β-谷甾醇对脂多糖诱导的小鼠乳腺上皮细胞炎症反应及乳蛋白合成的影响
2023-10-16刘莉莉
刘莉莉 陈 敏
(黑龙江中医药大学药学院,哈尔滨 150040)
在乳业中,奶牛乳腺炎是一种病因复杂且高发的疾病,如果治疗不及时或不彻底,则很难恢复,并有复发和乳房脓肿的风险。乳腺炎发生后,乳汁的乳脂率和乳蛋白率等显著降低,降低了乳汁的产量和品质,造成的经济损失在奶牛的各种疾病中居于首位。乳腺炎主要由病原微生物引起,尤其是革兰氏阴性菌[1-2]。细菌感染后,乳腺内会产生大量的内毒素。内毒素的主要成分是脂多糖(LPS),它能使机体产生强烈的免疫原性反应。LPS可诱导乳腺上皮细胞或巨噬细胞的免疫应答,分泌大量炎症因子,因此LPS常被作为建立炎症动物模型的重要诱导物[3-4]。研究表明,LPS可与Toll样受体4(TLR4)结合,激活乳腺上皮细胞中的核因子-κB(NF-κB)等多种信号通路,进而促进细胞中白细胞介素-1β(IL-1β)、肿瘤坏死因子-α(TNF-α)等促炎细胞因子的释放以及环氧合酶-2(COX-2)和诱导型一氧化氮合酶(iNOS)的产生,明显扩大炎症反应,促进炎症加重[5]。奶牛乳腺炎发生时,乳腺组织的损伤会导致乳汁中乳蛋白等营养成分含量的降低,同样,奶牛乳腺上皮细胞炎症模型中乳蛋白相关因子的表达量也会显著下调[6]。哺乳动物雷帕霉素靶蛋白(mTOR)信号途径以及酪氨酸激酶2(JAK2)/细胞信号转导和转录激活因子5(STAT5)信号途径主要调控乳腺上皮细胞中乳蛋白的合成,而且2条信号途径之间可相互影响,共同调控泌乳相关因子等合成乳蛋白[7]。研究发现,LPS可能通过抑制乳蛋白合成通路mTOR和STAT5及其下游相关基因的表达,从而降低奶牛乳腺上皮细胞乳蛋白合成分泌功能[8]。
在临床应用中,乳腺炎的主要治疗方法是使用抗生素,但其不能有效控制炎症过程[9]。近期研究表明,联合使用抗生素和天然抗炎药物可有效缓解乳腺炎的进一步发展[10-11]。与抗生素相比,天然产物往往具有较强的抗炎作用,而且具有低毒、无残留、不易产生耐药性等优点,牛奶中也不会发生抗生素和耐药菌残留进入食物链而影响人体健康。因此,天然产物在乳腺炎治疗中的应用日益广泛,寻找新的有效的天然抗炎药物是目前奶牛乳腺炎治疗的热点。β-谷甾醇(BSS)是植物甾醇的一种,是很多药用植物的重要活性成分。临床试验证实β-谷甾醇具有降胆固醇、降血糖、抗氧化、抑菌和抗癌等多种生物活性。β-谷甾醇在小鼠胃组织炎症[12]、葡聚糖酸钠诱导的小鼠急性结肠炎[13]、慢性肥胖相关炎症[14]等炎症模型中发挥一定的抗炎作用。然而,β-谷甾醇对乳腺炎动物重要营养成分乳蛋白合成的影响尚未见报道。因此,本研究利用LPS构建了小鼠乳腺炎的体外模型,旨在探讨β-谷甾醇对LPS诱导的小鼠乳腺上皮细胞炎症反应及乳蛋白合成的影响,为β-谷甾醇在缓解乳腺炎的进一步应用提供理论依据。
1 材料与方法
1.1 试验材料
小鼠乳腺上皮细胞(HC11细胞)购自北京北纳创联生物技术研究院。
1.2 主要试剂
β-谷甾醇(纯度>98%)购自成都某生物科技有限公司。噻唑兰(MTT)法细胞活力检测试剂盒购自上海碧云天生物技术有限公司;RPMI-1640干粉、胎牛血清(FBS)购自美国Gibco公司;Trizol购自美国Invitrogen公司;Prime ScriptTMRT reagent Kit试剂盒、SYBR Premix Ex TaqTM试剂盒购自日本TaKaRa公司。STAT5、磷酸化信号转导和转录激活因子5(p-STAT5)、JAK2、磷酸化酪氨酸激酶2(p-JAK2)、mTOR、磷酸化哺乳动物雷帕霉素靶蛋白(p-mTOR)和β-肌动蛋白(β-actin)一抗购自美国Cell Signaling Technology公司。
1.3 试验设计和方法
1.3.1 HC11细胞的培养
采用生长培养基于37 ℃、5% CO2条件下培养HC11细胞,每隔24 h更换新鲜培养基。
1.3.2 β-谷甾醇及LPS对HC11细胞活力影响的检测
试验分为对照组(CON组,生长培养基处理)、乳腺炎症模型组(LPS组,1 μg/mL LPS处理)以及炎症模型药物组(BSS+LPS组,分别用5、10、20 μmol/L的β-谷甾醇预处理1 h后加入1 μg/mL LPS),每组设置5个重复。将处于对数生长期的HC11细胞接种于96孔板(1×104个/孔)中培养过夜,更换新鲜培养基并添加药物处理24 h后,各孔加入10 μL MTT(5 mg/mL),继续培养4 h后吸出培养液,各孔加入100 μL二甲基亚砜(DMSO)以溶解细胞中的甲瓒结晶,振荡10 min后用酶标仪检测各孔在490 nm的吸光度(OD)值并进行计算,试验重复3次。
1.3.3 β-谷甾醇对LPS诱导的HC11细胞炎性细胞因子、乳蛋白及乳蛋白合成信号通路相关基因mRNA表达的检测
试验分组及药物处理方法同1.3.2。将处于对数生长期的HC11细胞接种于6孔板中,待细胞融合度达到70%~80%时,添加药物作用细胞24 h后收集细胞,Trizol法提取RNA,根据Prime ScriptTMRT reagent Kit试剂盒说明书进行反转录,利用实时荧光定量PCR(qRT-PCR)试剂盒(SYBR Premix Ex TaqTM)检测HC11细胞TNF-α、COX-2、白细胞介素-6(IL-6)、IL-1β、iNOS、β酪蛋白(CSN2)、κ酪蛋白(CSN3)、乳清蛋白(LALBA)、STAT5、JAK2、mTOR、真核细胞翻译启动因子4E结合蛋白1(4EBP1)、核糖体蛋白S6激酶1(S6K1)的mRNA表达水平。试验所用基因的引物序列见表1,选用β-actin作为内参基因。qRT-PCR结果采用2-ΔΔCt相对定量的方法计算各目的基因的mRNA相对表达量。

表1 引物序列
1.3.4 β-谷甾醇对LPS诱导的HC11细胞乳蛋白合成信号通路相关蛋白表达的检测
试验分组及药物处理方法同1.3.2。将处于对数生长期的HC11细胞接种于6孔板中,待细胞融合度达到70%~80%时,添加药物作用细胞24 h后收集细胞提蛋白,Western blotting方法检测STAT5、p-STAT5、JAK2、p-JAK2、mTOR和p-mTOR的蛋白相对表达量。
1.4 数据处理
采用SPSS 22.0软件对试验数据进行单因素方差分析和Duncan氏法多重比较。试验数据均以平均值±标准差表示,P<0.05表示差异显著。
2 结果与分析
2.1 β-谷甾醇对LPS诱导的HC11细胞活力的影响
利用MTT法检测LPS单独处理以及5、10、20 μmol/L BSS+LPS共处理对HC11细胞活力的影响,结果如图1所示。与CON组相比,LPS组和5、10、20 μmol/L BSS+LPS组HC11细胞相对增殖率均无显著差异(P>0.05)。这说明LPS和β-谷甾醇对HC11细胞活力无影响,后续试验结果不受HC11细胞活力变化的影响。

数据柱标相同或无小写字母表示差异不显著(P>0.05),不同小写字母表示差异显著(P<0.05)。下图同。
2.2 β-谷甾醇对LPS诱导的HC11细胞炎性细胞因子mRNA表达的影响
如图2所示,与CON组相比,LPS组的HC11细胞TNF-α、IL-6、IL-1β、COX-2和iNOS的mRNA相对表达量均显著升高(P<0.05)。与LPS组相比,5、10、20 μmol/L BSS+LPS组的HC11细胞TNF-α、IL-6、IL-1β、COX-2和iNOS的mRNA相对表达量均显著下降(P<0.05);其中,5 μmol/L BSS+LPS组和10 μmol/L BSS+LPS组的HC11细胞炎性细胞因子的mRNA相对表达量较低。
2.3 β-谷甾醇对LPS诱导的HC11细胞乳蛋白mRNA表达的影响
如图3所示,与CON组相比,LPS组的HC11细胞CSN2、CSN3和LALBA的mRNA相对表达量均显著降低(P<0.05)。与LPS组相比,5、10、20 μmol/L BSS+LPS组的HC11细胞CSN2、CSN3和LALBA的mRNA相对表达量均显著升高(P<0.05);其中,5 μmol/L BSS+LPS组的HC11细胞CSN2、CSN3和LALBA的mRNA相对表达量较高。
2.4 β-谷甾醇对LPS诱导的HC11细胞乳蛋白合成通路相关基因mRNA和蛋白表达的影响
如图4和图5所示,与CON组相比,LPS组的HC11细胞STAT5、JAK2、mTOR、4EBP1和S6K1的mRNA相对表达量均显著降低(P<0.05),同时p-STAT5、p-JAK2和p-mTOR的蛋白相对表达量也显著降低(P<0.05)。与LPS组相比,5、10、20 μmol/L BSS+LPS组的HC11细胞STAT5、JAK2、mTOR、4EBP1和S6K1的mRNA相对表达量均显著升高(P<0.05),同时p-STAT5、p-JAK2和p-mTOR的蛋白相对表达量也显著升高(P<0.05);其中,5 μmol/L BSS+LPS组和10 μmol/L BSS+LPS组的HC11细胞STAT5、JAK2、mTOR和S6K1的mRNA和蛋白相对表达量较高。
3 讨 论
牛乳腺炎是影响世界乳业的重要疾病,会导致牛奶产量和质量的降低,从而给乳业造成巨大的经济损失。牛乳腺炎可由多种因素引起,最常见的因素是病原体感染,包括革兰氏阴性菌在内的多种微生物可引起乳腺炎,其中乳腺上皮细胞参与了抵御病原微生物入侵的第1道防线。
3.1 β-谷甾醇对LPS诱导的HC11细胞炎性细胞因子mRNA表达的影响
研究表明,LPS通过与乳腺上皮细胞膜的模式识别受体TLR4结合,激活多种炎症反应信号通路,导致各种炎性细胞因子TNF-α、IL-1β、IL-6、iNOS和COX-2的产生,这些炎症介质能够进一步放大炎症过程[15]。因此,常用LPS诱导乳腺上皮细胞炎症反应模型来研究乳腺炎潜在治疗药物的抗炎机制。炎性细胞因子能促进炎症的发生发展,TNF-α是感染早期的主要炎症因子,对中性粒细胞具有趋化作用。与TNF-α一样,IL-6和IL-1β在炎症过程中发挥重要作用。研究表明,一些植物活性成分可以通过抑制TNF-α、IL-6和IL-1β的产生来减弱LPS诱导的乳腺炎[16-17]。
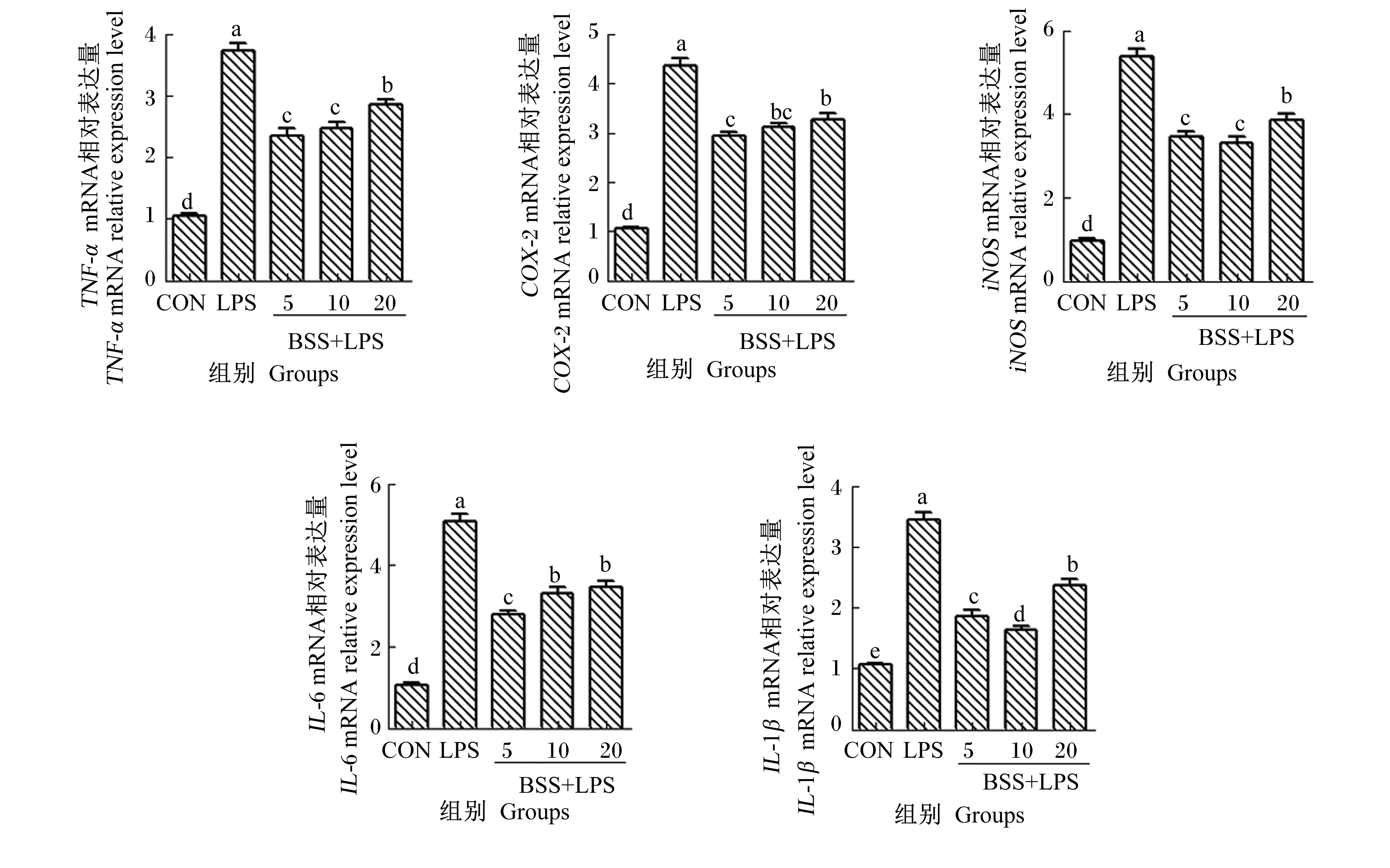
图2 β-谷甾醇对LPS诱导的HC11细胞炎性细胞因子mRNA表达的影响
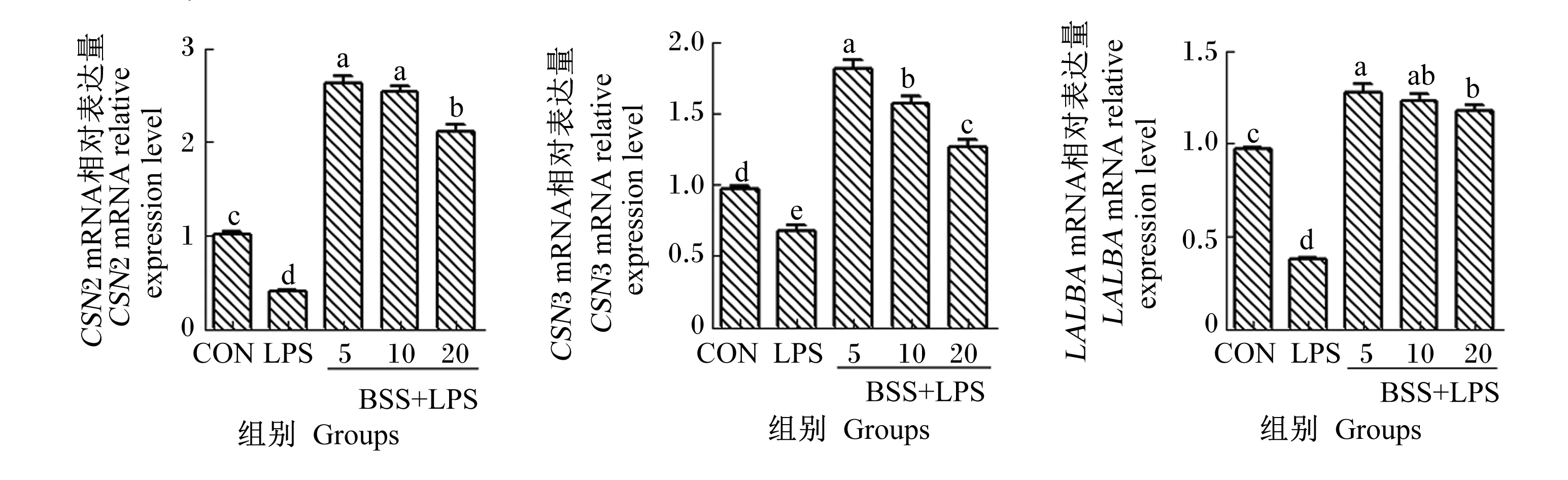
图3 β-谷甾醇对LPS诱导的HC11细胞乳蛋白mRNA表达的影响
本研究发现,与CON组相比,LPS组的HC11细胞TNF-α、IL-6和IL-1β的mRNA相对表达量显著升高;β-谷甾醇能显著抑制TNF-α、IL-6和IL-1β的mRNA表达,其中5和10 μmol/L β-谷甾醇抑制效果较好。Bi等[18]研究发现,β-谷甾醇可以抑制LPS刺激的人脐静脉内皮细胞中TNF-α、IL-6和IL-1β的mRNA表达。β-谷甾醇降低了过敏性哮喘大鼠炎症模型肺脏组织中TNF-α、IL-6等炎症因子的含量[19]。这些研究为本试验的结果提供了依据。COX-2是一种由细胞外和细胞内生理刺激(如LPS和TNF-α)激活的诱导酶;iNOS是一种一氧化氮合酶,被炎症、组织损伤等病理刺激激活,抑制iNOS的mRNA表达可以降低炎症介质一氧化氮(NO)含量。因此,COX-2和iNOS常被作为炎症标志物,它们的高表达与氧化应激和炎症的发生密切相关[20-21]。研究发现,iNOS和COX-2在LPS诱导的乳腺炎小鼠乳腺组织中也大量表达[22]。芹菜素和大豆活性肽等植物活性成分可以通过抑制LPS诱导的体内、体外乳腺炎模型中iNOS和COX-2的产生缓解乳腺炎[23-24]。Sun等[25]证明β-谷甾醇可抑制LPS诱导的小鼠小胶质细胞炎症介质IL-6、iNOS、TNF-α和COX-2的mRNA表达。本研究也发现,β-谷甾醇能抑制LPS诱导的小鼠HC11细胞iNOS和COX-2的mRNA表达,并且5和10 μmol/L β-谷甾醇的抑制效果较好。这些结果表明,β-谷甾醇可通过抑制促炎介质TNF-α、IL-6、IL-1β、iNOS和COX-2的mRNA表达来抑制乳腺炎的发展,而且研究提示低剂量的β-谷甾醇对小鼠乳腺炎症反应的抑制效果较好。
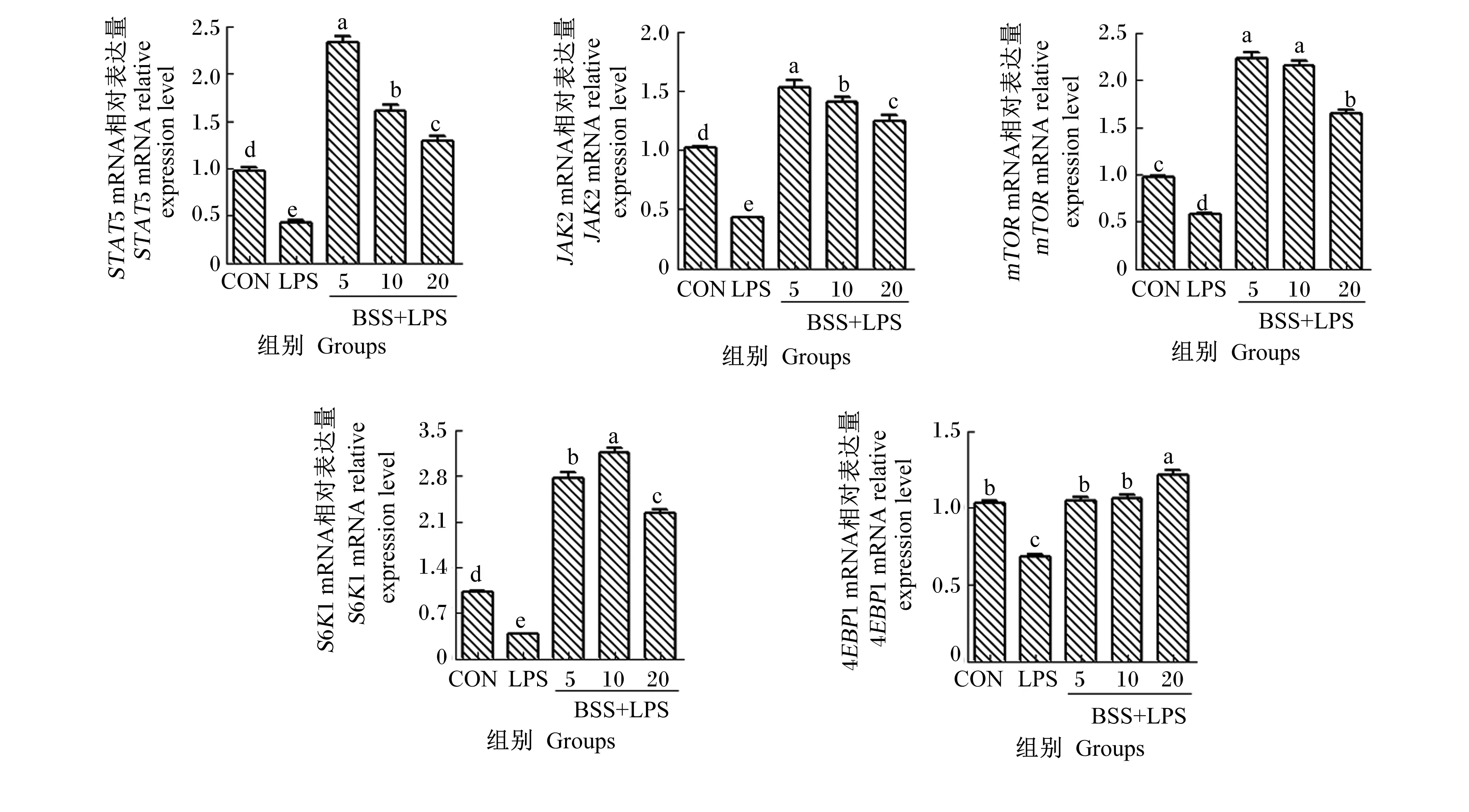
图4 β-谷甾醇对LPS诱导的HC11细胞乳蛋白合成信号通路相关基因mRNA表达的影响

图5 β-谷甾醇对LPS诱导的HC11细胞乳蛋白合成信号通路相关蛋白表达的影响
3.2 β-谷甾醇对LPS诱导的HC11细胞乳蛋白mRNA表达的影响
乳蛋白是牛乳的重要营养成分,主要成分为酪蛋白和LALBA。牛乳中酪蛋白有αs1酪蛋白(CSN1S1)、CSN2、αs2酪蛋白(CSN1S2)和CSN3等4种亚型。乳蛋白的合成受多种信号通路和激素在多个水平上的调控。LPS对乳蛋白合成有显著的抑制作用,Spitzer等[26]通过检测乳蛋白基因表达发现LPS显著降低了CSN2和LALBA的mRNA相对表达量。原利荣[27]发现LPS刺激能降低奶牛乳腺组织CSN2、CSN1S1和CSN3的mRNA相对表达量。李彦霞等[28]研究表明,大肠杆菌型乳房炎奶牛的乳腺组织中CSN1S1、CSN2和LALBA的mRNA相对表达量与健康奶牛相比均显著下调。植物提取物活性成分丰富,对乳腺炎奶牛乳蛋白率有明显的改善作用。Liu等[29]研究发现,薄荷醇可能通过抑制LPS诱导的奶牛乳腺上皮细胞炎症因子的表达,进而上调β酪蛋白的表达。辣木提取物能上调LPS刺激的奶牛乳腺上皮细胞CSN1S1、CSN1S2和CSN2的mRNA相对表达量[30]。为了探究β-谷甾醇在抑制小鼠乳腺上皮细胞炎症反应的同时是否可以改善乳蛋白的合成,本研究检测了β-谷甾醇对LPS诱导的HC11细胞乳蛋白mRNA表达的影响,结果表明LPS诱导的炎症反应可降低HC11细胞CSN2、CSN3和LALBA的mRNA相对表达量;而不同浓度的β-谷甾醇能不同程度地提高LPS诱导的HC11细胞CSN2、CSN3和LALBA的mRNA相对表达量,其中5 μmol/L BSS+LPS组CSN2、CSN3和LALBA的mRNA相对表达量最高。这些结果表明,β-谷甾醇在缓解LPS诱导的HC11细胞炎症反应的同时可提高乳蛋白的合成,而且低剂量的β-谷甾醇上调乳蛋白mRNA表达的效果最明显。
3.3 β-谷甾醇对LPS诱导的HC11细胞乳蛋白合成通路相关基因mRNA和蛋白表达的影响
哺乳动物mTOR通路和JAK2/STAT5通路是众所周知的调控乳腺中乳蛋白合成的关键信号通路。催乳素和促泌乳相关因子可通过JAK2使STAT5磷酸化,p-STAT5形成二聚体,进入细胞核与乳蛋白基因的启动子和增强子结合,诱导其表达,从而促进乳蛋白合成。激活的mTOR磷酸化其下游效应因子4EBP1,真核细胞翻译起始因子4E(eIF4E)与未磷酸化的4EBP1结合可抑制蛋白质的翻译;而4EBP1经mTOR磷酸化后会与eIF4E脱离,促进乳蛋白的合成。mTOR还能够促进S6K1磷酸化,增强含嘧啶基因mRNA的翻译,进而调节蛋白质的合成[31]。占今舜等[32]研究发现,LPS刺激的奶牛乳腺上皮细胞乳蛋白合成通路相关基因STAT5、mTOR和4EBP1的mRNA相对表达量显著降低。Zhang等[33]研究发现,LPS能降低奶牛乳腺上皮细胞中CSN1S1、CSN3、JAK2、STAT5、mTOR、4EBP1、S6K1的mRNA相对表达量。研究表明,植物提取物可能通过促进乳蛋白合成信号通路相关基因转录及磷酸化蛋白表达而提高LPS诱导的乳腺上皮细胞中乳蛋白合成水平。胡耀等[34]研究发现,黄花蒿乙醇提取物能上调LPS诱导损伤的奶牛乳腺上皮细胞中mTOR、S6K1、4EBP1、JAK2和STAT5的mRNA相对表达量。黄芪甲苷处理炎症乳腺上皮细胞时可促进CSN2基因表达,并可显著激活p-mTOR蛋白表达[35]。本研究结果显示,LPS诱导显著下调了HC11细胞乳蛋白合成信号通路相关基因的mRNA相对表达量,与前人研究结果一致。同时本研究发现,与LPS诱导损伤的炎症模型HC11细胞相比,β-谷甾醇能上调mTOR、S6K1、4EBP1、JAK2和STAT5的mRNA相对表达量及p-mTOR、p-STAT5、p-JAK2的蛋白相对表达量,其中5和10 μmol/L β-谷甾醇提高乳蛋白合成信号通路相关基因mRNA和蛋白表达的效果最好。这表明β-谷甾醇可能通过影响mTOR及JAK2/STAT5信号通路来促进炎症模型HC11细胞中乳蛋白的合成。
4 结 论
β-谷甾醇可抑制LPS诱导的小鼠乳腺上皮细胞炎性细胞因子的mRNA表达,同时可上调LPS诱导的小鼠乳腺上皮细胞乳蛋白合成信号通路(mTOR通路及JAK2/STAT5通路)相关基因的mRNA和蛋白表达,进而促进乳蛋白的mRNA表达。综合比较,以低剂量的β-谷甾醇缓解LPS诱导的HC11细胞炎症反应以及促进乳蛋白合成的效果较好。
