大量研究表明,肠道菌群、宿主基因型和饮食之间的相互作用可能是导致肥胖的关键因素[59]。SPs的抗肥胖能力可能归因于通过抑制α-淀粉酶和脂肪酶等消化酶活力[24]、通过下调脂肪酸合成基因的表达来调节脂质代谢以及通过降低附睾脂肪组织基因表达来抑制脂肪的积累[60]。越来越多的研究表明,肠道菌群是参与体质量管理的一个新因素,在一对双胞胎中,肥胖个体与瘦弱个体相比肠道微生物多样性有明显降低,这种肠道菌群的差异与肥胖的发生密切相关[61]。在随后的研究中发现,减肥手术后患者体质量减轻伴随着细菌多样性的增加[62]。革兰氏阳性菌(G+)与革兰氏阴性菌(G-)的比例也与肥胖有关,肥胖个体的G+比例增加,而G-比例减少[63],已知肠道菌群中门水平的相对丰度变化与肥胖有关,特别是Firmicutes和Bacteroidetes[61]。在人类和动物肥胖模型中发现F/B比例增加,且宿主从食物中提取能量的有效能力增加[64]。目前,大多数SPs的抗肥胖作用与降低F/B比例有关。例如,从格皮式海参中提取的FUC[65]、从海地瓜中提取的FCS[17]以及从刺参中提取的SPs[33]均能降低肥胖小鼠的F/B,达到了减肥的作用。另外,在肠道菌群与肥胖的研究中,肠道菌群与肥胖在种水平上的相关性要显著高于门水平,虽然转基因与肥胖之间的具体关系尚不确定,但肥胖与肠道菌群之间确实存在着密不可分的联系[66]。Verrucomicrobia丰度与啮齿动物、人类的体质量减轻具有相关性,其中Akkermansia是隶属于Verrucomicrobia的一个属,在肥胖成年人中,高丰度的嗜黏蛋白阿克曼菌(Akkermansia muciniphila)与更健康的代谢状态以及限制热量摄入后更好的临床结果具有相关性[67]。这些研究表明,控制肠道菌群中的Akkermansia对于肥胖患者来说可能是一种潜在的治疗方法。
肠道菌群的变化通过各种潜在机制诱发肥胖的发生和发展(图2)。肠道菌群可以参与能量调节、发酵部分人体胃肠道中不能被消化酶消化的多糖,从而增加单糖和SCFA的含量[68]。SCFA通过降低禁食诱导脂肪因子(fasting-induced adipose factor,Fiaf)的表达和脂蛋白脂肪酶(lipoprteinlipase,LPL)的活性,导致脂肪细胞中甘油三酯聚集[63]。肠道微生物可通过抑制肝脏和其他外周组织中的腺苷——磷酸激活蛋白激酶(adenosine 5’-monophosphate-activated protein kinase,AMPK)增加脂肪积累,从而减少脂肪酸氧化[69]。此外,肠道菌群通过调节免疫反应和改变肠道通透性而导致肥胖;LPS可与肠上皮细胞的Toll样受体4(Toll-like receptor 4,T L R 4)结合,激活和上调炎症因子(白细胞介素(interleukin,IL)-1、IL-6、肿瘤坏死因子-α(tumor necrosis factor,TNF-α)等)。肠动力的增加导致LPS进入血液循环,血液中的内毒素增加导致全身炎症反应,从而促进脂肪生成并导致肥胖[70]。近期,Wang Qian等[71]首次发现IL-12细胞因子家族中的IL-27可以直接靶向促进脂肪细胞棕色化和产热,通过燃烧脂质以减轻肥胖和T2DM,为治疗肥胖及代谢性疾病提供了新的靶点。
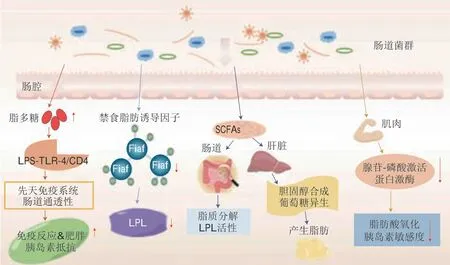
图2 肥胖的主要诱发机制[66,76]Fig.2 Pathogenesis of obesity[66,76]
多糖抑制饮食诱导肥胖的一个重要机制可能是肥胖受试者中产生SCFA的菌群数量增加,同时SCFA(主要包括醋酸盐、丙酸盐和丁酸盐)含量也增加。研究表明丁酸盐和丙酸盐可预防高脂膳食诱导的小鼠肥胖,并通过游离脂肪酸受体3(free fatty acid receptor 3,FFAR3)非依赖性机制调节肠道激素[72]。在饮食中补充醋酸盐、丙酸盐、丁酸盐或其混合物可以通过改变G蛋白偶联受体(G protein-coupled receptors,GPR)43和GPR41的表达来预防饮食诱导的肥胖;另有研究表明,SCFA可通过肠内分泌细胞中的GPR41发出信号,诱导肽YY的分泌,从而通过增加饱腹感预防肥胖[73]。SPs可通过增加相关的SCFA产生菌来抑制肥胖,其中通过口服分离自菲律宾刺参的FUC、分离自美国肉参的FCS以及刺参的SC-SP均可使SCFA产生菌的丰度增加,例如Alloprevotella、Bacteroides和Allobaculum等[44],这都与抑制小鼠的肥胖有关。现已知许多途径通过控制肠道菌群来调节肥胖,有研究表明,细菌多样性和丰富度低的个体要比细菌多样性和丰富度高的个体具有更显著的肥胖以及炎症特征[74]。同时有研究表明,益生菌的丰度与肥胖以及肥胖相关疾病呈高度负相关[75],例如Bifidobacterium和Lactobacillus。然而,目前关于SPs对肥胖受试者肠道菌群多样性、丰富度以及益生菌比例调节作用的研究较少。因此,有必要进一步研究通过肠道微生物群确定硫酸多糖,尤其是SC-SP的减肥作用机制。
3.2 肠道菌群与2型糖尿病
T2DM是一种复杂的代谢性疾病,在世界各地都普遍存在[77]。根据国际糖尿病联合会的数据,预计到2045年将达到6.93亿,其中T2DM至少占90%,此外,2017年全球约有500万人死于糖尿病[45]。因此,T2DM已成为21世纪人类健康的主要威胁之一。
T2DM患者的系统和器官长期处于高血糖环境中,并伴随着不同程度的并发症,其特点是糖耐量和胰岛素抵抗升高,且致残率及死亡率高[78]。T2DM被认为是遗传和环境相互作用的结果,其发生不仅与人类基因组的差异、饮食结构的改变和运动量的减少有关,近年来肠道菌群和T2DM发生发展之间的联系也逐渐被揭示。研究表明,动物和人类中肠道菌群平衡的变化可能是T2DM的致病因素之一。T2DM患者的特征是中度肠道菌群失调,有研究表明,T2DM患者的长双歧杆菌(Bifidobacterium longum)水平低于正常人[79]。一项对T2DM患者肠道菌群进行全基因组关联研究发现,一些丁酸产生菌的丰度有所降低,其中包括罗斯拜瑞氏菌(Roseburia intestinalis)和普拉梭菌(Faecalibacterium prausnitzii)[80]。由此可知,导致T2DM的原因不仅是单一微生物的改变,还与肠道菌群的多样性和平衡有关。
与非糖尿病大鼠相比,糖尿病大鼠肠道中Lactobacillus和Bifidobacterium显著减少[81]。Larsen等[82]研究结果表明,与对照组相比,糖尿病组中Firmicutes和Clostridium的相对丰度显著降低,Bacteroidetes/Firmicutes的比率与血糖水平呈正相关。近年来,海参多糖作为降血糖功效因子逐渐受到人们的关注。研究表明,美国肉参和海地瓜中的岩藻聚糖能够改善胰岛素抵抗、轻炎症反应、调节肝脏糖代谢,有效改善T2DM及其并发症[83];叶瓜参(Cusumaria frondosa)硫酸软骨素可提高胰岛素敏感性,降低血糖水平[84]。摄入HLP的糖尿病大鼠可以通过增加有益细菌的丰度和减少条件致病菌(如Anaerobiospirillum、Colinsella和Treponema)而有效优化肠道微生物群的组成,Lactobacillus和Bifidobacterium的减少与糖耐量受损密切相关,糖尿病大鼠摄入HLP后对这两种益生菌具有积极的调节作用[45]。SCFAs是由肠道菌群产生的关键信号分子,摄入海参多糖后,SCFAs产生菌Clostridium、Turicibacter、Allobaculum及Ruminococcus等丰度显著增加,此外研究表明,通过肠道发酵促进SCFAs的产生可以增强代谢功能,如调节葡萄糖平衡和抗炎活性[45]。因此,SC-SPs对宿主的肠道菌群有积极的调节作用,并能产生有益的代谢物,从而对T2DM有一定的缓解作用。然而,SC-SPs对肠道微生物群改善作用的更精确机制仍需要进一步确定。
肠道菌群失衡引起T2DM的机制如图3所示。SCFAs水平的异常是引起T2DM的重要因素,SCFAs可以直接作用于胰岛β细胞、促进肝脏糖代谢,并为肝脏代谢提供30%的能量;它还可以促进结肠上皮细胞的代谢,为结肠上皮细胞、肾脏和肌肉提供能量[85]。Bacteroides、Bifidobacterium、Streptococcus和Clostridium是常见的SCFA产生菌[85],这些肠道菌的缺乏会引起SCFAs的异常,而低水平的SCFAs会通过影响与巨噬细胞和T细胞相关的信号通路来促进炎症因子的释放,导致肠道炎症,从而引起胰岛细胞功能受损和胰岛素抵抗[86]。肠道菌群的失衡会导致G-的比例增加,LPS可以与CD14/TLR4结合,从而引起一系列的炎症反应[87]。肠道通透性的改变,促进LPS进入血液,引起低水平的慢性炎症,并使胰岛素分泌不足[63]。肠道菌群也会通过BAs来影响糖尿病的发展,其中肠道菌群可以作为BAs的调节器,影响BAs的产生和代谢。作为信号分子,BAs参与了能量代谢的调节,并抑制了肠道细菌的过度增殖;肠道菌群将初级BAs转化为次级BAs,促进G蛋白偶联受体5(G protein coupled receptor 5,TGR5)和法尼酯X受体(farnesoid X receptor,FXR)的激活[88],并在BAs重吸收和能量代谢中具有重要作用。肠道微生物群失调导致BAs的形成和其受体的激活受阻,并引发T2DM。

图3 肠道菌群失调导致的T2DM的机制[66,89]Fig.3 Mechanism by which intestinal microbiota disorder causes type 2 diabetes[66,89]
使用SC-SPs处理后,SCFAs浓度的升高可能会促进肠道上皮细胞的生长,导致肠道激素如胰高血糖素样肽-1(glucagon-like peptide-1,GLP-1)水平的增加。肠道中的这些变化可能导致胰腺B细胞淋巴瘤/白血病-2(B cell lymphoma/leukemia-2,Bcl-2)基因的表达量增加、促细胞凋亡基因Bax水平降低,从而使胰腺的损伤和功能得到改善。此外,激素水平的变化,包括瘦素、胰岛素、GLP-1和脂肪素,可能激活过氧化物酶体增殖物激活受体/磷脂酰肌醇3-激酶/蛋白激酶B信号通路,抑制下游信号CD36的表达,并促进人葡萄糖转运蛋白4(glucose transporter 4,GLUT4)的表达[45]。上述作用可以增强肝脏的胰岛素敏感性,使葡萄糖和脂质代谢活跃,因此SC-SPs具有缓解T2DM的潜力。
4 结 语
海参中重要的功效成分SC-SPs无毒性、结构独特,且因其广泛的生物活性而被大量学者研究,肠道菌群也成为研究热点。肠道菌群通过选择性地降解SPs而发挥有益作用,而SPs可作为肠道菌群的重要能量来源,调节肠道菌群的组成、维持人体的生理效应。与传统药物相比,SC-SPs安全且副作用小、对抗肥胖和T2DM有积极作用,可能的机制是通过其对糖代谢和脂质代谢的调节作用,而这种调节可能来自于SC-SPs在肠道中的消化产物及其对肠道菌群的影响。尽管现有的研究证实了SC-SPs对肠道菌群具有积极的调节作用,但需要进一步确定确切的分子机制。因此,未来应更加重视菌种之间的相互作用、细菌的代谢及其代谢产物和代谢物的功能对整个机体的影响。
