鳞翅目昆虫生物钟的研究进展与展望
2023-10-15全林发董易之池艳艳陈炳旭
全林发,姚 琼,董易之,徐 淑,池艳艳,陈炳旭
(广东省农业科学院植物保护研究所/农业农村部华南果蔬绿色防控重点实验室/广东省植物保护新技术重点实验室,广州 510640)
0 引言
地球自转和公转产生光、温可预测的周期性变化,生物长期适应昼夜光温变化及四季轮换而演化出内在自主计时机制——生物钟(circadian clock)。几乎所有的生物,包括细菌、真菌、植物和后生动物,都维持内源性的24 h计时制[1]。生物钟赋予昆虫预测时间和外界环境变化(如光照或温度)的能力,以协调代谢、生理、行为等生命过程与环境信号的同步,减少不必要的物质和能量消耗,极大地增强自身生存和竞争能力[2]。
在昆虫中,生物钟可调控其卵孵化、化蛹、成虫羽化、成虫运动、交配、取食行为节律及生理代谢活动等。1971 年,Konopka 等[3]在黑腹果蝇Drosophila melanogaster发现了第一个生物钟基因Period(Per),自此黑腹果蝇也成为了研究昆虫生物钟分子遗传机制的典范,同时推动了鳞翅目昆虫分子生物钟的研究。鳞翅目昆虫包括蝴蝶和飞蛾大约有16万种,是当前物种最丰富的昆虫群体,其中大部分具有夜行性(约占所鉴定种类的75%~85%),其余15%~25%属于日出型种类[4]。鳞翅目昆虫在生态系统中扮演着传粉者和猎物的重要角色,已知的主要农林害虫近70%属于鳞翅目,其幼虫部分以粮食作物为食,对经济和全球粮食安全有着重大影响。此外,一些体型较大的鳞翅目物种已成为研究昆虫发育、生理学、生态学和进化等生物现象的典范[5]。Williams[6]和Truman等[7]以鳞翅目昆虫为基础的开创性研究首次证实了昆虫昼夜节律主时钟存在于大脑,该成果极大推动了鳞翅目昆虫时间生物学学科的发展。
目前,从事鳞翅目昆虫生物钟研究的科学家以帝王蝶Danaus plexippus与鳞翅目模式昆虫家蚕Bombyx mori为主要对象,初步建立了鳞翅目昆虫生物钟分子调控模型,即生物钟基因的转录-翻译反馈环路(transcription-translation feedback loops,TTFLs)[2]。近些年来,鳞翅目昆虫生物钟的研究也渐渐拓展到其他非模式农林害虫生长发育与行为调控、生理解毒代谢与防治等领域,对深入了解鳞翅目昆虫丰富多变的生理行为调控机制,有效开展鳞翅目经济昆虫的生产及农林害虫防治具有重大意义。本研究总结了昼夜节律生物钟对鳞翅目昆虫孵化与取食、生长与变态、生殖与滞育、求偶与迁徙等生理行为的影响,重点概述了鳞翅目昆虫生物钟分子调控机制、生物钟与内分泌激素协同调控机制等重要研究进展,并基于生物钟原理探讨了鳞翅目昆虫与植物协同进化关系及重要生态意义,最后展望了将生物钟理论应用于农业害虫防治和经济昆虫饲养改良的应用前景。
1 鳞翅目昆虫生理行为生物钟
1.1 生物钟与鳞翅目昆虫发育和变态
鳞翅目昆虫属于完全变态昆虫,个体发育经过卵、幼虫、蛹和成虫4 个不同的虫态。生物钟系统在鳞翅目昆虫卵孵化、幼虫蜕皮生长、化蛹和羽化整个发育历程中起到不可或缺的调节作用。昆虫卵孵化是最早被描述的生物钟事件[2]。在棉铃虫Helicoverpa armigera、柞蚕Antheraea pernyi、玉米螟Ostrinia nubilalis等鳞翅目昆虫中,卵孵化节律受环境光或温度刺激诱发,这种节律属于内源性的生物钟节律,在转入持续黑暗后仍能维持。许多鳞翅目昆虫成虫羽化也具有明显昼夜节律性,如蚕蛾总科,夜蛾科,螟蛾科和凤蝶科等。而成虫的羽化节律不仅受光照影响,还具有一定温度补偿效应,同时会受饮食和营养元素的影响[8]。此外,鳞翅目昆虫滞育是研究昆虫感应光周期变化的典范。昆虫滞育发生与内源生物钟息息相关。目前鳞翅目昆虫滞育生物钟研究主要集中在蛹或卵滞育发生前的诱导阶段(diapause induction)。其中家蚕卵滞育研究最为深入,其卵滞育发生在受精核15次分裂的阶段,并受亲代胚胎发育后及胚后发育期生物钟授时因子(光照、温度等)的诱导[9]。对于蛹滞育的昆虫,蛹滞育诱导取决于暗期的长短,在暗期介入光照中断后会降低蛹滞育诱导的效果,在二化螟Chilo suppressalis、玉米螟、环带锦斑蛾Pseudopidorus fasci、黑纹粉蝶Pieris melete等研究中均得以证明[2]。除此之外,有研究认为鳞翅目昆虫其他时期发育同样受生物钟系统和光周期的影响[2]。
1.2 生物钟对鳞翅目昆虫生殖的影响
早期,鳞翅目昆虫生物钟研究着眼于生殖行为及生理方面,包括成虫交配行为、性信息素(pheromone)分泌与响应等。鳞翅目成虫交配由生物钟调控,交配前雌蛾求偶鸣叫,并通过尾部性腺节律性合成和释放性信息素,雄蛾则同步响应雌蛾性信息素,从而产生昼夜节律性的交配行为。例如:黄地老虎Agrotis segetum成虫交配行为受信息素生物合成激活肽(Pheromone biosynthesis activating neuropeptide,PBAN)调节,并由中枢生物钟系统控制[10]。灰翅夜蛾Spodoptera littoralis成虫成功交配的重要前提是两性生殖行为的节律同步,否则交配率会降低[11]。二点织螟Aphomia sabella雌蛾会周期性分泌6种性信息素类似物,且对雄蛾具有引诱活性[12]。此外,许多蛾类雄虫精子由睾丸释放到上层输精管的过程具有明显昼夜节律,如苹果小卷蛾Cydia pomonella和灰翅夜蛾等[7,13-14]。在灰翅夜蛾中,雄蛾睾丸中卵黄原蛋白释放与分子震荡器有关,其中Per基因沉默使得精子释放节律异常,进而影响卵受精[15]。除此之外,部分鳞翅目昆虫的精子活力也存在节律性变化。
1.3 生物钟与鳞翅目昆虫求偶及其他行为
鳞翅目昆虫求偶行为(courtship behavior)是生物钟行为学研究中非常丰富的部分。在许多夜蛾科、螟蛾科、天蛾科等鳞翅目昆虫中,均发现其节律性求偶行为[2]。近年来,鳞翅目幼虫周期性取食行为(feeding behavior)研究对于昆虫生产和害虫控制具有重要意义。鳞翅目幼虫主要以植物组织为食,随着龄期增加幼虫食量往往大增,最新研究表明夜蛾科的斜纹夜蛾和灰翅夜蛾末龄幼虫取食行为存在明显的昼夜节律性,而部分鳞翅目昆虫取食花蜜(或花粉)或访花行为也受生物钟系统的影响[14]。另外,帝王蝶Danaus plexippus通过自带的时间补偿-太阳罗盘精准定位并达到迁徙目的是鳞翅目昆虫经典的生物钟行为学案例[16]。除此之外,鳞翅目幼虫排泄、清理肠道等自由运动节律事件也有报道[17-18]。
2 鳞翅目昆虫生物钟分子调控机制
2.1 鳞翅目昆虫信号转导的核心生物钟基因
目前,鳞翅目昆虫生物钟基因的克隆和鉴定相关的报道不算多,最早在家蚕和帝王蝶中生物钟研究中开展,主要包括周期蛋白基因(Period,Per)、永恒蛋白基因(Timeless,Tim)和2 个隐花色素基因(Cryptochrome,Cry1和Cry2)、周期循环蛋白基因(Cycle,Cyc)和时钟蛋白基因(Clock,Clk)等核心生物钟基因。随后,人们又分别在鳞翅目天蛾科柞蚕[19];夜蛾科灰翅夜蛾[20]、粘虫Pseudaletia unipuncta[13]、斜纹夜蛾Spodoptera litura[21]、甜菜夜蛾Spodoptera exigua[22]、黄地老虎[10]、东方粘虫Mythimna separata[23],以及卷叶蛾科苹果蠹蛾Cydia pomonella[24]和梨小食心虫Grapholita molesta[25]中鉴定出上述核心生物钟基因。近期,双时蛋白基因(Doubletime,Dbt)在斜纹夜蛾[21]和棉铃虫[26]中被鉴定;旋转蛋白基因(Vrille,Vri)在斜纹夜蛾[17]中被鉴定;最新发现的核心钟基因(Clockwork orange,Cwo)也在棉铃虫[27]和斜纹夜蛾[18]中被鉴定。这些核心生物钟基因的克隆和鉴定结果为阐明鳞翅目昆虫生物钟分子机制研究奠定了基础。
2.2 鳞翅目昆虫的生物钟分子调控信号路径
昆虫周期性节律特征通常由中枢和外周组织、信号输入和节律输出等成分构成的调控系统来产生和维持。其中以温度和光照为代表的环境信号可借助输入途径传递到中枢生物钟系统,经过处理、整合后经输出系统将节律信号传递出去,或联结外周生物钟系统共同调节并维持机体的正常生理活动[21]。
对于昆虫目而言,生物钟研究最早起源于模式生物果蝇昼夜节律的探索,而后逐渐拓展到各种非模式昆虫。在果蝇中,生物钟调节系统是由一系列生物钟基因构成的至少3 个相互依赖的转录-翻译反馈环路(transcription-translation feedback loops,TTFLs)(图1A)。第1个TTFL中,PER/TIM蛋白在细胞质中形成二聚体,夜间转移至细胞核,抑制CLK/CYC的转录活性,从而抑制自身的转录。第2 个TTFL 中,PDP1ε是Clk基因转录的激活物,VRI 与PDP1ε竞争P/V-box 结合位点,抑制或去抑制Clk基因的转录。第3 个TTFL中,Cwo基因与CLK/CYC的靶标E-box元件特异性结合而抑制CLK/CYC 介导的转录活性,抑制自身转录和Pdp1ε、Vri、Tim和Per等基因的转录。上述TTFLs中的基因被转录、翻译为蛋白质,随后累积至触发转录关闭的阈值,此模型是昆虫生物钟研究的基础。与果蝇的TTFLs 有所不同,鳞翅目昆虫由于进化出2 种类型的CRY,一是与果蝇dCRY 同源光敏感CRY1,另一个是与脊椎动物类似的光不敏感CRY2,从而演化出不同于果蝇的新TTFL(图1B)。在帝王蝶季节性迁徙机制研究中发现,DpCRY1 能够介导DpN1 胚胎细胞系中DpTIM降解[28-29]。在DpN1胚胎细胞系和大脑中,DpCRY2 与DpPER 和DpTIM 相互作用,共同抑制DpCLK:DpBMAL1介导的转录活性[30-31]。由此衍生了以帝王蝶为代表的鳞翅目昆虫分子生物钟环路模型。在新的TTFL中,CRY一旦接受光照,便引起TIM磷酸化,使得PER/TIM/CRY2三聚体降解,然后CRY2:PER二聚体结合到CLK和BMAL1的E-BOX上,从而调节Cry2、Per和Tim等基因的表达,进而产生相应的节律变化,其中CRY2 主要起到转录抑制的作用[29,32](图1B)。这一环路模型在帝王蝶Cry2和Clk基因敲除等试验中得到了验证[33-35]。随后,人们基于果蝇核心生物钟元件同源序列比对与分析,对帝王蝶和秋尺蛾Operophtera brumata基因组进行组装与鉴定,逐步推测整合出鳞翅目昆虫分子生物钟的第2 和第3 个TTFL[32,36](图1B)。在第2个TTFL中,CLK:BMAL1复合体诱导Vri和Pdp1 基因转录,这2 个基因一旦被翻译,就会竞争结合VP-box 位点,从而抑制Clk基因转录[35]。第3 个TTFL 中,CLK:BMAL1 二聚体激活Cwo基因转录,CWO 进入细胞核,通过与时钟基因E-box元件结合,抑制CLK:BMAL1 的活性。其中光在CRY1 和JET 介导的TIM 降解过程中起重要作用,但在鳞翅目昆虫中引发CRY2降解的途径仍然未知。此外,由DBT,CK2 和SGG 介导的磷酸化可调节生物钟蛋白的活性、蛋白互作、核易位和降解等途径。上述鳞翅目昆虫3个TTFLs模式图的绘制为进一步研究鳞翅目昆虫生物钟调控机制奠定了理论基础。
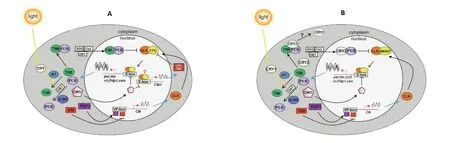
图1 昆虫生物钟基因的转录-翻译反馈模型[14]
2.3 鳞翅目昆虫发育与变态生物钟分子作用机制
目前,揭示鳞翅目昆虫发育过程中的“守时”奥秘方兴未艾,而探明该过程中分子作用机理对于理解昆虫发育与变态进程具有重要生物学意义。从胚胎发育中期开始,卵孵化节律便受内源生物钟调控。柞蚕脑移植实验最早证明,控制卵孵化节律的生物钟基因存在于大脑中[19]。近期研究发现,在正常光周期(LD)和持续黑暗(DD)条件下,家蚕胚胎中Per、Tim、Cry和Clk基因表达随时间节律性震荡,其中Tim和Per参与卵孵化昼夜节律的信号输出,且Per在此过程中起到负反馈调节作用[37-39]。然而,Per和Tim基因在鳞翅目昆虫生物钟行为中的负反馈调节作用仍存在一定争议。近期研究发现,家蚕Per基因敲除后试虫出现羽化规律紊乱的现象[38]。类似的,梨小食心虫Per和Tim基因表达干涉后对其成虫羽化节律有显著影响[25]。上述结果为进一步探明Per和Tim基因在鳞翅目昆虫昼夜节律调控中的作用机制提供了理论基础。
除昆虫大脑外,在鳞翅目昆虫头部、中肠、脂肪体和腺体等组织中的外周生物钟(peripheral clocks)基因也参与调控整个发育历程。人们首先在柞蚕脂肪体细胞质中检测到非循环的PER染色[40]。在幼虫发育过程中,部分幼虫组织同时表达多个钟基因。例如,在家蚕末龄幼虫的头部、中肠、脂肪体和丝腺中Tim、Per、Clk和Bmal1 生物钟基因转录活跃,且表现出不同日节律变化[14]。同样,不同钟基因在斜纹夜蛾末龄幼虫中,头部、中肠和脂肪体中也存在节律性表达特性。在全黑暗条件下,斜纹夜蛾幼虫头部Cwo、Bmal1 和Clk基因表达仍维持节律震荡,但Per和Tim基因日表达节律丧失[18]。由此推测,斜纹夜蛾幼虫没有一个完全成熟且独立于Per和Tim的具有自我持续振荡功能的昼夜节律生物钟。另有研究表明,其他独立于Per或Tim基因调控机制也能维持幼虫发育阶段的内源性计时系统,例如斜纹夜蛾幼虫从5 龄开始就表现出进食、排便和运动行为的昼夜节律[18]。类似的,灰翅夜蛾脂肪体中运行着一个独立于大脑中枢生物钟的外周生物钟系统[20]。然而,目前关于鳞翅目昆虫外周生物钟与中枢生物钟(central clocks)的协同互作机制仍待进一步挖掘明晰,以期进一步理解生物钟对昆虫生理行为节律的调控作用及昆虫-环境进化关系。
3 鳞翅目昆虫生物钟与内分泌激素协同调控机制
生理时钟和内分泌系统在生理和行为上的协同调控是一个令人着迷而又困惑的课题,近年来备受关注。研究表明,生物钟调节机体的代谢和内分泌系统,使机体适应环境温度、光照和食物[41-43]。这是由一系列转录因子通过转录-翻译反馈环(TTFLs)和内分泌激素的相互调节而介导的[44-46]。其中昆虫滞育是一个受生物钟和内分泌系统共同调控的经典而长期备受关注的课题。
在蚕业和昆虫学领域,温度和光诱导的滞育机制与滞育激素(Diapause hormone,DH)协同调控机制是一个持续了100多年的话题。在许多成虫滞育的昆虫中,昼夜节律系统的紊乱会影响滞育的发生,然而关于生物钟与内分泌激素调控的分子调控机制的研究仍缺乏深入报道。家蚕是鳞翅目昆虫唯一的模式昆虫,其滞育是研究光、温两大授时因子协同作用机制的理想状态。近期,Ikeda 等[47]利用TALEN 技术构建了一个家蚕生物钟基因Per敲除的突变体,并证明了敲除Per基因可破坏家蚕昼夜行为节律和时钟基因的时序表达模式,进而影响光周期对家蚕胚胎滞育诱导的调控作用。Cui 等[48]研究表明,家蚕生物钟基因Per敲除后,试虫生物钟系统TTFLs受损,在蛹期通过转录调控因子Cyc基因直接上调GABA受体GRD亚基的表达,增加GABA的合成并限制其分解,不断促进GABA-ergic信号发挥作用,最终抑制(延迟)DH 的释放,降低DH的滞育诱导作用。这项研究证实了Per基因敲除后改变了家蚕典型的温度和光周期依赖的滞育决定,并通过GABA-DH 神经递质-内分泌激素途径影响蛹期滞育发生,并表明了GABA受体GRD亚基由家蚕的生物钟和内分泌系统共同控制。该结果为解释家蚕生理时钟对内分泌激素的调节机制提供了一个实例。
4 鳞翅目昆虫-植物生物钟系统的协同进化
在昆虫-植物长期互作的进化史中,植物逐渐演化出躲避昆虫侵害的防御策略,同时吸引鳞翅目成虫传粉者;相对应的,幼虫演变出躲避或适应植物防御系统的机制,成虫则进化出了高效获取食料的结构[14]。在这个生存游戏中,植物和鳞翅目昆虫的生物钟系统似乎在控制不同活动/生理现象的时序上发挥了重要作用。
在植物-昆虫协同进化研究中,两者在生态学和进化上建立一种有效的互惠关系,而植物-昆虫生物钟的同步性是互惠共存的基础。例如,矮牵牛花Petunia axillaris气味释放与烟草天蛾触角感知花香信号而启动的飞行活动表现出高度同步性,从而确保了成虫高效获取蜜源及传粉(图2A)[49-50]。此外,植物-昆虫生物钟系统一方面会增强植物对害虫昼夜攻击的抵抗力,另一方面又会促使昆虫进化出躲避植物的生物钟防御策略,即在24 h内的特定时段产生有毒物质(植物)和解毒酶(幼虫)[14]。在模式植物拟南芥-粉纹夜蛾Trichoplusia ni互作研究中,拟南芥通过生物钟介导茉莉酸积累的模式与取食行为节律一致以抑制害虫取食活动(图2B)[51]。而鳞翅目夜蛾科昆虫为避免寄主植物防御策略演化出“白天代谢解毒,夜间取食”的节律性取食活动[18,20,51-52]。在斜纹夜幼虫“取食-解毒代谢”研究中,幼虫中肠和脂肪体中成簇的解毒基因在24 h内呈现出“白天高,晚上低”的震荡表达,使得幼虫对烟碱类杀虫剂的敏感性呈现出时节性差异,其中Clk,Bmal1,Per和Cwo等钟基因在其解毒基因震荡表达调控中发挥重要作用(图2C)[18]。由此可见,斜纹夜蛾生物钟基因参与了自身对异生物质的解毒代谢调控,研究结果为进一步解析斜纹夜蛾昼夜习性和农药耐受能力提供了理论基础,同时为农药喷施的时机选择提供了参考,为夜蛾科害虫精准防控开拓了新思路。例如,利用人工饲料饲养的烟草天蛾,其幼虫取食昼夜节律的丧失目前被认为是一种有效降低自然寄主烟草Nicotiana attenuata昼夜防御机制负面影响的生存策略[14]。
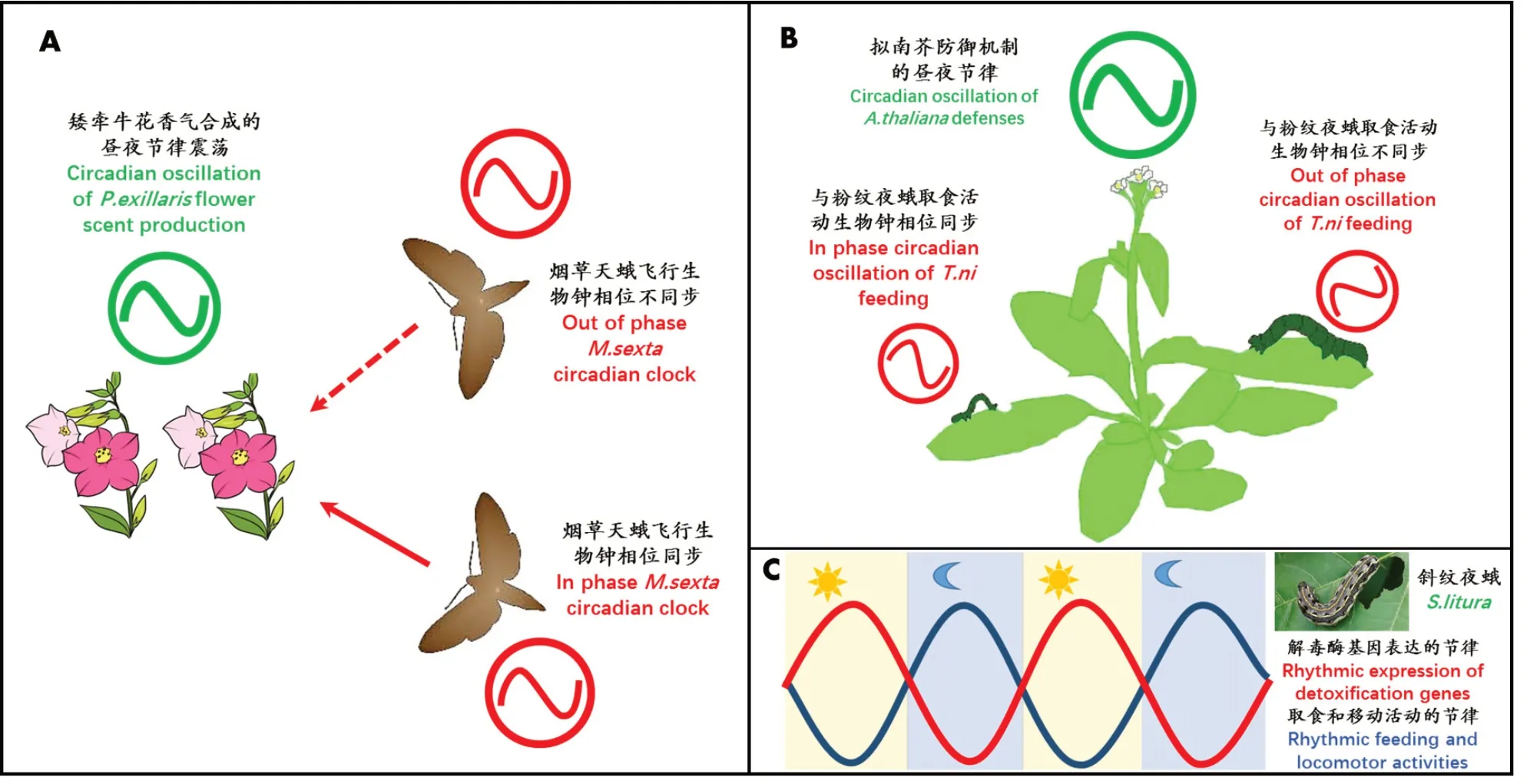
图2 鳞翅目昆虫与植物相互作用的生物钟调控事件
5 展望
近年来,生物钟生物学研究一直是科学界备受瞩目的前沿课题,其研究成果常常发表于Nature、Science和Cell等顶尖期刊。随着时间的推移,生物钟的研究愈加深入,揭示了越来越多的复杂机制,令人振奋的是,该领域渐渐重视多学科交叉渗透与协同,深入浅出地揭示自然生物“守时”的奥秘。然而,与现存众多的鳞翅目物种相比,人们对鳞翅目昆虫生物钟的研究微乎其微,尤其是农业害虫生理行为的生物钟调控机制及害虫遗传防治技术研究等方面。近期,鉴于生物钟对鳞翅目昆虫解毒和代谢等生理功能的调控作用,科学家们开始重视昆虫时间毒理学的研究,以期将生物钟理论应用于农业害虫防治生产实践,为促进农业害虫综合防控提供科学依据和新的有效抓手。近年来,鳞翅目昆虫昼夜节律授时机制虽然取得了较大进展,但主要集中在核心生物钟调控方面,对于普遍存在于其他组织中的外周生物钟研究还非常有限,外周生物钟的调控机制及其与中枢生物钟系统的互作机制研究将是生物钟研究的重要发展方向。另外,生理时钟和内分泌系统在生理行为上的协同调控仍是一个耐人寻味的话题,而目前家蚕滞育的生物钟-内分泌激素协同调控机制研究成果为昆虫滞育的研究开辟了新思路。
生物钟除了陚予生物对环境的适应能力外,同时也能对环境与生态产生重要影响[53]。迄今为止,许多研究已道明了昆虫生物钟系统的重要生态学意义。在自然界中,鳞翅目昆虫生物钟系统不仅是维持个体正常生命活动的关键,同时通过植物-昆虫长期的“攻守”互作而协同进化,进而影响整个生态系统物种多样性的发展。近期,基于生物钟系统的昆虫-植物互作机理研究为阐明植物防御与昆虫反防御的内在调控机制提供了新的线索和研究方向。不难预测,下一代组学和基因组编辑技术的蓬勃发展将极大促进模式昆虫和非模型昆虫时间生物学的研究和应用,进一步加深人们对生物环境适应与进化途径的理解。此外,家蚕作为最早实现人工驯化和饲养的经济昆虫,同时是后基因组时代用作功能基因分析的重要鳞翅目模式昆虫[54],近年来基于ZFN、EEN、TALEN 及CRISPR/Cas9 系统介导的基因组靶向编辑技术的快速发展,使得家蚕基因组靶向编辑技术的研究和应用取得了一系列重要进展[38-39,47]。不难预见,利用家蚕基因组靶向编辑技术,将有望促进蚕丝遗传改良、丝腺生物反应器开发等产业瓶颈问题的突破[55]。
