自噬在卒中后抑郁中的作用及中药干预机制的研究进展
2023-10-09陈景云巩婷李妍怡苟梅丽张静惠渊周学锐
陈景云,巩婷,李妍怡,苟梅丽,张静,惠渊,周学锐
(1. 甘肃中医药大学,甘肃 兰州 730000; 2. 甘肃省中医院,甘肃 兰州 730050)
卒中后抑郁(post-stroke depression,PSD)是一种发生在脑卒中后的神经精神疾病[1],是脑卒中常见的并发症,具体表现为兴趣淡漠、情绪低落、食欲不振或暴饮暴食、注意力不集中等[2]。据统计目前PSD的患病率为31.1%,且呈逐年上升趋势,极大地降低了卒中患者的生活质量,成为全球所面临的共同的医疗难题[3-4]。调查发现,卒中后抑郁发生的影响因素主要有年龄、性别、文化程度、家庭年收入、睡眠障碍等[4],卒中前抑郁、肢体残疾、认知障碍和卒中严重程度是PSD 最一致的预测因素,其发病机制尚不明确,比较公认的机制有炎症反应机制、神经网络破坏、遗传易感性、神经营养因子改变和去甲肾上腺素、神经生物学改变等[1]。自噬可通过溶酶体分解代谢途径降解和回收寿命过长及错误折叠的蛋白质和受损的细胞器,以维持细胞内稳态和正常的功能[5]。自噬与气虚痰瘀、中医阴阳理论的相关性研究备受关注,因此深入研究中医药对PSD 干预自噬治疗,对PSD的防治有重要意义,现将自噬在PSD的作用及中医药干预自噬治疗PSD 的相关研究进展综述如下。
1 细胞自噬
1.1 自噬的细胞类型
哺乳动物真核细胞自噬类型主要有巨型自噬、微自噬以及伴侣介导的自噬,无论何种类型的自噬都涉及细胞质物质,严格依赖溶酶体并终将被溶酶体所降解。微自噬是指胞质物质或者细胞器质被溶酶体或液泡膜进行降解的过程。伴侣介导的自噬(chaperonemediated autophagy,CMA)只在哺乳动物细胞中被提出过,与微自噬和宏自噬非特异性地吞噬大量细胞质不同,CMA 是高度特异性的。五肽靶向是所有CMA 的底物,与KFERQ 生化相关,KFERQ 共识基序的靶蛋白通过胞质伴侣蛋白的作用并直接跨过溶酶体膜,在管腔中被降解。在过程中,KFERQ 基序被热休克70 kDa 蛋白8(HSPA8/HSC70)识别,然后HSPA8将底物输送到溶酶体膜上。在溶酶体膜上底物与CMA底物受体的单体结合产生跨膜蛋白LAMP2A。随着多聚体易位复合体的形成,复合体的亚单位被HSP9(8)稳定在溶酶体膜的管腔一侧。在底物移位到溶酶体腔内后,部分通过管腔HSPA8 的作用,转位复合体被胞浆HSPA8 主动分解;相关研究较多的典型自噬是巨自噬,这种自噬是指在饥饿、应激等刺激诱导下,自噬体是通过细胞质内的双层囊泡结构包裹非特定蛋白产生的,随后自噬体与溶酶体/液泡进行融合,而其中自噬体包裹的物质最终被溶酶体/液泡里的酸性水解酶降解的过程[6-7],这是通常意义上的自噬。自噬的三种类型及分子过程见图1。

图1 自噬的三种类型及分子过程
1.2 自噬的过程及相关的蛋白调控
典型的自噬反应可概略地分为五个阶段,分别为启动、噬菌体成核、噬菌体扩张和底物选择、自噬小体-溶酶体融合和溶酶体底物降解,自噬是由VPS15、VPS34、ATG14 和Beclin1 组成的Ⅲ型P13-K 复合体介导的双膜结构的吞噬或隔离膜的形成而启动的,然后噬菌体拉长并吞噬要降解的物质。随后,自噬小体本身形成,伴随着ATG4对LC-Ⅱ的脱脂作用,自噬小体的外膜与溶酶体融合。最后,溶酶体水解酶降解内膜和溶酶体含量,简单的降解产物如氨基酸、脂肪和糖用于合成和生产三磷酸腺苷等物质。
自噬最开始是在酵母中被发现,是真核细胞中一种保守的分解代谢过程,参与了细胞器和细胞质的靶向降解。自噬是对大量生理和病理刺激的反应,有助于维持胞质内的动态平衡,即蛋白质、核酸和许多细胞器的合成和降解之间的平衡,对酵母的研究大大加深了对自噬过程的了解,并揭示了酵母和哺乳动物自噬之间的差异。例如,在哺乳动物中没有发现自噬前结构。哺乳动物细胞中的自噬小体可能来自不同的细胞膜结构,如内质网、质膜或线粒体外膜。自噬在胚泡着床、胚胎发育、红细胞成熟、抑制癌变、抗衰老、清除病原微生物等多种生理过程中发挥着重要作用。然而,自噬也参与了几个病理过程,包括肿瘤、肺纤维化及阿尔茨海默病或帕金森病中出现的神经退化等[8-10]。研究发现,ATG 复合体——ATG 蛋白,在动植物细胞自噬过程中发挥着不可或缺的作用。在哺乳动物中,ATG9是真核细胞自噬过程中唯一的跨膜蛋白,它为形成自噬前结构(PAS)和驱动隔离膜的延伸提供了膜来源。ATG2(哺乳动物中的ATG2A)与ATG18 哺乳动物中的WD-Repeat 蛋白与磷脂酰肌醇相互作用,作用于吞噬小体形成的后期阶段,将PAS/隔离膜固定在内质网上并转移脂类,有效地完成闭合过程。磷脂酰肌醇-3-激酶(PI3K)复合体PI3K 复合体介导小泡成核。PI3K 复合体可进一步分为复合体Ⅰ和Ⅱ,复合体Ⅰ由ATG6、ATG14L、VPS15 和VPS34 组成,复合体Ⅱ由ATG6、VPS15、VPS34 和VPS38 组成。PI3K 复合体和磷脂酰肌醇-3-磷酸(PI3P)负责修饰噬菌体的从头合成,PI3P 将ATG2-ATG18 复合体招募到吞噬体膜上,并参与吞噬体的延伸ATG5/ATG12、ATG8-磷脂酰乙醇胺(PE)结合系统ATG5-ATG12 和ATG8-PE 类泛素结合系统,不仅调节噬菌体形成的起始,而且在下游步骤发挥作用。其中,ATG4是一种关键的半胱氨酸蛋白酶,对ATG8(哺乳动物中LC3)的脂化和ATG8 的自由翻转具有重要的功能。在自噬过程中,ATG8 的C 末端首先被ATG4 酶识别和切割。随后,具有泛素激活酶E1 活性的ATG7 与ATG8 和ATG12 结合,激活其成熟形式。激活的ATG8 和ATG12 分别转移到具有泛素结合酶E2 活性的ATG3 和ATG10 上,并最终连接到底物上。此外,ATG12-ATG5 加合物与ATG16(哺乳动物中的ATG16L1)结合形成低聚复合体,从而促进ATG8-PE加合物的形成[11]。
2 中医学与细胞自噬和PSD
“阴平阳秘、阴阳自和”理论与细胞自噬在生理、病理状态下维持机体的稳定及自愈功能相一致,与“肾虚肝郁”相联系。目前对于PSD 的中医的病因病机及中医药治疗研究逐渐增多,这为进一步研究中医药对细胞自噬的调控作用从而治疗PSD 提供了较好的理论基础。
2.1 阴平阳秘与细胞自噬
“阴平阳秘”是机体内阴阳互相制约使机体的活动和功能处于一种有序的动态平衡,是最佳的机体健康状态[12]。《素问·阴阳应象大论》曰:“阴在内,阳之守也,阳在外,阴之使也”,故在人体正气充盛的自然状态下,阴阳的相互制约维持了机体内环境的稳定,使机体处于一种自我稳定的平衡状态。这与自噬的正常功能相一致,自噬的正常功能是通过蛋白调控将胞内物质输送到溶酶体中并在溶酶体中降解,此降解不是简单地消除物质,而是一种细胞内的动态循环的降解系统,从而维持了细胞内环境动态稳定[13]。故“恬淡虚无,真气从之,精神内守,病安从来”,人体阴阳平衡,达到形神一体的健康状态。
2.2 阴阳自和与细胞自噬
《伤寒论·辨太阳病脉证并治》曰:“凡病,若发汗、若吐、若下、若亡血、亡津液,阴阳自和者,必自愈”,在病理状态下,机体阴阳平衡的状态被打破,此时,可通过药物及自身的调控使机体重新实现阴阳平衡,自身的调控主要通过阴阳的“自我消长”“相互转化”“互根互用”特点来实现机体自身的内稳[14]。而自噬是在受到如饥饿、损伤、缺氧、感染、缺血再灌注等细胞应激反应中被诱导,将衰老或受损的细胞物质包括细胞器和蛋白质运送到溶酶体用于降解和回收,这为细胞更新和动态平衡产生新的构建块和能量[12,15],这一过程与通过相互转、此消彼长以及相互转化达到平衡的阴阳自和相一致。细胞自噬通过溶酶体实现物质的降解和再次吸收完成自我更新与“阴阳自和”的内涵相似,故从中医药调控的细胞自噬的相关机制进行研究并应用于临床,对于自噬相关性的疾病有更佳的应用前景,PSD 与自噬关系非常密切,从阴阳自和的角度调治是一种新的临床思路。
2.3 肾虚肝郁与细胞自噬
“肾主骨,生髓”“脑为髓海”,卒中后抑郁的病位在脑,其病机主要为肾虚肝郁,肾中精气不足是PSD发病的基础,肝气郁结是发病的根源。内伤积损遇劳倦内伤、情志不遂、气候骤变等因素引动痰浊、瘀血内生或阳化风动,血随气逆,导致脑脉痹阻或血溢脑脉之外发为卒中,卒中后情伤日久,肝疏泄失常,气机不畅,肝郁日久则情志内伤则随之加重,年老之人多患此病,因其年事渐高,肝之疏泄功能随之渐低,日久则肝气不舒,进则肝气郁结,故肝气郁结可视为本病最主要的根源,并贯穿脑卒中后抑郁症的全程[16-18]。有学者认为,微观自噬的产生过程与中医学中“气虚痰瘀”病机发生有一定的相关性,机体正常的生命活动的维持和机体在脏腑功能低下时通过自身调节清除内生邪使机体能够维持内环境的阴阳平衡,这个过程是机体通过“精化气”的方式完成的,这一过程与自噬存在一定的相似性[19]。机体正气不足,气机失调时易内生痰、热、火、瘀等实邪,当细胞自噬功能不足时,降解和吸收功能的减弱使衰老的细胞器、异常蛋白、代谢产生的废弃物质等产生沉积,这与机体内生痰浊血瘀相互胶着有关[20-21]。研究发现,以“肾虚肝郁”“气虚血瘀”为基础,大鼠海马神经重塑的进程被JAK2/STAT3 信号通路的过度活化可能PSD 发生的病理基础,其病机是人体肾精不足、正气亏虚从而进一步导致痰、瘀、湿、热等实邪停聚于脑[22]。对其治疗现已形成了较为完整的理、法、方、药中医治疗体系,故进一步从肾虚肝郁角度出发,深入研究中药对细胞自噬调控的相关性,对PSD 的治疗具有深远的临床及理论意义。
3 自噬对PSD的作用
自噬与PSD 密切相关,继发性损伤是脑出血患者神经系统恶化的关键因素。脑出血后继发性脑损伤是由脑实质内出血引起的,继而激活了炎症、氧化、自噬和凋亡等信号通路。在体内,这些通路成为连接脑内出血和抑郁的桥梁,参与了PSD的发生过程,而自噬参与了其发生的多个环节。
3.1 炎症反应与自噬
PSD 的炎症反应机制非常复杂,涉及多种细胞因子及蛋白质之间的相互作用。促炎症细胞因子IL-1β、IL-6、IL-8 和TNF-α 的分泌增加引起神经元变性和凋亡,而IL-6、干扰素-γ(TNF-γ)和TNF-α诱导色氨酸异常降解,并导致终末产物喹啉酸在常驻胶质细胞中积聚。通过N-甲基-D-天冬氨酸受体的过度兴奋促进谷氨酸的兴奋性毒性,并拮抗5-羟色胺,降低突触的可塑性和神经元的存活。在卒中后阶段,CX3CL1 和CD200-CD200R 的相互作用介导了胶质细胞的激活,而CCL-2 则吸引了浸润性巨噬细胞。CD206阳性细胞清除过多的危险信号;高数量的调节性T 细胞、IL-4、IL-10、转化生长因子-β(TGFβ)以及通过cAMP 反应元件结合蛋白(CREB)的细胞内信号支持M2 型分化[23]。促炎和抗炎细胞因子的动态平衡决定了炎症反应的结果和患者的远期预后[24]。自噬在PSD 炎症反应中主要与小胶质细胞中BDNF 的合成和释放与ATP 敏感的嘌呤能受体,尤其是P2X4R的激活密切相关。在脑卒中期间,来自缺血区的垂死神经元和其他细胞释放的三磷酸腺苷解决了神经胶质细胞的跨膜P2X4嘌呤能受体,这些受体的上调伴随着小胶质细胞的激活、P2X4R 触发的驻留小胶质细胞和渗透的巨噬细胞,激活的P2X4R 促进这些细胞释放BDNF。P2X4 受体的全局缺失具有神经保护作用,可以延缓神经炎症,但当缺失只影响小胶质细胞时,小胶质细胞倾向于抑郁和促炎表型。这种二分法的一个可能原因是缺乏P2X4 的小胶质细胞合成BDNF 的能力较低[23]。自噬小胶质细胞的表型变化被自噬通过核因子-κB 途径所调节,这对于缺血后神经组织恢复是有利的[25]。当自噬在一种适度的范围时可对缺血的神经组织起到保护作用,但是当自噬超过适度的范围时可能会造成细胞死亡。炎症反应在缺血后的神经细胞的存活和神经组织的恢复中起着重要作用。
3.2 氧化应激与细胞自噬
氧化应激(oxidative stress,OS)是生理氧化系统和抗氧化系统因组织或细胞中自由基的增加或清除能力的降低而紊乱,从而进一步导致体内自由基的积累而造成氧化损伤的过程[26]。在正常生理条件下,人体当中的活性氧(ROS)会被抗氧化系统及时清除,内氧化和抗氧化活性的平衡因此系统得以维持。然而,当机体受到各种应激源的刺激或受到病原体感染时,体内产生的ROS 超过细胞的抗氧化防御能力,导致氧化还原状态失衡。组织或细胞中过量的ROS 可诱导氧化应激,导致DNA 羟化、蛋白质变性和组织损伤等氧化损伤。为了防止进一步的氧化损伤,生物体可以激活一系列防御反应,如启动溶酶体降解途径。氧化应激过程中产生的ROS 可通过各种机制诱导自噬,通过自噬可以去除线粒体、内质网成分、过氧化物体和因氧化应激而受损的蛋白质,从而延缓细胞死亡[27]。Redd1是一种常见的应激反应元件,可以参与多种疾病的进展,抑制Redd1 通过抑制氧化应激和mTOR-自噬途径来拮抗OGD/R 诱导的神经元损伤对OGD/R 诱导的神经元损伤具有神经保护作用,这提示了对包括缺血性中风在内的缺血性脑疾病的治疗干预[28]。氧化应激和自噬之间有着密切的关系。
3.3 细胞凋亡与自噬
细胞凋亡是细胞基因程序激活的程序性死亡,是维持组织、器官大小和功能所必需的高度调节和关键的动态平衡机制[29]。细胞凋亡通路可分为两种主要的信号级联,分别是外源性和内源性。外源性是由特定的配体结合到跨膜死亡受体,激活半胱氨酸依赖的天冬氨酸定向蛋白酶(Caspases)形成了一个死亡诱导信号复合物(DISC),随后激活执行者Caspase,实现凋亡细胞死亡。内源性称为线粒体凋亡,是由氧化或缺氧应激、DNA损伤、生长因子剥夺等多种细胞应激引起的,进而导致线粒体膜电位(MMP)丧失和细胞色素C释放到胞质中。细胞色素C与凋亡蛋白酶活化因子1形成凋亡小体,激活Caspase-9,Caspase-9 进一步激活Caspase-3 诱导细胞凋亡。这一过程是通过释放溶酶体蛋白酶到细胞质中介导的,这可归因于溶酶体膜的渗漏或溶酶体膜完整性的丧失。因此,细胞凋亡与自噬呈负相关关系[30]。自噬和凋亡是细胞命运调节的两种基本病理和生理机制。且通过信号通路相互串联,一方面PINK1 和Beclin1 之间的相互作用负责启动自噬小体的形成;另一方面,自噬与细胞凋亡的相互作用涉及强烈依赖于Ca2+的信号转导通路[31]。最新的一项研究表明,proBDNF 参与了PSD 的神经可塑性。proBDNF 与p75NTR 受体结合后RhoA/JNK 信号通路被激活,随后凋亡相关蛋白(PSD95、synaptophysin 和P-cofilin)的表达促进了PSD 的进展。其机制可能与proBDNF 促进细胞凋亡和抑制神经突触再生有关。proBDNF 与p75NTR 受体结合,激活RhoA/JNK 信号通路,促进或抑制抗凋亡相关蛋白的表达。突触后膜中最重要、最丰富的支架蛋白是突触后密度蛋白95。是突触后膜的主要受体,主要存在于成熟的兴奋性谷氨酸突触中,在突触可塑性中起着重要作用。研究显示PSD 中PSD95 的表达降低,重组p75NTR 和siJNK 预处理逆转了PSD95的表达[32]。
3.4 PSD中自噬的相关信号通路
3.4.1 BDNF/TrkB信号通路
BDNF/TrkB 信号通路是PSD 中较为典型的信号通路,由具有高亲和力的TrkB 受体介导。脑源性神经营养因子(BDNF)是一种神经营养因子,广泛存在于中枢神经系统,在脑神经元的存活、发育、分化、增殖和损伤修复中起重要作用。一旦BDNF与TrkB结合,配体介导的复合体在细胞表面发生二聚化,导致下游丝裂原激活蛋白激酶(MAPK)、磷脂酰肌醇-3-激酶(PI3K)和磷脂酶CC(PLCC)通路的激活。TrkB 的这种激活也有助于神经发生、胶质形成、轴突生长和提高神经元存活率[33]。而在PSD 的发展进程中,BDNF/TrkB 信号通路与自噬密切相关,BDNF 通过改善自噬促进神经元存活,海马区BDNF/TrkB 通路通过增强海马区自噬,从而介导了抗抑郁作用的发挥[34]。此外有研究发现通过诱导激活BDNF/TrkB通路来抑制促炎细胞因子和ROS的产生,以进一步促进BDNF/TrkB 的抗抑郁作用。炎症细胞因子的产生和氧化应激也可通过BDNF/TrkB/MEK/ERK 途径和BDNFTrkB/p38 MAPK 途径进行调节从而减轻炎症以达到抗抑郁的作用[33]。
3.4.2 mTOR信号通路
自噬在阻止异常蛋白的积累,清除细胞中受损的细胞器维持细胞内稳态,细胞内稳态的维持依赖于细胞成分的合成和降解之间的严格调控,西罗莫司复合体1(MTORC1)是细胞营养感知中心的机械靶点,同时也是合成代谢和分解代谢过程的交汇点,它将环境信号与代谢过程联系起来,以维持细胞的自稳。mTOR信号通路是自噬的至关重要的调节剂,依赖于负反馈调节机制激活溶酶体促使自噬的发生,在机体营养充裕的条件下,mTOR 刺激生物合成途径促进细胞生长,并抑制自噬降低细胞代谢。能够感知细胞外和细胞内营养物质的波动,以调节细胞的生长、代谢和存活。然而,在应激条件下(包括氨基酸剥夺),mTORC1的失活刺激含有ULK1 的自噬前复合体的形成,这最终导致自噬小体的形成,促进自噬的发生[35]。有研究发现神经营养因子可能上调哺乳动物西罗莫司(mTOR)信号通路靶点。mTOR 信号的激活参与了脑出血的病理、生理过程,西罗莫司诱导mTOR 失活可降低脑出血中IL-1β、IL-6、TNF-α 和Caspase-3 的水平,神经营养因子通过抑制mTOR 信号通路促进细胞自噬,减少细胞凋亡和炎症反应[36]。
4 中药干预细胞自噬治疗PSD的作用
随着对PSD 的研究发现,中药对于延缓PSD 进展及改善患者生活方面具有较好作用,中药复方具有多靶点、多成分及多效应的优势,通过中药干预自噬治疗PSD 成为新的研究方向,现将中药复方、单体或成分、制剂通过自噬靶向治疗PSD作一系统总结。
4.1 中药单体及成分
近年来,多项研究表明众多单体中药成分如葛根乙醇提取物、柴胡皂苷A、反式白藜芦醇、石杉碱甲(石杉)、巴戟天寡糖、芍药苷等具有良好的抗抑郁作用,其主要目标靶点集中在TNF-α、IL-1β、IL-6、IL-10、ERK、CRE、BDNF、p-CREB、cAMP、p-CREB、Neun 等主要通过促进自噬的表达以达到抗抑郁的作用。见表1。
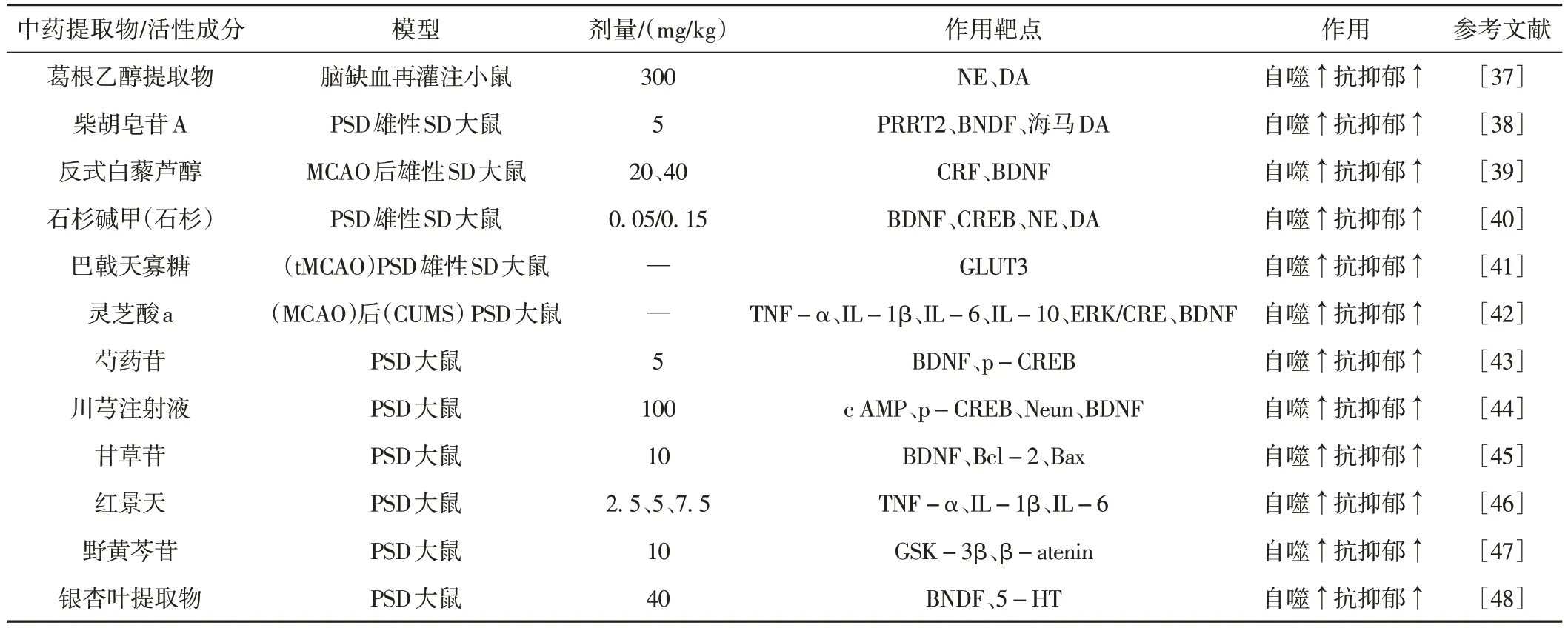
表1 中药提取物/活性成分通过调控自噬抗抑郁
4.2 中药复方及制剂
袁敏皎等[49]通过中国知网、中国生物医学文献数据库、维普等数据库收集了关于中医药治疗卒中后抑郁的相关文献经中医传承辅助平台总结归纳分析后发现,治疗PSD的中药药味以苦、辛为主,药性以温、寒、平为多;药物归经多归于肝、心、脾;多使用疏肝健脾类中药,常配伍理气、养心安神之品。多种药物协同作用、增效减毒,以达到更好的治疗作用,具体成方及机制见表2。
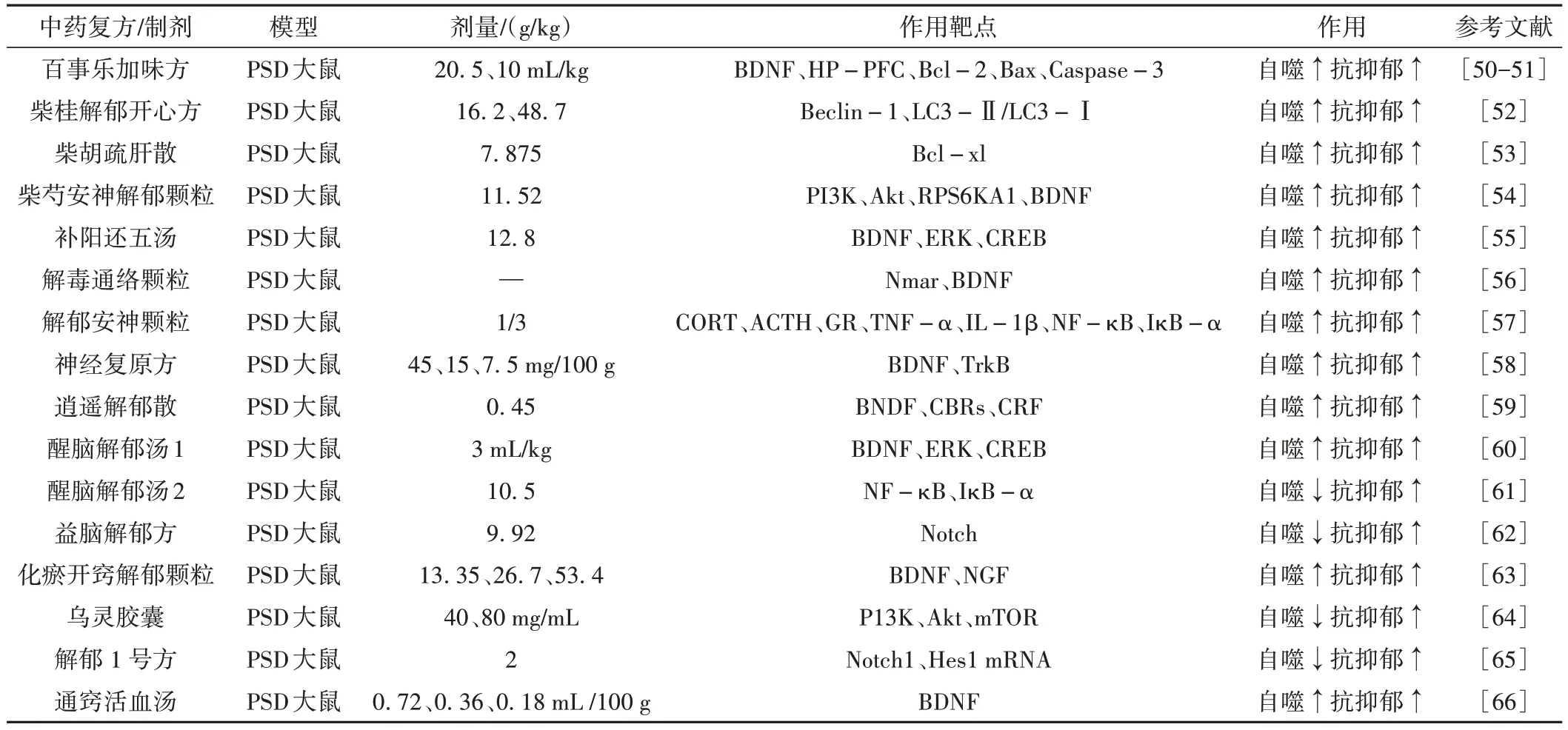
表2 中药复方及制剂通过自噬抗抑郁
5 结语与展望
综上所述,自噬通过利用自身细胞降解和重吸收功能维持细胞内环境的稳定,而中医理论中的“阴阳自和”同样是通过自身的调节使得失衡的阴阳重新回归平衡。自噬在PSD的发生、发展中有非常重要的作用,适度的自噬可以维持脑内环境的稳定,过多的自噬会破坏内环境稳定。自噬通过参与调控炎症反应、细胞凋亡以及氧化应激来达到抗抑郁的作用,其相关通路有BDNF/TrkB 和mTOR 信号通路,是目前PSD 靶点治疗的研究热点,中医药在治疗PSD具有较好的优势,众多研究表明,中药单体或成分及成方制剂通过调控自噬能够有效改善PSD 症状,但对于中医药对自噬与PSD 的相关性研究较少,对于其作用机制的研究主要集中于ERK、CRE、BDNF、mTOR 等相关靶点上,通路主要有BDNF/TrkB 和mTOR 信号通路,更多的信号通路及作用靶点有待进一步地研究,从自噬及阴阳理论的相似性出发,运用中医药多成分、多靶点的优势,深入探寻中医药通过微观自噬抗抑郁的作用,为新药研究及临床治疗思路提供新的方向。
