不同放牧模式对祁连山高寒草甸植物群落特征的影响
2023-10-08于泽航董全民张春平刘文亭杨晓霞刘玉祯张振祥吕卫东杨增增
于泽航, 董全民*, 曹 铨, 俞 旸, 张春平, 刘文亭,杨晓霞, 刘玉祯, 张振祥, 冯 斌, 吕卫东, 杨增增
(1.青海大学畜牧兽医科学院, 青海 西宁 810016; 2.青海省高寒草地适应性管理重点实验室, 青海 西宁 810016;3.三江源区高寒草地生态教育部重点实验室, 青海 西宁 810016)
高寒草甸作为青藏高原广泛分布的草地类型,具有涵养水源、碳固持、生物多样性保护和缓减气候变化等多种生态功能[1-3],对维持区域经济发展和生态平衡具有重要作用[4-5]。上世纪末,随着全球气候变暖、高寒牧区人口激增、人为活动加剧和过度放牧,青藏高原高寒草甸出现不同程度的退化[6-7]。过度放牧是造成高寒草甸退化的重要因素之一,该方式破坏了植物组织和功能,减弱了植株再生能力,降低了植物生物量,加之牧民不予管理的态度,高寒草甸进一步退化[8-12]。
草甸植物群落的稳定性和生产力是植被对环境变化响应的指示器,能充分反映植被对于环境和人类干扰的变化特征[13]。近年来,我国学者针对放牧导致高寒草甸持续退化的问题,通过比较放牧强度、放牧制度及放牧季节等管理方式,探讨了放牧对高寒草甸植物群落特征的影响。王向涛等[14]基于放牧强度的研究发现,草甸地上总生物量、物种丰富度在围封样地中最高,而可食牧草的生物量和盖度在中度放牧中最高。刘玉祯等[15]关于放牧方式的研究表明,在基于中度强度放牧条件下,藏羊、牦牛1∶2混合放牧对草地净初级生产力和家畜采食量最高。不同放牧管理方式对植物群落物种产生的影响不同,而物种又是组成群落的基本单位,物种的变化直接影响植物群落的变化,不同的物种在植物群落生态恢复过程中并非同等重要,往往关键种的损失会导致生态系统更大甚至不可逆的改变,且关键种在生态系统中发挥功能的重要性远大于其它物种[16]。关键种的确定有移除法和对比法,这2种方法对关键种的确定存在诸多主观因素,限制性较强,其准确性也有待商榷。网络分析广泛应用于微生物群落中关键种的识别,Zhang等[17]利用网络分析评价不同围封年限对高寒草甸植物群落结构及稳定性的影响中发现,基于物种重要值构建的网络图最具代表性,为确定植物群落的关键种提供了新方法,本研究植物群落关键种计算也使用此方法。综上所述,系统评价植物群落特征应从不同方面分析,多方面的研究结果更有利于高寒草甸植物群落稳定和健康发展。
本研究在青海省祁连县野牛沟乡开展试验,通过设置不同的牦牛放牧模式(放牧、放牧+补饲、围封),系统性分析植物群落结构、关键种及功能群特征,旨在明确不同放牧模式对高寒草甸植物群落特征的影响,以期为祁连山冬季草场的科学合理利用提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于青海省海北藏族自治州祁连县野牛沟乡(38°39′N,99°19′E),平均海拔3 300 m左右,年均温—1.3℃左右,年降水约在420 mm,降雨多集中于5—9月,气候类型属于典型的高原大陆性气候。全年无绝对无霜期,多以冷暖区分,冷季平均气温—25.6℃,暖季平均气温13.4℃,年均日照时数约2 900 h。研究区草地类型为高寒草甸,土壤类型为高山草甸土[18],研究点的优势物种有西北针茅(Stipasareptana)、矮生嵩草(Carexalatauensis)、洽草(Koeleriamacrantha)、珠芽蓼(Bistortavivipara)、鸡冠茶(Sibbaldianthebifurca)、多茎委陵菜(Potentillamulticaulis)等。
1.2 研究方法
1.2.1样地设置 于2019年5月,通过对当地牧区进行实地走访和调查,选取地形一致且植被特征较均一的冬季草场试验样地,放牧时间为每年11月—次年5月,其余时间禁牧,以牦牛为研究家畜,牦牛体重为(110±5) kg,样地设置3种不同放牧模式:放牧(M1)、放牧+补饲(M2)以及围封(M3)。放牧(0.35 AU·hm-2)指冬季草场连续放牧,不进行补饲;放牧+补饲(0.38 AU·hm-2)指冬季草场连续放牧,同时每头牛每日补饲1.2 kg干草;围封地禁牧不利用。
1.2.2植物样品采集方法 植被群落特征的测定是在2022年8月牧草生长期进行,在不同放牧模式的样地中以样带的方式进行取样,每个处理设置3条相距100 m样带,每条样带等距设置3个取样点,采用50 cm×50 cm的样方进行,分种测定了各物种的盖度、高度(每个样方内记录5株,不足5株的按实际株数记录)、频度、密度,并对牧草分种齐地面进行刈割装入信封,根系采集利用内径5 cm的土钻采集0~10 cm土层土样,将根系清洗干净后装入信封,最后把地上植物和根系带回实验室在烘箱中65℃恒温烘至恒重。
1.3 数据处理与分析
物种重要值(Important value,IV)和多样性采用以下公式计算[19]:
物种重要值(IV):IV=(Hr+Br+Dr)/3
(1)
Patrick丰富度指数(R):R=S
(2)
Shannon多样性指数(H):H=-∑(VilnVi)
(3)
Pielou均匀度指数(D):D=H/lnS
(4)
式中:Hr为相对高度;Br为相对物种生物量;Dr为相对频度;S为物种数;Vi为物种i的重要值。采用单因素方差分析(One-way ANOVA)不同放牧模式下植物群落特征指标的变化。
其次,应用网络分析(Network analysis)确定植物群落关键种。关键种确定分为3个步骤[20]:计算每个物种的重要值(IV)[21];基于物种重要值计算物种相互间的Pearson相关系数r,用于构建网络图的边[22];把物种当作节点,将相关性系数r绝对值小于0.8的边且显著性水平大于0.05的值剔除。根据上述的节点和边,利用Gephi 0.9.2制作不同放牧模式下的无向随机网络图。在网络图中,特征向量中心度(Hub value)的大小能够代表物种(节点)在网络中的重要性[23],在网络图中节点最大的即为该模式下植被群落的关键种[24]。平均连通度(Average connectivity)来权衡物种间的竞争关系,数值越高说明该群落竞争越强、复杂性越高[25],平均聚类系数(Average clustering coefficient)来权衡植被群落的组织程度,数值越高说明该群落的组织程度越高[26]。
以上全部统计、绘图都在R 4.0.3中进行。
2 结果与分析
2.1 不同放牧模式对植物群落结构的影响
由图1可知,M2,M3模式下植物地上生物量和高度显著高于M1(P<0.05),但M2,M3之间无显著差异;M3模式下植物地下生物量和盖度显著高于M1,M2(P<0.05);M1,M2,M3模式下植物丰富度差异显著(P<0.05);M1,M2,M3模式下植物群落的Shannon-Wiener指数和Pielou均匀度指数无显著差异。
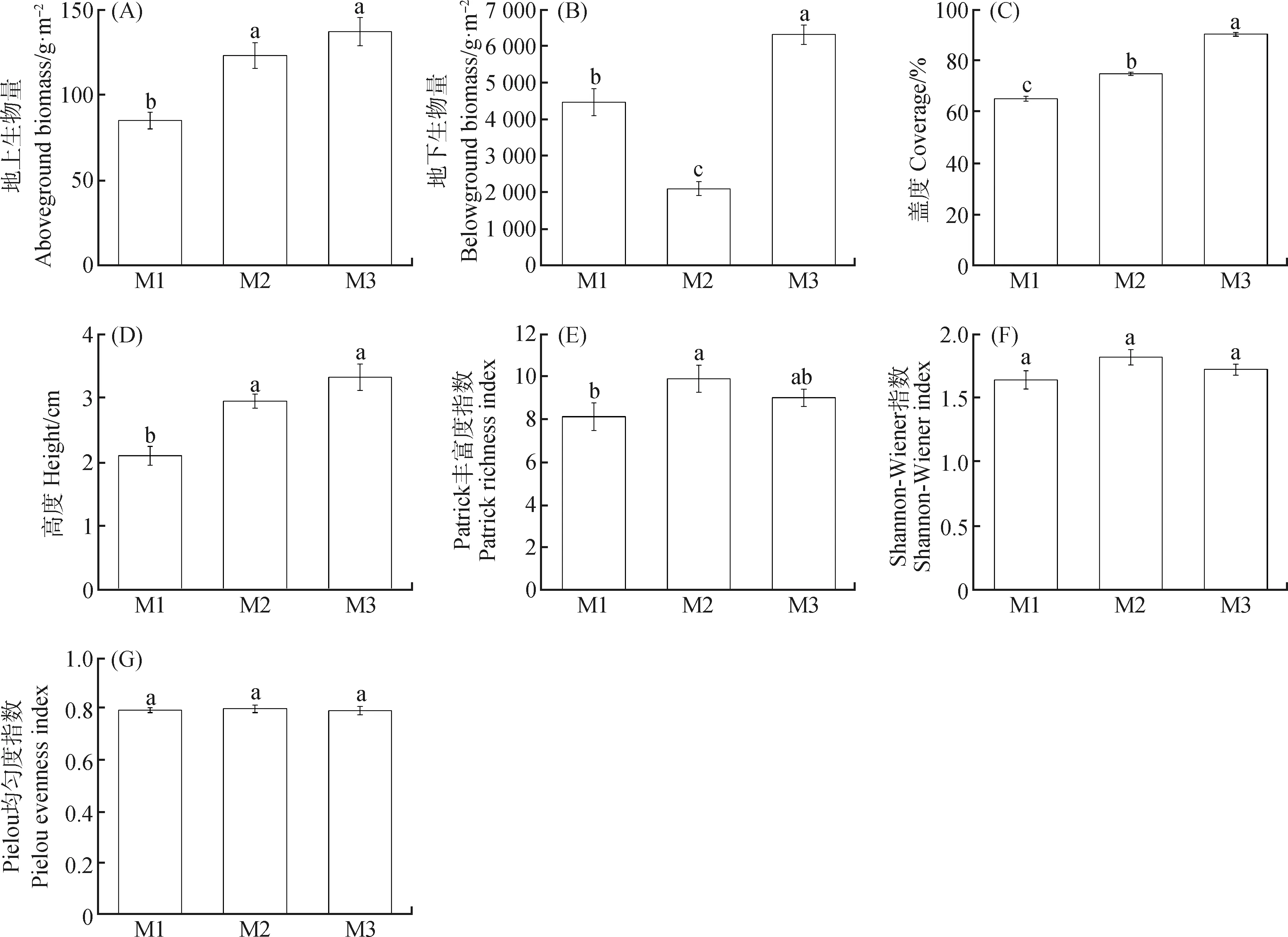
图1 不同放牧模式对植物群落结构的影响
2.2 不同放牧模式对群落网络结构及关键种的影响
由图2A可知,M1模式下的植物群落网络图中有13个节点26条边,植物群落关联性较差;M2模式下的植物群落网络图中有18个节点28条边,呈现出较好的植物群落关联性;M3的植物群落网络图中有16个节点23条边,植物群落关联性中等。M1,M2,M3放牧模式下关键种依次为鸡冠茶、洽草、多茎委陵菜(图2B)。
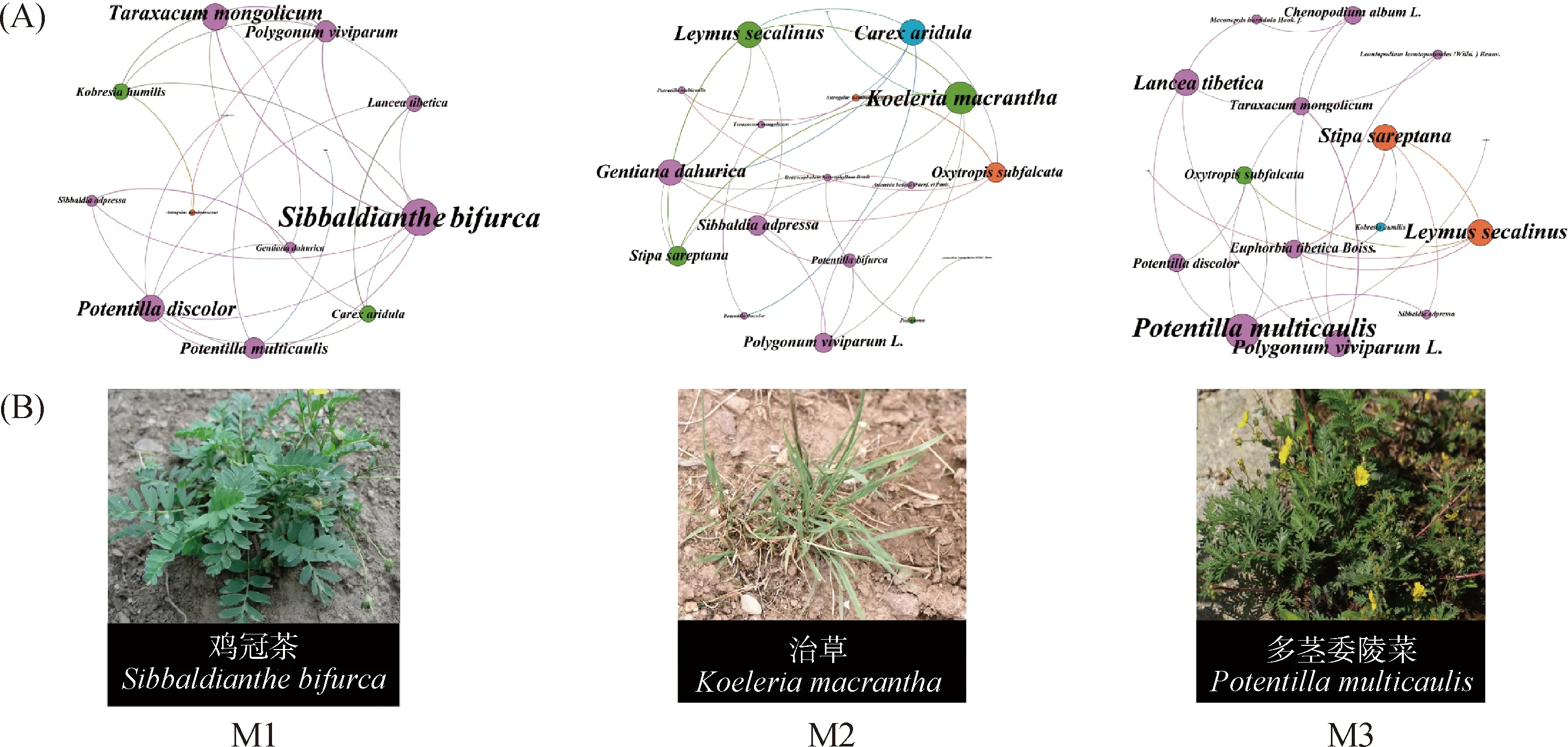
图2 不同放牧模式下的植被群落网络图
由图3可知,M2模式具有最高的平均连通度,表明群落物种间竞争平衡性、复杂度最高;M3的平均连通度次之,M1的平均连通度最差。不同放牧模式下的平均聚类系数排序为:M2>M1>M3,表明在放牧时添加补饲能够维持植被群落物种间竞争力和复杂度。

图3 不同放牧模式下平均连通度和平均聚类系数的变化
由图4可知,M1模式的拟合程度最高,群落间的连通度和Hub值较聚集,物种间的竞争强,复杂性高,因此,群落稳定性高,关键种更替速度慢;M2模式拟合度处于M1和M3之间,物种竞争和稳定性相对较平衡;M3模式的拟合程度最低,群落间的连通度和Hub值差异较分散,物种间竞争弱,复杂性低,使得群落稳定性低,从而关键种更替速度快。

图4 不同放牧模式下平均连通度与特征向量中心的关系
2.3 不同放牧模式对植物功能群特征的影响
M2和M3模式下禾本科的生物量高于M1模式,M1模式下莎草科的生物量高于M2,M3,而豆科和杂类草的生物量在3种模式下差异不大(图5A)。

图5 不同放牧模式下植物功能群特征
不同放牧模式下,各植物功能群占比不同(图5B)。禾本科功能群重要值呈M2>M3>M1的趋势变化,莎草科、豆科和杂类草功能群重要值呈M1>M3>M2的趋势变化。
3 讨论
3.1 不同放牧模式对植被群落结构的影响
植被群落结构是评价高寒草甸生态系统健康的重要指标,其变化过程与变幅显著影响高寒草甸生态系统功能与生物多样性[27]。本研究发现,不同放牧模式会显著改变植被的地上生物量、高度和盖度。连续放牧导致群落盖度、高度、生物量和物种数减少,且可食牧草占比减少,进一步降低了高寒草甸植物群落的稳定性,这和唐刘燕等[28]的研究结果相似。一方面由于家畜的放牧,使得植物光合组织和生长组织长期被家畜采食,牧草结构被反复损伤,导致牧草的生长速率变缓,受损伤的植被组织易遭受病菌侵染,增大了牧草的死亡率。此外,放牧还导致牧草的繁殖组织被转移,牧草繁殖受到影响,进一步延缓了牧草的繁殖期[29]。另一方面可能是资源分配权衡的作用[30],当严重放牧胁迫到植物时,其更倾向于避牧防御而不是非补偿生长,会把更多的资源分配到地下部分来促进根系的生长。从地下生物量角度分析,M1模式下,牦牛不间断的践踏和采食,使得植被补偿性生长持续进行,促进了根系的生长;M2模式下,植被采食减少,更多的资源向地上部分聚集;M3模式下由于不受家畜干扰,土壤紧实度下降,土壤养分逐步改善,也促进了地下生物量的积累,这一结果与张晓艳等[31]和赵景学等[32]的研究相似。植被群落物种的多样性和丰富度是群落的重要性质,其对不同的环境异常敏感,容易受外界环境影响发生改变[33]。本研究发现,三种模式下的物种丰富度有着显著的差异,而Shannon-Wiener指数和Pielou指数没有显著差异。在M2模式下,物种的丰富度最高。依据植物群落构成过程及过滤机制(Community assembiy and filtering mechanism),在生存环境空间相对较好的情况下,由于物种竞争时间漫长及存在短暂的排斥现象,加入适当放牧干扰,能刺激植株再生,促进植被补偿生长,使群落的物种数增多[34]。
3.2 不同放牧模式对关键种及稳定性的影响
关键种对支撑草地生态系统结构有着重要的作用,倘若关键种丢失,生态系统结构会发生巨大改变[35]。关键种的存在削弱了物种之间的竞争,进而维持了生态系统的稳定性。关键种在维持生态系统稳定性的同时,进一步影响其他物种的分布与数量,最终决定了植物群落的物种多样性、稳定性和其它一连贯的生态效益[36]。本研究中,M1和M3模式下的关键种都为杂类草,其耐牧性和耐干扰性相对较高,可以有效延缓高寒草甸退化[37],但由于M1模式下长时间的放牧导致了禾本科牧草减少,牧草质量下降,进而高寒草甸群落发生了改变。在M2模式下,由于补饲效应的存在,降低了牦牛对牧草的采食率,进而对草地植物群落结构产生一定的调节作用[38]。Newman等[39]研究发现,饥饿与饱腹状态下,不仅家畜的采食时间和采食率有差异,而且牧草中禾本科植物比例也有较大变化,这是由于不同状态下家畜的采食量不同。由于M2模式下存在补饲效应,相对减少了家畜对禾本科植物的采食,使得长势较高的禾本科植物获得更多的光能资源,在物种竞争中占据优势[40-41]。从植被群落的平均连通度和平均聚类系数分析,M2模式下植被群落物种竞争力和复杂度较高,这是由于在适当放牧和补饲下,物种的互作强度提高,这与Gao等[42]的研究较为一致。
3.3 不同放牧模式对植物功能群特征的影响
草甸植被群落中功能群生物量及占比可直接反映草地的分布格局,不同放牧模式下的地上生物量占比可作为判断草地状况、生产力和演替趋势的重要指标[43]。放牧过程中,家畜采食和践踏直接作用于草地,草甸土壤发生了变化,从而严重干扰植物的生长发育,导致植物群落的高度和盖度下降,生物量大幅度减少,其中禾本科牧草的生物量变化最为显著[44-45]。不同放牧模式下的禾本科植物功能群生物量及重要值发生显著变化,禾本科绝大多植物属于家畜喜食的牧草,放牧过程中,其被家畜大量采食,使得禾本科牧草生物量大幅降低,这与张娜等[46]的研究结果一致。M1模式下的莎草科功能群占比最大,这是由于家畜优先采食上层优质牧草后,从而使得作为下层植物的莎草科牧草获得了较多的光热资源,使其在群落功能群占比提高[47-48];M2模式由于存在补饲效应,家畜采食率降低,禾本科植物高大直立的特征使其获得更多的光合优势,冠层低矮的莎草科植物得不到更好的生长,所以在该模式下占比最低。在M1模式下豆科和杂类草植物重要值最高,因为适度放牧刺激植物产生补偿生长,未被采食的植物光合效率增大,增强了植物的适应能力,使得在群落中占比最高。M3模式下植物功能群组成发生改变,禾本科和莎草科植物增多。原因是消除了家畜对植物的采食和践踏作用,禾本科植物和莎草科植物因其形态特征高大,较其它植物拥有更充足的环境优势,从而生长较快,在群落中占据主要地位,这与周华坤等[49]研究相似。
4 结论
通过对高寒草甸冬季牧场的不同利用发现,草甸植物群落的物种组成和结构发生了变化,但对植物群落的Shannon-Wiener指数和Pielou均匀度指数无显著影响。“放牧”“放牧+补饲”“围封”的关键种分别为鸡冠茶、洽草和多茎委陵菜。从植物功能群生物量分配角度来看,“放牧+补饲”下植物群落生物量分配更为合理,能有效增强群落的稳定性。因此,高寒草甸冬季牧场利用中,建议采用“放牧+补饲”方式,从而降低物种之间的竞争,使植物具有更好的生存空间,更有利于维持冬季牧场草地群落稳定性,能对高寒草甸生态系统的健康发展产生积极影响。
