氮肥后移对中、弱筋小麦籽粒淀粉合成及其糊化特性的影响
2023-10-10吴东明韩雅楠马宏亮祁鹏飞魏育明樊高琼
吴东明,韩雅楠,马宏亮,祁鹏飞,魏育明,樊高琼,刘 琼,郑 亭
(四川农业大学农学院,四川 成都 611130)
西南冬麦区是我国继黄淮海和长江中下游地区以外的第三大小麦优势产区,主要以四川为主,属于中、弱筋型小麦优势产区[1-3]。施氮是小麦生产中重要的栽培措施,直接影响小麦的品质[4-6]。适当增施氮肥可显著提升小麦籽粒加工品质,而合理安排追施氮肥需结合品种筋型特征才能最大限度地发挥品种品质优势,真正做到产、质协同。分次追施氮肥不仅能满足小麦不同时期生长发育需求,有效地降低氮肥损失[7-8],更能精准调控籽粒内含物积累,改善其加工品质。
小麦籽粒主要由淀粉和蛋白质组成,其中淀粉和蛋白质分别占成熟籽粒干质量的60%~65%和8%~15%[9-11]。目前,大多数有关施氮时期对小麦籽粒加工品质的研究集中于蛋白质,而对淀粉的相关研究较少,并且结论并未统一。盛靖等[12]研究认为,施氮量180 kg·hm-2条件下,随氮肥后移,皖麦直链淀粉、支链淀粉、总淀粉含量及直/支比均下降,但淀粉峰值粘度升高,从而加工品质得到改善。郭天财等[13]认为随施氮肥时期的后移,小麦籽粒直链淀粉含量下降,而支链淀粉、总淀粉含量及淀粉糊化特性则以拔节期追氮处理最高。李春燕等[14]研究表明,增加后期氮肥追入量后,小麦灌浆期籽粒SSS、ADPGase、GBSS活性增加,总淀粉和直、支链淀粉积累速率增加,最终淀粉积累量提高。由此可见,合理追施氮肥对淀粉理化特性起着至关重要的作用。
基于此,试验以大面积推广的四川中、弱筋小麦品种为研究对象,运用现代分子生物学及生理生化技术,研究施氮时期对小麦籽粒淀粉的理化特性、淀粉合成相关酶活性以及关键酶基因表达的影响,解析小麦淀粉及其组分变化的调控机理,以期进一步探讨四川地区中、弱筋专用小麦品质形成相关理论,并为当地优质专用型中、弱筋小麦生产提供依据。
1 材料与方法
1.1 试验地概况与试验设计
试验于2014年10月—2015年5月与2015年10月—2016年5月连续两年在崇州市四川农业大学现代化农业研发基地(川西平原地区:30°58′N,103°53′E)和眉山市仁寿县珠嘉乡踏水村(盆中丘陵地区:30°04′N,104°13′E)进行。两地均属于亚热带湿润气候,两个播种点的土壤基础理化特性如表1所示,小麦生育期气象条件见图1。

图1 崇州和仁寿2014—2015年和2015—2016年气象数据图Fig.1 Chongzhou and Renshou meteorological data during 2014-2015 and 2015-2016 wheat growing seasons

表1 两个地点试验田的土壤基础理化性质(2014年)Table 1 Physical and chemical properties of the soil in two trial sites in 2014
采用二因素裂区试验设计,主区为品种,供试品种为中筋小麦‘蜀麦482’(SM482)和‘蜀麦969’(SM969)、弱筋小麦‘川农16’(CN16)和‘绵麦51’(MM51)。其中‘绵麦51’由四川省绵阳市农业科学院提供,其余3个品种由四川农业大学小麦研究所提供;副区为施氮时期,施氮时期分别为底肥一次性施用(T1)、底肥+拔节期追肥(T2)、底肥+孕穗期追肥(T3)。全生育期总施氮量为纯氮150 kg·hm-2,除底肥一次性施用外,其余处理按底肥∶追肥=60%∶40%(简记为6∶4)施用;每公顷施用P2O5和K2O 各75 kg,全作底肥一次施入;供试肥料种类为尿素(N 46.2%)、过磷酸钙(P2O512.5%)和氯化钾(K2O 60%),购于当地农资市场。其他栽培措施与一般大田生产相同。于2014年10月28日和2015年10月26日播种。行窝距为20 cm×10 cm,播种量为1.8×106kg·hm-2,小区面积为4 m×3 m,试验重复3次。
1.2 测定项目与方法
1.2.1 田间标记与取样方法 在小麦开花期对同一天开花、长势一致的穗进行挂牌标记。从花后10 d起开始取样,每隔5 d取样1次直至成熟。每次取18穗,在液氮中剥取籽粒,一部分鲜样迅速放入液氮中,然后冷藏于-80℃超低温冰箱,用于测定酶活性、提取RNA;其余籽粒样品在105℃下杀青30 min,70℃烘至恒重后用于测定淀粉含量。
1.2.2 淀粉含量的测定 淀粉含量采用Megazyme公司的总淀粉测定试剂盒测定,按照说明书进行操作。
1.2.3 关键酶活性的测定 粗酶液的提取:取20 粒种子称重,研磨成粉,边加液氮边磨,防止酶失活。然后加4 mL提取液(含100 mmol·L-1HEPES-NaOH (pH=7.5)、8 mmol·L-1MgCl2、2 mmol·L-1EDTA、12.5% (V/V) 甘油、1% (W/V) PVP-40、50 mmol·L-12-巯基乙醇),研磨成匀浆,10 000 r·min-1下离心30 min,然后收集上清液置于冰浴,用于酶活性的测定。
AGPase活性的测定参照程方民等[15]的方法,略有改动;SSS活性和GBSS活性测定参照梁建生等[16]的方法。
1.2.4 糊化特性(RVA参数)的测定 根据AACC76-21方法,利用澳大利亚NewPort快速粘度分析仪测定糊化特性。
1.2.5 提取总RNA 取10~15粒大小一致的籽粒放入预冷的研钵中边加液氮边进行研磨,将籽粒磨成粉末,然后利用植物RNA提取试剂盒(百菲特公司)提取总RNA,按照说明书进行操作。提取后进行琼脂糖凝胶电泳,检测RNA质量,完整的小麦籽粒RNA在18 S和28 S会有两条清晰的条带。最后在紫外分光光度计上测定 260 nm/280 nm 吸光值确定RNA 浓度和纯度。
1.2.6 cDNA的生成 取RNA 1 μl按照PrimeScriptTM RT Reagent Kit with gDNA Eraser(Perfect Real Time) 试剂盒(TaKaRa)的说明书进行操作。
1.2.7 淀粉合成关键酶基因引物的设计 利用Primer premier 5.0 引物设计软件,根据美国国立生物技术信息中心(NCBI)公布的AGP-L、Wx-DI、Sbe1D基因序列设计引物,由生工生物(Sangon Biotech)公司负责合成。具体引物序列如表2。

表2 qPCR引物序列Table 2 Primer pairs used for qPCR analysis
1.2.8 荧光定量RT-PCR 将上一步所获得的cDNA用dd H2O稀释10倍,用于接下来的qPCR 反应中。
qPCR分析使用上海BioRad公司的荧光仪器IQ 5(96孔)和大连宝生物公司荧光试剂SYBR Premix EX TaqII,按照说明书进行操作。具体体系配置以及扩增程序如表3所示。

表3 反应体系Table 3 Reaction system
在qPCR反应中,小麦样品的3个内参基因分别是甘油醛-3-磷酸脱氢酶(w-GAPDH, NCBI unigen Ta.66461), Heterogeneous nuclear ribonucleoprotein Q (w-HnRNPQ, Ta.10105) 和Scaffold-associated regions DNA binding protein (Ta.14126)。采用ΔΔCT法利用CT值和扩增效率(E值)计算相对丰度。
1.3 数据处理
试验数据通过Microsoft office excel汇总整理后,采用DPS 15.05进行统计分析处理。
2 结果与分析
2.1 施氮时期对淀粉及其组分含量的影响
两试验点结果均表明,施氮时期显著影响总淀粉和直链淀粉含量及直/支比,但基因型和两者的交互作用对其影响较小(表4)。而支链淀粉含量变化较小,支链淀粉主要取决于基因型。施氮时期分别可解释总淀粉含量、直链淀粉含量和直/支比23.9%~39.0%、37.7%~45.1%、24.3%~29.0%的变异。随着氮肥后移(T1~T3),4个品种的单粒淀粉积累量均增加,但淀粉含量在2个试验点均表现出显著下降规律,总淀粉、直链淀粉含量分别显著下降0.20~1.75个百分点、0.10~1.85个百分点,直/支比下降了4.88%~10.42%。相比仁寿点,崇州点总淀粉含量、直链淀粉含量、直/支比较低。氮肥后移后,2个试验点支链淀粉表现相反,崇州点略微下降,仁寿点略微上升,由此说明氮肥后移对支链淀粉的影响效力大小可能受环境控制。
2.2 施氮时期对淀粉糊化特性的影响
由表5、表6得出,2015年和2016年2个试验点试验结果基本一致,施氮时期对淀粉糊化特性的影响较小,相比崇州点,仁寿点各糊化特性指标数值较高。两年2个试验点施氮时期可解释糊化特性0.01%~13.5%的变异,远低于基因型解释4.21%~94.98%的变异,略低于交互作用解释的0.50%~15.76%的变异。‘CN16’和‘MM51’的峰值粘度、低谷粘度和最终粘度在T1、T3之间差异显著(P<0.05)。整体上,4个品种在不同追氮时期下表现出相同的变化趋势,即随追肥时间的推迟,糊化特性指标基本均有所提升。特别是峰值粘度(P1)、低谷粘度(T1)、最终粘度(FV) 和回复值(SB),随施氮时期后移,两年2个试验点分别提升3.3%~11.9%、3.8%~26.1%、3.0%~20.5% 和1.6%~15.0%。综上可知,推迟追肥时间显著提升淀粉的糊化特性数值。
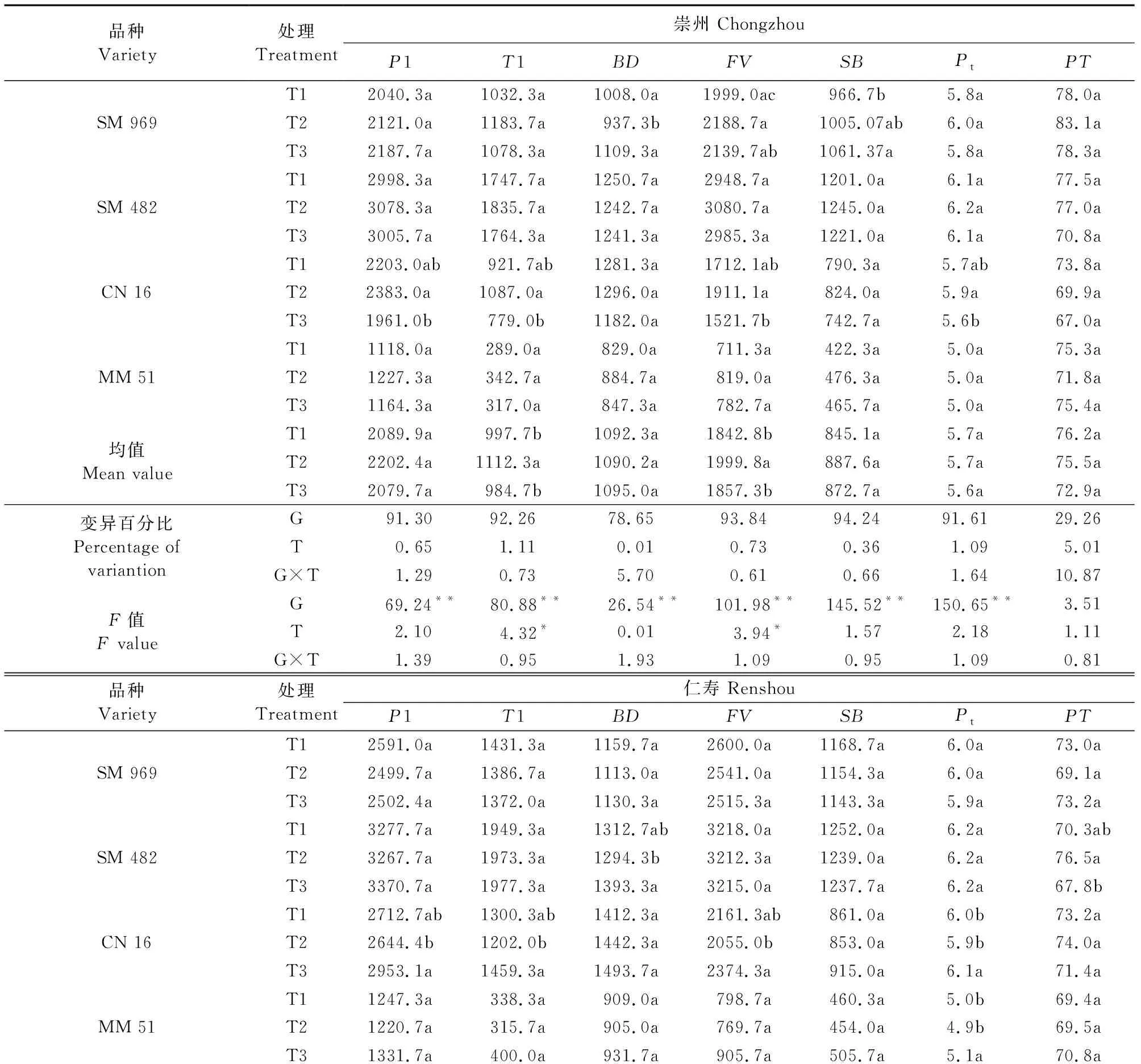
表5 不同追氮时期下成熟期小麦籽粒淀粉糊化特性(2015年)Table 5 Starch pasting characteristics of mature wheat grain at different nitrogen topdressing stages in 2015
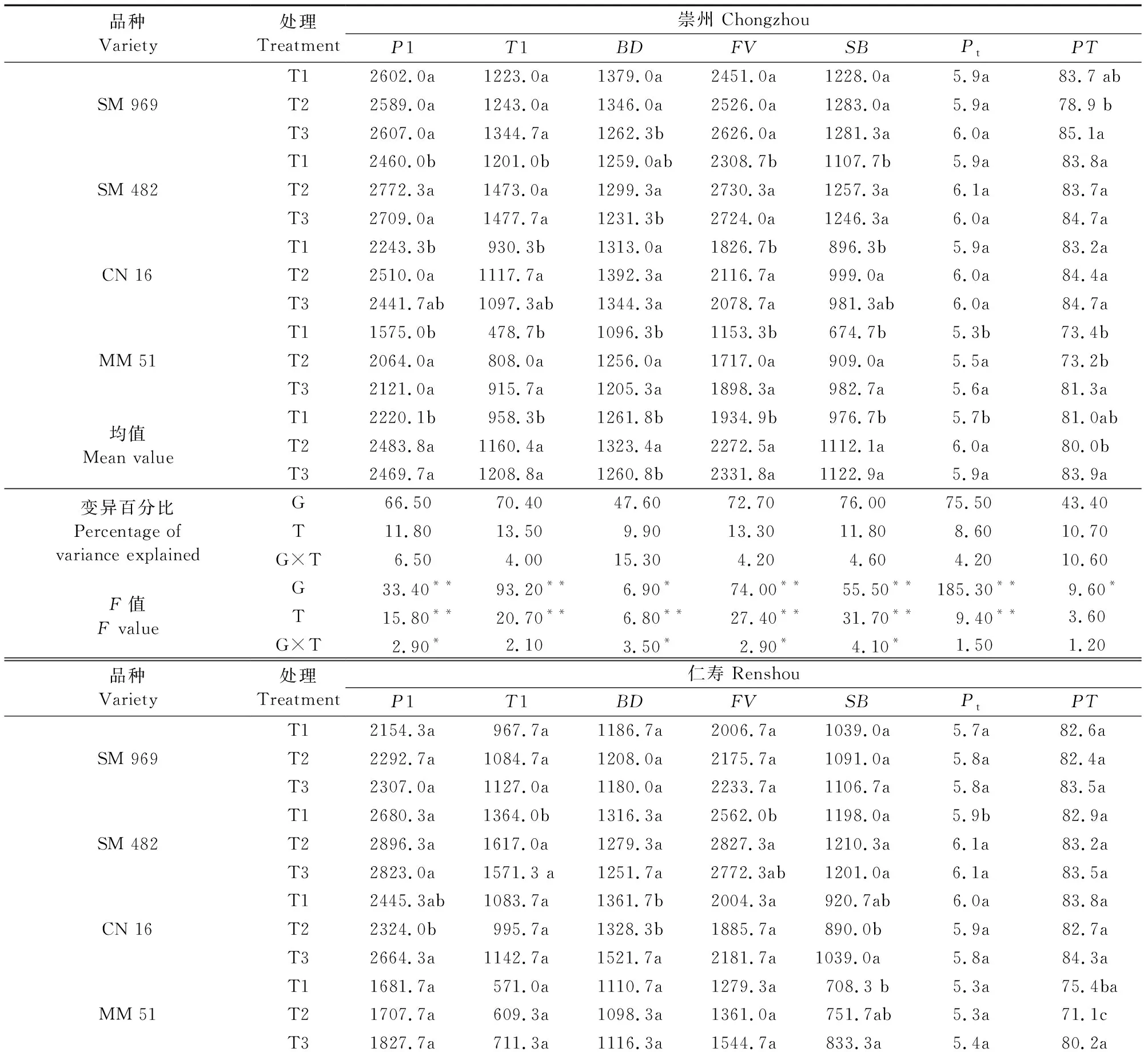
表6 不同追氮时期成熟期小麦籽粒淀粉糊化特性(2016年)Table 6 Starch pasting characteristics of wheat grain during maturity under different nitrogen topdressing stages in 2016
2.3 糊化特性与淀粉含量及其组成比例间的相关分析
由表7得出,总淀粉、支链淀粉基本与糊化特性呈较强的正相关关系,直链淀粉和直/支比与糊化指标大多呈负相关关系,但直链淀粉和直/支比与PT在两生态点均呈较强的正相关关系。FV、SB、Pt与支链淀粉的相关系数较大,其中支链淀粉与SB的相关性在2个试验点均达显著水平。淀粉相关指标中,支链淀粉与糊化指标的相关性最大。

表7 淀粉含量与糊化参数相关性分析(2015年)Table 7 Correlation analysis of starch content and viscosity characteristics (harvest in 2015)
2.4 施氮时期对淀粉合成关键酶活性的影响
通过对崇州点2016年淀粉合成关键酶活性的测定(图2~图4)发现,AGPase、SSS以及GBSS活性在灌浆期总体上呈先上升后下降的单峰曲线,在花后25 d或30 d达到峰值。‘SM482’和‘MM51’追肥后移AGPase、SSS、GBSS活性在花后15 d内无显著差异,在灌浆中后期(花后25 d后)酶活性的提高幅度更大,差异更明显。但‘SM482’和‘MM51’的SSS、GBSS活性随施氮后移的响应略有差异。施氮后移,蜀麦482的SSS酶活性灌浆中后期(花后25 d)基本表现为T3> T2>T1,‘MM51’的SSS活性在灌浆中前期(花后20 d)受到抑制,表现为T1>T2、T3,灌浆后期(花后35 d)为T3>T2>T1。两品种GBSS活性在T2处理下灌浆中期(花后25 d)即显著大于T1,在T3处理下灌浆后期(花后35 d)显著大于T1。‘SM482’在氮肥后移后,相比‘MM51’的GBSS酶活性提高的幅度更大。

注:误差线表示3次重复的标准误, 其上所标不同字母表示同一天处理间差异显著(P<0.05)。下同。Note: Error bars represent standard errors of three replicates. Different letters above error bars indicate significant differences among treatments (P<0.05). The same as blew.图2 花后籽粒AGPase活性变化Fig.2 Changes of AGPase activity in grains after anthesis
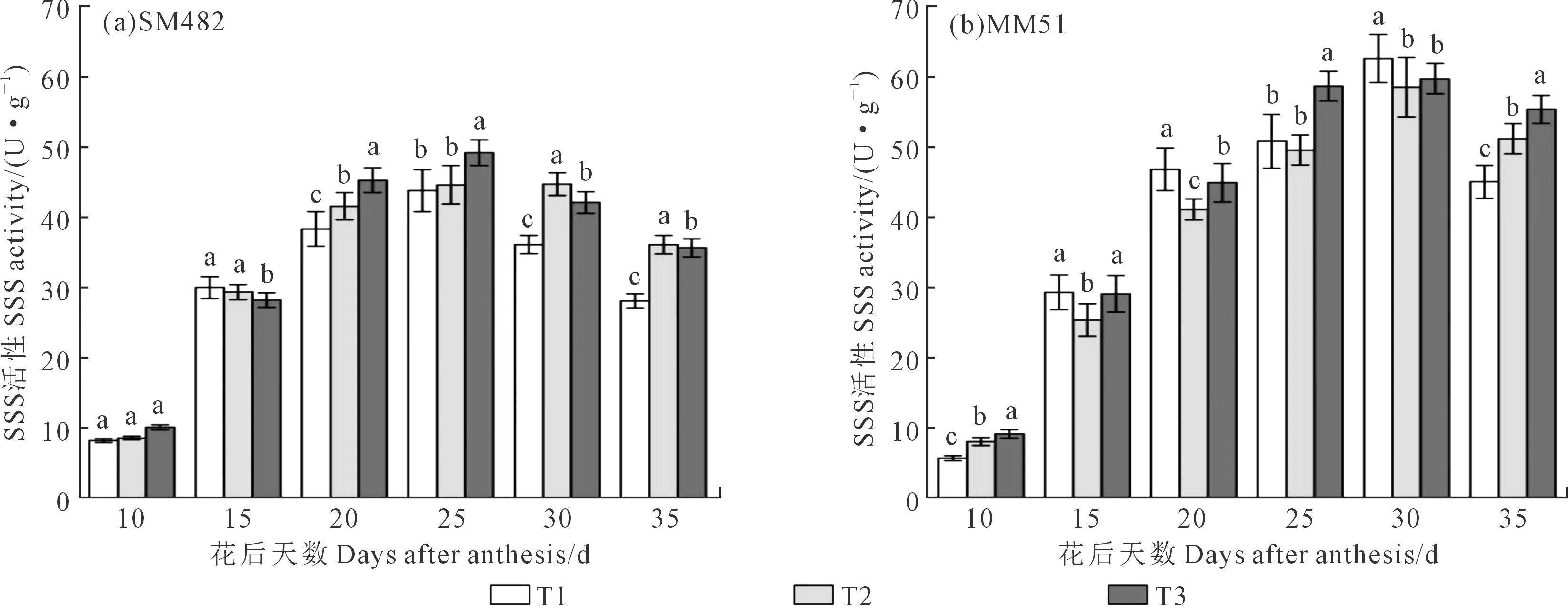
图3 花后籽粒SSS活性变化Fig.3 Changes of SSS activity in grains after anthesis

图4 花后籽粒GBSS活性变化Fig.4 Changes of GBSS activity in grains after anthesis
2.5 施氮时期对淀粉合成关键酶基因表达量的影响
通过对崇州点‘SM482’进行RNA-seq(T1 vs T3)分析发现了AGP-L、Sbe1D和Wx-DI这3个分别负责编码AGPase、SSS和GBSS的DEGs,随即进行了qRT-PCR验证(图5)。结果表明,AGP-L和Wx-DI表达量在花后20 d 最高,AGP-L在T3处理下整个灌浆期均受到抑制。Wx-DI在T3处理下花后10~20 d表达量上升,花后15 d显著上调了0.66倍;花后20~35 d表达量受到抑制,花后25、30、35 d分别显著下调0.18倍、0.15倍、0.32倍。Sbe1D的表达量在花后25 d达到最大,25~35 d表达量受到抑制,花后30、35 d显著下调了0.19倍和0.22倍。

图5 AGP-L、BE1D、Wx-DI基因的相对表达量Fig.5 Relative expression of AGP-L、BE1D、Wx-DI gene in developing grain
3 讨 论
3.1 直/支比下降是造成氮肥后移糊化特性变化的主要原因
面条加工品质和食用品质与小麦淀粉的糊化特性有很大联系。研究发现小麦淀粉的糊化温度、峰值粘度和崩解值越大,面条的质量越好[17-18],淀粉峰值粘度和面条硬度呈负相关关系[19],淀粉的回复值与面条的硬度呈正相关关系[20]。同时,糊化特性又与淀粉含量及其组成高度相关,高崩解值和低回复值表明直链淀粉含量偏低,高峰值粘度表明支链淀粉含量和直/支比较高[21-22]。研究发现,直链淀粉含量较低的品种在面条软度、黏性、光滑性、口感和综合评分等品质参数上有较好的表现;支链淀粉的含量与面条评分的各项指标和总分成正相关关系[23]。 所以淀粉组成及糊化特性是评价面粉是否适宜制作面条的重要参考标准[24-26]。
目前,氮肥后移对淀粉组成及糊化特性的影响尚无定论。郭天财等[13]、赵庆玲等[27]认为随施氮时期的后移,中强筋小麦籽粒直链淀粉含量下降,总淀粉和支链淀粉含量均为拔节期追氮处理最高。胡宏、吴进东等[28-29]认为弱筋小麦在高氮和低氮处理下,随氮肥后移总淀粉和支链淀粉含量均增加,但直链淀粉含量在高氮处理下呈下降趋势。但陆增根等[30]认为增加后期的施氮比例可显著提高直链淀粉的含量以及直/支比。郭天财等[13]认为在拔节期追氮时小麦籽粒的支链淀粉、总淀粉含量最高,淀粉糊化特性最优。但也有研究发现增加后期施氮比例使支链淀粉的含量呈降低趋势[22]。阎俊、张学林等[31-32]发现,追施拔节肥有利于提高强筋、中筋、弱筋小麦的糊化特性,尤其是提高了峰值粘度、最终粘度和稀懈值,从而提高了淀粉的糊化特性、改良了面条的品质。本试验结果表明随追肥时期推迟,总淀粉含量、直链淀粉含量和直/支比下降,淀粉糊化特性提升,尤其是峰值黏度、低谷黏度、最终黏度和回复值显著上升。由此可见,追肥时期推迟有利于改善淀粉糊化特性,与张学林和阎俊等[31-32]结论相符,淀粉组分的变化同郭天财[13]和胡宏[28]等人的部分结论相符。前人的研究认为,直链淀粉含量与峰值粘度、稀懈值等参数呈显著负相关关系[33-34],这与本文的结论基本吻合。本研究中峰值黏度、低谷黏度、最终黏度和回复值与直链淀粉以及直/支比呈负相关,与支链淀粉呈显著正相关关系。由此推测,追氮时期后移糊化特性提升是直链淀粉含量和直/支比下降的结果,从而有利于中、弱筋小麦蒸煮品质的提升[35]。同时,弱筋既要求淀粉含量较高,又要求蛋白含量较低,但追肥后移会显著提高籽粒蛋白含量,所以弱筋小麦追肥不宜太迟。
张铭等[36]研究发现提高土壤肥力可以降低直/支比,由表1可知仁寿的土壤肥力低于崇州,这是崇州点各处理小麦直/支比低于仁寿的部分原因。同时张学林等[37]研究了不同生态点不同筋型对淀粉糊化特性的影响,发现日照较多有利于糊化特性的提高,而降雨过多则作用相反;渍水条件下,支链淀粉含量显著下降[38]。由图1可以看出,仁寿的降雨量低于崇州,这可能是崇州点和仁寿点2015年淀粉糊化特性高于2016年,以及仁寿点糊化特性高于崇州点的部分原因。两年两试验点间淀粉糊化特性的结果不完全一致,说明环境的差异对糊化特性及淀粉组分均有较大影响,并且氮肥后移对淀粉合成的影响效力大小受如温度、土壤肥力和水分等环境影响[39]。
3.2 灌浆中后期关键淀粉酶合成相关基因的表达可直接响应氮肥后移对淀粉的调控
研究认为[40-42]AGPase可以催化ADPG的生成,从而控制淀粉的合成与积累,SSS和SBE基因在支链淀粉的生物合成中起着不可或缺的作用,当它们发生突变,相关酶的活性和支链淀粉的含量会相应降低,胚乳中直链淀粉的含量直接由GBSSI基因的转录水平决定,而直链淀粉的生物合成受GBSSII基因控制[43-44]。灌浆期小麦籽粒淀粉的合成与AGPase、SSS、GBSS以及SEB的活性成正相关关系,影响着淀粉的积累动态[45-47]。淀粉合成通路中AGP-L、Sbe1D、Wx-DI基因分别负责编码AGPase、SSS、GBSS,任何一个编码淀粉酶的基因亚型的变化都可能导致淀粉特性发生变化[48-50]。谭彩霞等[50]认为这3个基因的相对表达量在灌浆期峰值与相对应的酶活性值呈现极显著正相关关系。
目前,有关氮素对淀粉合成关键酶基因表达量的研究多集中在施氮量上[51],而对氮肥后移的研究较少[48],对淀粉合成关键酶基因表达量、淀粉合成关键酶及总淀粉、直链淀粉、支链淀粉的影响尚无定论。谭彩霞等[50]认为在小麦籽粒灌浆后期适当增加追施氮肥比例可以提高淀粉合成关键酶基因表达量,直、支链及总淀粉积累量均呈现上升趋势。本文中灌浆期淀粉合成相关基因峰值表达时期与Morell、Ran等[52-53]的研究结果一致,AGP-L和Wx-DI基因在灌浆中期高效表达,Sbe1D在中后期高效表达,花后20 d达到最大表达水平。同时,氮肥后移后,籽粒灌浆中后期AGPase、SSS、GBSS的活性显著提高,与底肥一次施用处理差异更明显,这与谭彩霞等[50]的研究结果基本吻合。但基因表达量除了Wx-DI在中前期表达量明显提高外,中后期淀粉合成相关基因都受到抑制,与酶活性的趋势相反。这可能与AGPase、SSS、GBSS均由多个亚基控制有关[54]。与此同时,总淀粉、直链淀粉含量和直/支比下降,这与中后期淀粉合成相关基因的表达量变化趋势一致。此外,氮肥后移后AGPase酶活性变化趋势与总淀粉含量变化不一致,可能是氮肥后移延缓了叶片衰老,同时增强籽粒中蛋白质的合成与积累,提高千粒重,淀粉积累总量增加,但籽粒淀粉的含量相对降低[14,55]。因此,推测灌浆中后期关键淀粉合成相关基因的表达可直接响应氮肥后移对成熟籽粒淀粉积累量的调控,而酶活性的变化则比较滞后。
4 结 论
随施氮后移,AGP-L、Sbe1D和Wx-DI基因表达量在小麦灌浆中后期受到抑制,总淀粉含量、直链淀粉含量和直/支比下降,淀粉糊化特性有显著变化,均有利于中、弱筋小麦蒸煮品质的提升。小麦淀粉品质是地点、品种共同作用的结果,应根据不同品种、地点和最终用途采取适宜追肥方式调节小麦淀粉品质。综合来看,四川盆地弱筋品种追肥不宜太迟,适宜底肥一次性施用或者拔节前追肥;中筋品种可考虑追施拔节肥或孕穗肥。
