Regeneration of Nothofagus dombeyi (Mirb.) Ørst.in little to moderately disturbed southern beech forests in the Andes of Patagonia (Argentina)
2023-10-07StfnZrStfniStorzGorgLitingrNtliZoJolsonJosBvStffiHinrihsChristophLushnrGrilLogurioAloisSimonMrUrrtvizyHlgWlntowski
Stfn Zr, Stfni T.Storz,, Gorg Litingr, Ntli Zo Jolson, José Bv,Stffi Hinrihs, Christoph Lushnr, Gril Logurio, Alois Simon,f,Mrí F.Urrtvizy, Hlg Wlntowski,*
a Faculty of Agricultural, Environmental and Food Sciences, Free University of Bozen-Bolzano, Bolzano, Italy
b Department of Ecology, University of Innsbruck, Innsbruck, Austria
c Faculty of Resource Management, HAWK University of Applied Sciences and Arts, G¨ottingen, Germany
d Department of Plant Ecology and Ecosystem Research, Albrecht-von-Haller-Institute for Plant Sciences, University of G¨ottingen, G¨ottingen, Germany
e Centro de Investigaci′on y Extensi′on Forestal Andino Patag′onico (CIEFAP), Esquel, Argentina
f Department of Forest Planning, Tyrol Forest Administration, Office of the Tyrolean Government, Innsbruck, Austria
Keywords:
ABSTRACT Natural forests and stands subjected to little to moderate human impact are continuously declining worldwide and with these,their biodiversity and ecosystem services.Many Nothofagus forests in the south of the South American continent are in a pristine state or only moderately impacted by humans.Forest grazing by livestock, in the past and still today often practiced in a non-sustainable way is,however,increasingly under discussion to meet current environmental and socio-economic challenges.Accordingly, we investigate the regeneration of Nothofagus dombeyi, a keystone species of the Patagonian Andes, in secondary forests in Argentinian northern Patagonia,particularly addressing the role of disturbance through grazing by livestock at various intensities.We test the hypothesis that the regeneration of this tree species is favored by grazing impact and,thus,disturbance of the herb layer and soil surface.In support of our hypothesis, Nothofagus dombeyi regeneration was significantly higher in terms of individuals and height classes in moderately grazed forests.Multivariate analysis shows significant positive effects of moderate grazing pressure,herb layer cover,and the occurrence of bare soil on the regeneration of Nothofagus dombeyi.Our results show that an integration of livestock grazing and forest regeneration is possible and that agroforestry systems can be an adequate management option for stakeholders in the region.A grazing management can also be part of a forest fire prevention strategy.However, the regeneration success and grazing pressure should be continuously monitored.
1.Introduction
Worldwide, forest ecosystems are threatened by increasing deforestation,land-use intensification,and climate change(Keenan et al.,2015;FAO and UNEP,2020).The loss of forests and the unsustainable use and management of forest ecosystems result in a decline of biodiversity and ecosystem services such as the production of timber and non-timber forest products, erosion and flood control, carbon sequestration and storage,climate regulation,and water purification(Breshears and Allen,2002; Chiabai et al., 2011; Shimamoto et al., 2018; IPBES, 2019; IPCC,2019).Particularly near-natural old-growth forests with old trees and complex structures provide diverse ecosystem services which young or intensively managed forest plantations cannot offer (Felipe-Lucia et al.,2018;De Assis Barros et al.,2022;Gilhen-Baker et al.,2022).Additionally, there is evidence that old-growth and near-natural forests may be better adapted to climate warming than managed forests(Mausolf et al.,2018;Colangelo et al.,2021).
Forest regeneration is crucial for the long-term provision of ecosystem services, be it natural or supported by silvicultural measures such as thinning and tree planting.Near-natural forest management and forest restoration aim at promoting self-regenerating,site-adapted trees(Zerbe,2002; Chazdon and Guariguata, 2016) which offer not only environmental but also socio-economic benefits (Di Sacco et al., 2021).For the successful establishment of trees and the sustainable management of rejuvenating forests, knowledge on the ecology of tree regeneration is essential(Tinya et al.,2020).
The forests built-up by various species of the genus Nothofagus in Patagonia are among those temperate forests on the global scale which are still largely contiguous and in large parts not managed or influenced by strong human impact, although strongly affected by anthropogenic wildfires in the colonization period about 100 years ago (Willis, 1914;Veblen et al.,1999).However,they are subject to natural disturbances in different intensities, with wind and snow avalanches being the most important disturbance agents (Amoroso and Blazina, 2020).One of the species which naturally builds monospecific or mixed stands (particularly with Austrocedrus chilensis (D.Don) Pic.Serm.& Bizzarri) in Patagonia is Nothofagus dombeyi(Mirb.)Ørst.,an evergreen monoecious tree species,which belongs to the family Nothofagaceae(Puntieri et al.,2000;Gut,2008).It is found in forests from 36°30′to 47°00′S in Chile and from 38°40′to 44°00′S in Argentina(Veblen et al.,1996;Donoso et al.,2022)and occurs on a wide range of site conditions and thus, is an important component of the forest vegetation of the Patagonian Andes(Ward,1965;Seibert, 1972; Dezzotti, 1996; Eskuche, 1999; Donoso et al., 2022).It thrives best on moist sites and prefers south-facing slopes at low and medium altitudes of the Andes as well as floodplains(Veblen et al.,1996;Gut, 2008).In higher altitudes, Nothofagus pumilio (Poepp.& Endl.)Krasser becomes dominant and builds up the timber line.In places,N.antarctica (G.Forst.) Oerst.forms the timber line, also acting as a pioneer species resprouting after fires and being able to replace N.pumilio(Armesto et al.,1992;Mermoz et al.,2005;Kitzberger et al.,2016).
However, these natural and near-natural forests of Nothofagus dombeyi are increasingly damaged and declining due to climate change,fire,and land-use intensification (Giaché and Bianchi, 2018).Bran et al.(2001)report massive mortality events that have occurred in response to extreme droughts such as in the period of 1998–1999 in northern Patagonia (see also, Suarez et al., 2004; Suarez and Kitzberger, 2008;Molowny-Horas et al., 2017).Against the background of the ecological key role of Nothofagus dombeyi in the Patagonian forests and the necessity of sustainable management and restoration of still functional as well as degraded southern beech forests, we aim at an investigation of the tree regeneration of N.dombeyi mainly in response to grazing intensity that,besides fire,represents the most frequent and widespread anthropogenic disturbance agent in Patagonian forests (Veblen et al., 1992), but often also the livelihood for local stakeholders.
Forest grazing has a long history in temperate forests of Europe and Asia(Rotherham,2013)and was introduced to North and South America,Australia,and New Zealand a few centuries ago(Borman,2005).With a global review, ¨Ollerer et al.(2019)address the impact of domestic livestock grazing on temperate forest vegetation against the background that livestock is considered as competitor to timber production(e.g.,Graham et al.,2010)and thus,livestock grazing is seen as an undesirable practice that should be completely taken out of the forests (e.g., Bernes et al.,2018).However, the removal of livestock from forests has led to the disappearance of complex and specific disturbance patterns which created diverse habitats subject to nature conservation (Cooper and McCann, 2011; Zerbe, 2022).Accordingly, ¨Ollerer et al.(2019, p.14)recommend to re-visit the question of “what we expect from the forest and from grazing”and conclude that forest grazing can contribute to the well-being of local communities involved in animal husbandry,without greatly affecting timber production and, therefore, providing higher income from the same area of land.Given that vast areas of Patagonian forests are and will be in the future increasingly damaged by fires(Mundo et al., 2017; Kitzberger et al., 2022), forest grazing might,additionally, be a management option to reduce the fire risk (Lovreglio et al.,2014; Varela et al.,2018).
Our study motivation is to deepen our understanding of the regeneration biology of Nothofagus dombeyi which has been addressed earlier under different aspects, e.g., the establishment success of seedlings(Veblen et al.,1981,1996),the influence of disturbance on regeneration(Veblen,1989;Veblen et al.,1996;Larraín,1997),and the effects of light regimes on regeneration patterns (Encina et al., 2019).There is a knowledge gap on how grazing affects the regeneration of N.dombeyi.One of the few related studies is that of Blackhall et al.(2008) who examined the effects of livestock on the post-fire recovery of a mixed
Nothofagus dombeyi-Austrocedrus chilensis forest in the Nahuel Huapi National Park in Argentina.With regard to introduced livestock on post-fire recovery of Nothofagus forests, they recommend fencing for periods long enough to allow the dominant tree species to grow to heights at which they are no longer severely inhibited by cattle browsing.By focusing on tree regeneration in the seedling and sapling state within later successional forest stages, our hypothesis is, however, that N.dombeyi regeneration is favored by moderate grazing impact.From our findings,we derive recommendations for the management of N.dombeyi forests.
2.Materials and methods
2.1.Study area
The study area is located in the Province Río Negro in Argentinian Patagonia at the southern limit of the Nahuel Huapi National Park.We chose the El Manso Valley and the Valley of Río Villegas as the two core areas of our study (Fig.1).The climate in the Province Río Negro is characterized by a pronounced climatic gradient (Barros et al., 1983;Paruelo et al., 1998; Bianchi and Ariztegui, 2012), with most precipitation falling in the winter season (Gallopín, 1978).The highest mean annual temperatures (MAT) are found in the northern parts of the province (MAT >15°C), the lowest MAT values at higher elevations in the west of the study region(<10°C).Mean annual precipitation(MAP)sharply declines from West to East, i.e.from the Cordillera to the Patagonian steppe; according to Lenaerts et al.(2014), this gradient ranges from about 5,000 mm in the west to less than 500 mm in the east.The steppe is characterized by semi-arid conditions with MAP ranging from 200 to 700 mm(Karger et al.,2017).
For El Manso,Estancia Piedra Pintada(Oscar Lanfré;latitude:41°36′S and longitude: 71°35′W, altitude: 400 m a.s.l.), a long-term average(period 1980–1995 and 2002–2004) of an annual mean temperature(BIO1)of 9.3°C and a mean annual precipitation(BIO12)of 1,225 mm are reported (INTA, 2015; Table S1 in Supplementary Material).In this valley,the precipitation of the wettest quarter(BIO16)from May to July is 593 mm,that of the driest(BIO 17)from January to March is 124 mm.The precipitation seasonality(coefficient of variation; BIO15) is 60%.
The soils are mainly young Andosols of volcanic origin(IUSS Working Group WRB,2015;Buduba et al.,2020)and are,in general,characterized by a high cation exchange capacity, low to moderate base saturation(unpublished,field survey),a high-water storage capacity,and naturally moderate nitrogen contents (Kühn et al., 2021).One of the strongest limitations for plant growth on allophanic andosols is the low phosphorus availability (Matsuoka-Uno et al., 2022).Keystone tree species counteract the limitation by forming ectomycorrhizal (EcM) associations(Barroetave~na et al., 2019) or physiological adaptations for improved P resorption (Hayes et al., 2018).Barroetave~na et al.(2019) found the highest number of unique EcM species for Nothofagus dombeyi,followed by N.pumilio and N.antarctica.
The pronounced climatic gradient from west to east, particularly regarding mean annual precipitation, is reflected by a marked forest vegetation change from dense,productive rain forests on the Chilean side of the Andes(Valdivian temperate rain forests;e.g.,Donoso et al.,2020)to productive pure and less-mixed Nothofagus forests in the west of the study region in the Argentinian Andes (Donoso et al., 2022), and open xeric forests,patchy forest vegetation,and finally shrubland towards the east,which gives way to the steppe vegetation in the lowlands east of the Andes(Veblen et al.,2003;Kreps et al.,2012).
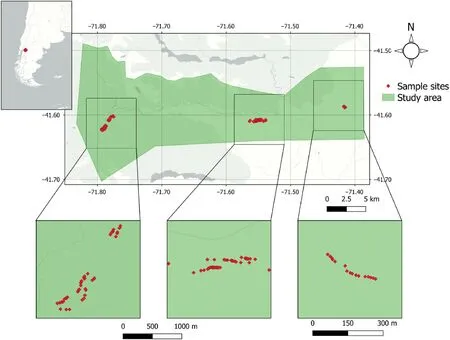
Fig.1.Study area in Argentinian Patagonia and location of transects with sample sites in the El Manso Valley and the Valley of Río Villegas at 41°58′49′′ S to 41°60′26′′ S and 71°41′54′′ W to 71.78′09′′ W.
The forests in the mountains of Rio Negro Province have been classified by Hueck and Seibert (1981) as “mainly deciduous forest of the temperate region,with Nothofagus pumilio and Nothofagus antarctica”and“forest of Austrocedrus chilensis”.In our study area,however,a mosaic of mostly pure N.dombeyi or mixed Nothofagus dombeyi-Austrocedrus chilensis stands are found, sometimes also pure Austrocedrus chilensis forest patches.The tree species Lomatia hirsuta Diels ex J.F.Macbr.,Embothrium coccineum J.R.Forst.& G.Forst., Luma apiculata (DC.) Burret, Maytenus boaria Molina,and Nothofagus antarctica(G.Forst.)Oerst.can also be part of the overstory, but in intermediate or suppressed positions.In mixed southern beech forests near to our study area(Los Alerces National Park,Loma del Medio-Río Azul Forest Reserve,Río Manso Valley),Caselli et al.(2021b)recorded up to 2,198 trees per ha,basal areas between 48.6 and 76.3 m2per ha, volumes between 436 and 977 m3per ha, and for N.dombeyi,tree heights between 20 and 30 m and a diameter generally exceeding 40 cm,with a maximum of 116 cm.
Human use of these forests has been very heterogenous both,in time and space.There is archaeological evidence of the presence of hunters and gatherers in the valleys already from 8200 BP onwards(Borrero and Barberena,2006;Caruso-Fermé,2021).Since 1700 BP,human impact on forests increased considerably with the expansion of settlements and population growth in the valleys.Larger forest clearings started in the 19th century with the arrival of European settlers (Giaché and Bianchi,2018).Today, forest grazing is performed at various levels of intensity,reflected by browsing and trampling damage or the presence of even bare soil as well the occurrence of exotic herb and grass species (Quintéros et al., 2017).Additionally, non-native tree species have been planted such as Pseudotsuga menziesii which has become invasive at places (Salgado Salomón et al.,2013).On the other hand,due to the inaccessibility of steep and remote slopes, some southern beech forests still show no visible human impact.Yet, fires are increasingly reported in our study area.Some of them have natural causes; however, the majority are man-made (Veblen et al., 1999) and can, due to strong winds, destroy large forest tracts within short time.
2.2.Methodological approach
Our study is based on the current knowledge of ecologically important traits of N.dombeyi.Hereby, we differentiate the ecological characteristics of this tree species in the individual development stages dispersal, germination, and seedling development (early development stages),on the one hand,and mature and old-growth stage,on the other(late development stages) (Table 1).N.dombeyi is an evergreen tree species(Veblen et al.,1996)and is characterized as an early-(Reyes-Díaz et al.,2005)to late-successional species(Veblen et al.,1981;Montenegro et al.,1997).Seed dispersal distance and velocity as well as viability are low.The seeds get mainly dispersed by gravity, 95% of the seeds occur within a 20-m distance to the mother tree.Frost and drought sensitivity are rather high, and the shade tolerance of seedlings is low to intermediate.The maximum age is recorded with 600 years, maximum growth height with 50 m, and maximum diameter at breast height with 4 m.Mature trees have a high sensitivity to late frost and fire, can be easily damaged by wind breakage and have a low capacity of re-sprouting.
For better understanding regeneration, we performed field work in the two valleys in Northern Patagonia.Our sample plots were laid out systematically along 10 elevational transects in the El Manso(9 transects)and the Río Villegas Valley (1; Fig.1).Starting points of the transects were the valley bottoms at around 450 m above sea level, from where transects with a width of 2.5 m were placed up to 1,000 m a.s.l.We established plots of 20 m × 20 m wherever one or more N.dombeyi seedlings or saplings (definition c.f.next paragraph) were found inside the transect,with the seedling or sapling as the plot center.Accordingly,a total transect area of around 25,000 m2was covered for our treeregeneration investigation.

Table 1 Ecological characteristics of Nothofagus dombeyi based on available data, with references given.

Table 1 (continued)
From the plot center,1)a regeneration(micro-)plot with a size of 5 m× 5 m, and 2) a forest structure (macro-)plot of 20 m × 20 m were demarcated.A total of 106 plots were sampled in February 2022.On the macro-plot,we recorded slope aspect and inclination(measured with an inclinometer), elevation above sea level (hand-held GPS), total vegetation cover of the herb,shrub(1–5 m height growth),and tree layer(>5 m;visual estimation of cover in percentage)and mean height of the herb and shrub layer(using a measuring tape in cm),total moss layer cover(in percent),the percentage of bare soil,and lying and standing dead wood(visual estimation of percent coverage), and the depth of the organic humus layer(in cm).Additionally,the forest stand was characterized by determining the mean upper height of the canopy and the tree species present with a diameter at breast height of >7 cm (Hui et al., 2019).Logging intensity was estimated by the number of tree stumps and differentiated into low(no stumps), medium (1–2), and high(>2).
On the micro-plot, we sampled the regeneration of all tree species,including N.dombeyi, counting all seedlings and saplings.Following Mirschel et al.(2011),the height-growth classes of tree individuals were differentiated in five classes, i.e.I, seedlings <10 cm; II, saplings of 10–50 cm; III, 50–100 cm; IV, 100–150 cm; and V ≥150 cm up to a diameter at breast height(DBH)of 7 cm.We consider grazing damage of regenerating trees mainly due to free-ranging cattle as a proxy for grazing intensity.Grazing damage of tree individuals was categorized with the browsing index after Gadola and Stierlin(1978)into three classes,with i= low (main shoot not damaged, lateral branches with slight grazing damage), ii = moderate (main shoot damaged and lateral shoots also damaged),and iii=high(main shoot damaged,“bonsai aspect”,or total loss of leaves).These values were summed up per tree individuals and weighed over the number of individuals to obtain the total grazing pressure per plot following Mirschel et al.(2011).Additionally, we measured the distance to the next diaspore source of N.dombeyi,i.e.the next mature tree.
2.3.Statistical analysis
For analyzing the general characteristics of the stand structure and regeneration plots,we applied descriptive statistics.To identify decisive variables for the regeneration of N.dombeyi, an ordinary least-square regression (OLS) based on 106 observations was computed, testing for the effect of various biotic and abiotic factors on the abundance of N.dombeyi regeneration as dependent variable.To do so,only the height classes I to III were examined,which represent the regeneration<100 cm height.The response variable was tested for normal distribution with a Shapiro-Wilk test after arcsine transformation with the R package best-Normalize(Peterson,2021)to approximate normal distribution.Firstly,we tested for non-linear dependencies, independency and normal distribution of error values, no multi-collinearity and strict exogeneity(Gauss-Markov assumptions).Secondly, the standard F test was used to check if the obtained regression model was significant and the significance of the regression coefficients was checked by the results of the model's internal t-test.The goodness of fit was evaluated by the adjusted R2.Using R2,the effect size f2was calculated,following Cohen(1992).
For the regression analysis, also the sum of individuals of all other tree species found in the regeneration layers,except N.dombeyi,in height classes I-III (≤100 cm) and IV-V (≥100 cm and <7 cm DBH) were calculated and included as independent variables.These variables are used as a proxy for interactions of N.dombeyi with other tree species through competition or facilitation.The statistical analyses were performed with R statistical software(R Development Core Team,2022).
Our sample plots mostly were located on north-to northwest-exposed slopes which range from 0% to 84% with a mean of 19.12% (Fig.2),which implies more exposure to direct solar radiation and higher temperatures during the summer compared to flat locations.Accordingly,the variable slope aspect was used as a matrix of dummy (dichotomy) variables for slope aspect North(315°-360°;0°–45°),East(45°–135°),South(135°–225°), and West (225°–315°).The integration of elevation as an independent variable into the regression models accounts for changes in temperature and precipitation across the regeneration transects.
3.Results
3.1.Structural characteristic of the investigated forest stands
The forest stands which were sampled in the macro-plots were mainly built up by N.dombeyi,mixed with Austrocedrus chilensis.To a much lesser extent, the tree species Luma apiculata (DC.) Burret, Lomatia hirsuta,Nothofagus antarctica, Azara microphylla Hook.f., Schinus patagonicus(Phil.) I.M.Johnst., Aristotelia chilensis (Molina) Stuntz, and Embothrium coccineum J.R.Forst.& G.Forst.were found in the tree layer which ranged from about 16 to 24 m height.The cover of the tree layer ranged from 10% to 70% (mean about 34%).The cover of the shrub layer was recorded from 0 to 70% with a mean height of 270 cm.Bare soil was abundant in most of the plots with a minimum cover of 5% and a maximum cover of 60%.The amount of lying dead wood was higher(with a mean cover of 17%), than the amount of standing dead wood(with a mean cover of almost 3.5%).The mean depth of the organic humus layer was around 2.7 cm.Since mature trees of N.dombeyi were present in most of the sample plots(on 104 out of 106),a mother tree as seed source and for regeneration was mostly present.Thirty-eight plots were assessed with low grazing intensity,33 with moderate,and 35 with high.
3.2.Regeneration of Nothofagus dombeyi and other species
On our 106 sample plots along the studied transects(5.3%of the total transect area),a total of 3,471 tree individuals(about 13,100 per ha)in the regeneration layers were found which belonged to 11 tree species(Fig.3).40% (n = 1386) of the total individual number belonged to N.dombeyi.Aristotelia chilensis was recorded with the second highest number of regenerating individuals(24%,n=839),followed by Schinus patagonicus(15%,n=507).One individual of Pseudotsuga menziesii was found in the height class III.It was recorded in an area close to plantations and high logging activity.Most saplings of N.dombeyi(about 30%)were found in class III, the lowest number occurred in class I (about 13%), and the classes I-III (used for the regression analysis) with altogether 955 individuals accounted for 68% of all seedlings/saplings.Schinus patagonicus follows this trend of the distribution, with more individuals in classes III and IV,and less in classes I,II,and V.However,the individuals of Aristotelia chilensis in the regeneration layer increase with height classes from 122(class I)to 245(class V).
Table 2 summarizes the descriptive statistics of the potential impact factors on the regeneration of N.dombeyi showing the mean,maximum,minimum and the standard deviation.
Regeneration of N.dombeyi was found in our sample plots with a mean of about 13 individuals per plot which corresponds to a mean of 5,200 per ha(Table 3).Since we sampled only plots in which N.dombeyi was present in the herb and/or shrub layer,we get a minimum value of 1 individual of N.dombeyi per plot.
The most influential variables(independent variables,cf.Table 2)on N.dombeyi regeneration (height classes I-III pooled) were selected by stepwise OLS regression.In the final model, four independent variables were included at a 5% significance level (Table 4), leading to a highly significant model (p < 0.001) with a determination coefficient of 0.273(adjusted R2).The decisive variables (in descending magnitude of influence, i.e.standardized coefficient Beta) for the regeneration of N.dombeyi were 1) grazing intensity (moderate), 2) percentage of bare soil, 3) elevation, and 4) percentage of herb cover.The model suggests that regeneration increases with medium grazing intensity(compared to low as a reference or high intensity), the occurrence of bare soil,increasing elevation and higher cover of the herbaceous layer.The number of individuals of other species had also a significant influence,but were not included as predictor variables in the final model, as they were correlated with other predictors and not seen as independent.
In Fig.4,the number of individuals of N.dombeyi in the regeneration layer differentiated to the height classes I to V is depicted and related to wood grazing intensity which range from low to moderate and high.This indicates that the highest individual densities are found in forests with moderate grazing(almost 50%), followed by almost 30% of individuals in plots with high grazing intensity.On low-intensity wood pasture plots,the fewest numbers of individuals were found(~20%).
4.Discussion
Our main finding from the field study of tree regeneration is that N.dombeyi is favored by a moderate grazing intensity in the forest,taking grazing damage of tree individuals as proxy for grazing intensity, with the respective browsing index.This has been shown by the multipleregression analysis which revealed moderate grazing intensity as decisive ecological factor for tree regeneration as well as the occurrence of bare soil,which could also be a consequence of forest grazing.The herb cover can be seen as an indicator for soil moisture,which explains the positive effect of herb layer coverage on the regeneration(Dezzotti,1996;Caselli et al.,2019,2021b)but also for growth performance in adult N.dombeyi stands and tree canopy gaps.Although not investigated in our study, it should be mentioned that low litter cover favors the establishment of N.dombeyi seeds (Sola et al., 2020).Indeed, a thick litter layer can prevent seeds from reaching the soil to germinate and establish seedlings,particularly also related to the small seed size of Nothofagus species(González and Donoso, 1999; Christie and Armesto, 2003; Donoso Zegers, 2006).The depth of the forest floor organic humus layer and the humus forms were not a significant factor influencing seedling density in our study (Table 2), probably due to the generally low thickness and relatively high turnover of organic material.
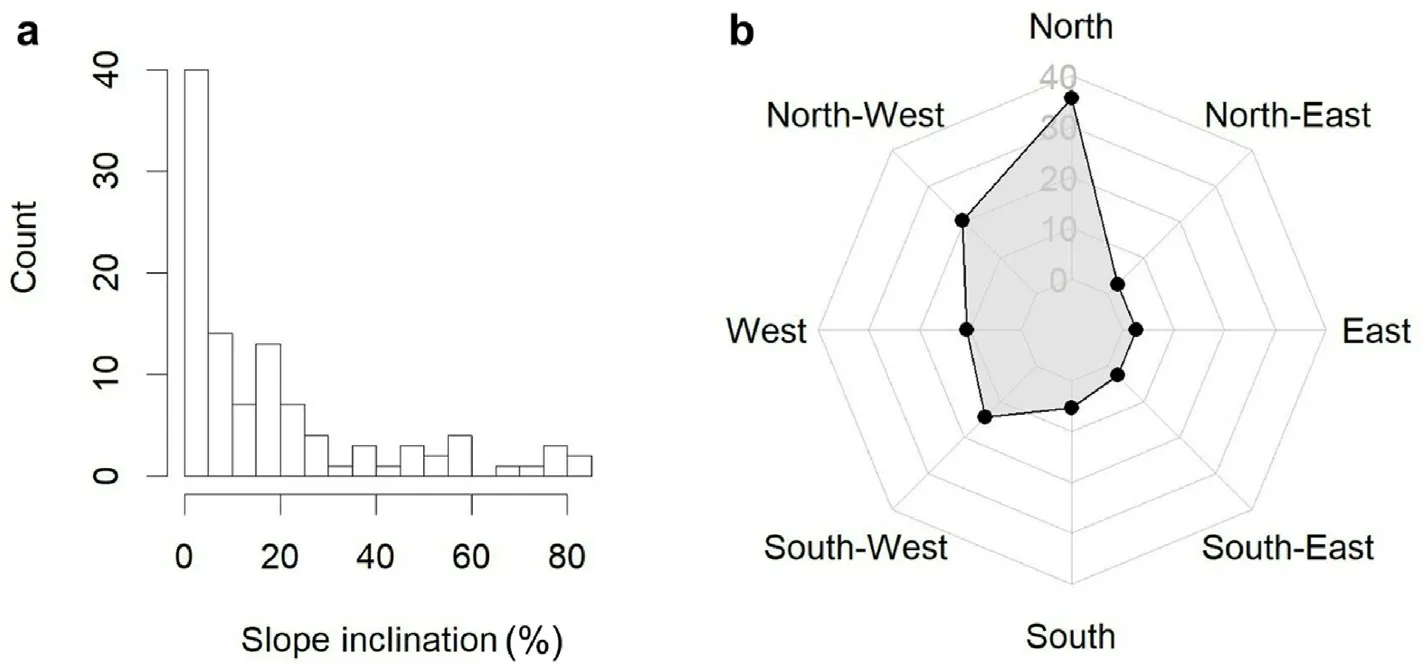
Fig.2.Slope inclination (a) and slope aspect (b) of sample plots along the studied transects.
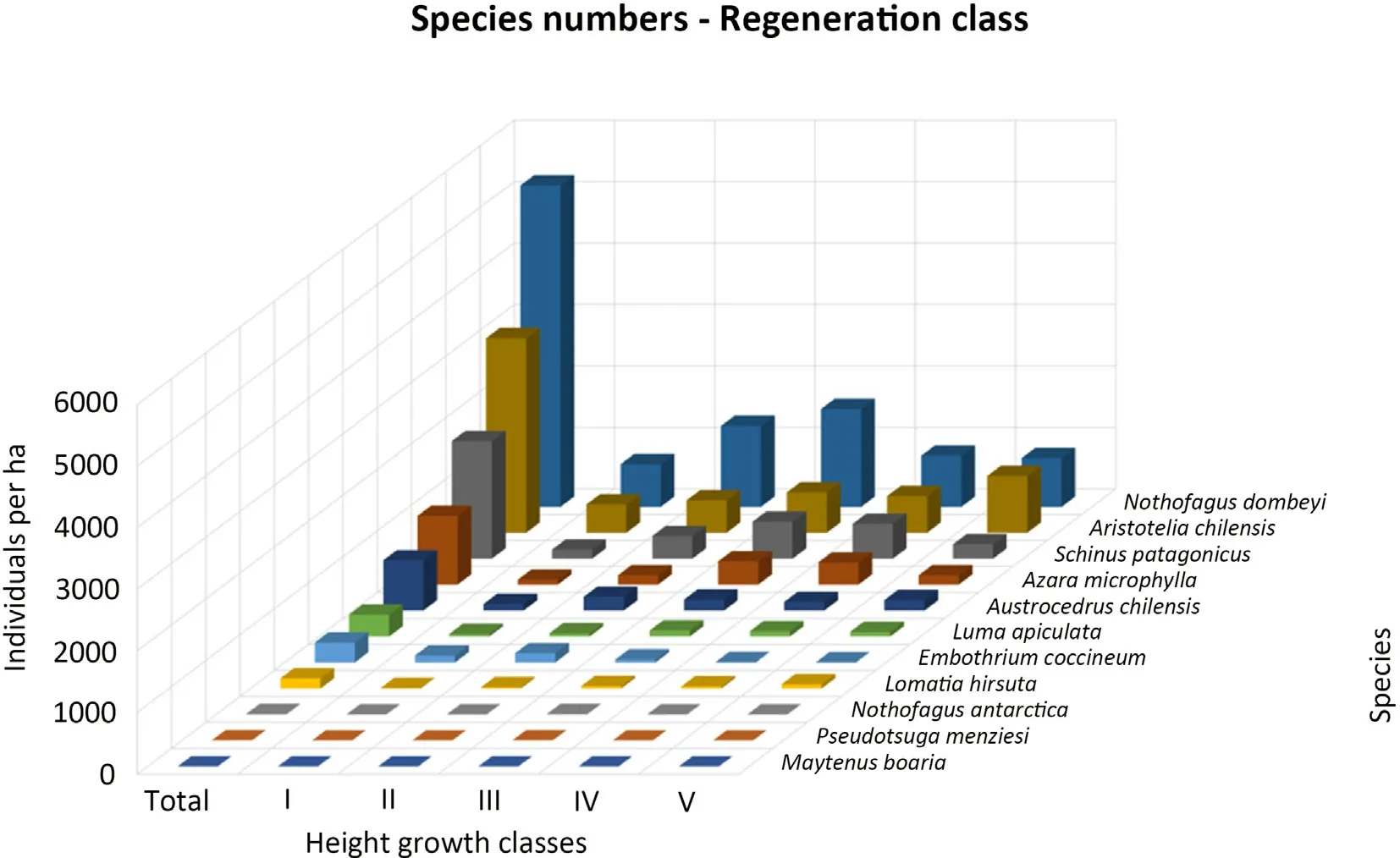
Fig.3.Mean individual numbers per ha of all 11 tree species recorded in the regeneration layers (height classes I to V) on the sample plots (n = 106 plots).
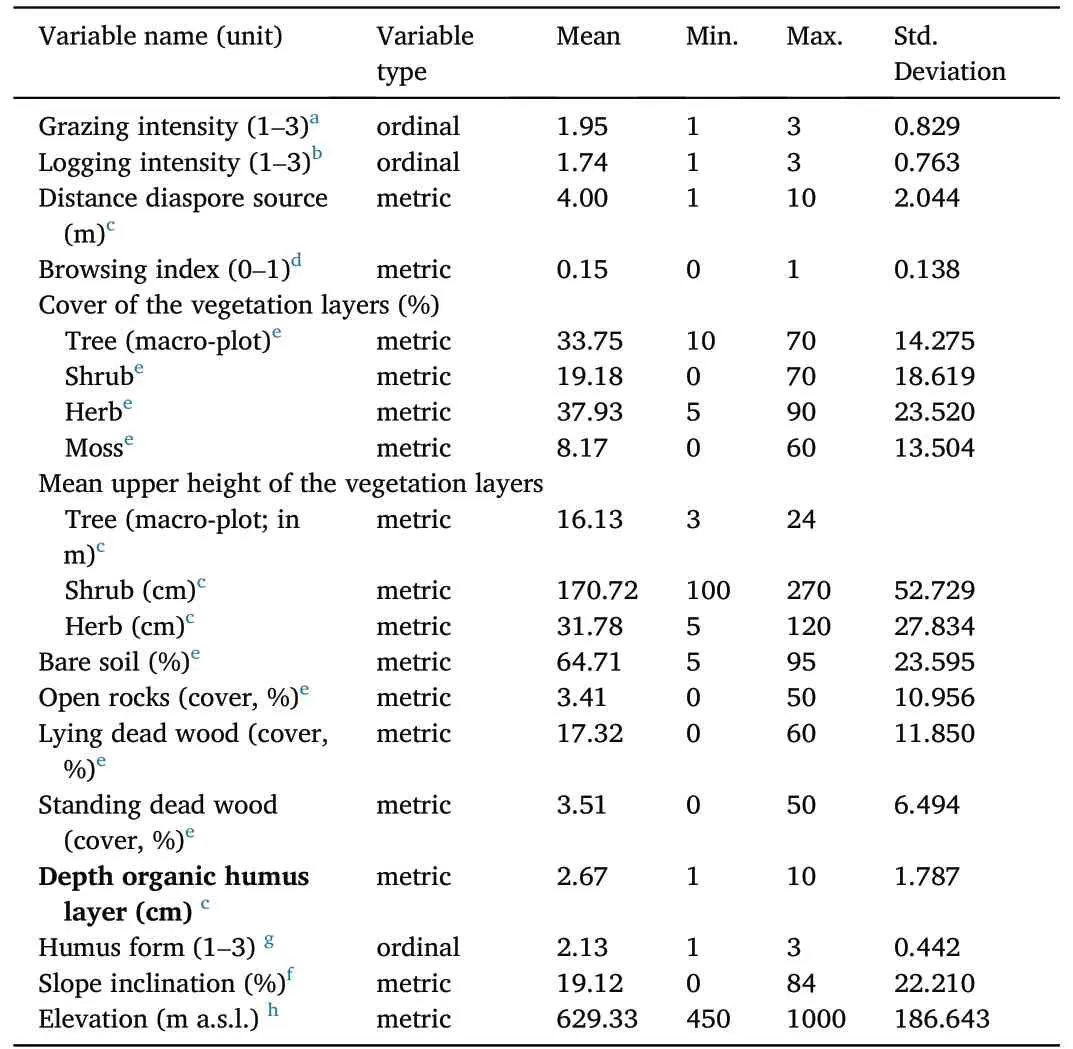
Table 2 Characteristics of the investigated forest stands (micro- and macro-plots);descriptive statistics of independent variables as potential influencing factors on the regeneration of Nothofagus dombeyi (n = 106 sampled plots).Sampling methods are indicated by superscripts and particular references are given as footnotes.

Table 3 Abundance of Nothofagus dombeyi regeneration in different height classes (n =106 sampled plots).
In our multivariate analysis, no significant relationship between the canopy cover as a measure for light availability and the regeneration of N.dombeyi has been found.In contrast,several studies have reported that the regeneration and growth of this tree species increases with light availability (Veblen, 1989; Donoso et al., 2013; Caselli et al., 2021b).Furthermore, Caselli et al.(2021a), state that the “lack of canopy cover can be beneficial for the growth[…]if moisture is not limiting”.At our study sites, light was obviously no limiting factor, which is partly a consequence of no records for sites without N.dombeyi regeneration that might be characterized by a higher canopy cover.Accordingly, onelimitation of our study is that we could not identify environmental factors which prevent N.dombeyi regeneration.
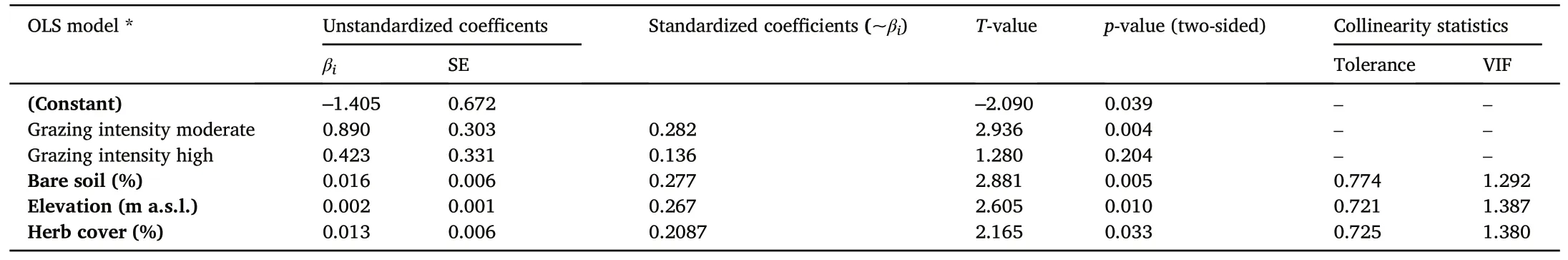
Table 4 Variables with significant effect on the abundance of Nothofagus dombeyi regeneration with ≤100 cm height.

Fig.4.Number of Nothofagus dombeyi seedlings(height class I)and saplings(height classes II-V)on 106 investigated sampling plots under different grazing intensities(low, moderate, high).
In our regression model, a linear relationship between N.dombeyi regeneration and elevation achieved the best fit.However, a unimodal distribution of N.dombeyi regeneration with increasing elevation is more likely and the data indicate a levelling off of successful regeneration at elevations between 650 and 1,000 m a.s.l.
We found no significant negative effect of competition with other recorded tree species or with conspecifics in larger height classes(IV,V)on N.dombeyi regeneration.Testing the count of other tree species as a predictor variable for N.dombeyi,revealed a collinearity,which could be interpreted either as facilitation or as aggregation at favorable sites for regeneration.An indication for the latter might be a relatively high cover of the herb layer and a positive relationship of N.dombeyi regeneration with the herb layer cover.No direct effects of logging on N.dombeyi regeneration were found.However, indirect impacts of logging (e.g.,exposure of bare soil)might well influence the N.dombeyi regeneration.
With regard to our studied species N.dombeyi and forest sites,we thus can make a plea for a careful consideration of forest grazing.This is supported by ¨Ollerer et al.(2019) who state several benefits of forest grazing, e.g., a potential cost-effective tool for forest management, the maintenance and enhancement of biodiversity, and the provision of higher income by combining forestry with agriculture(agroforestry).The latter has been also addressed by Quintéros et al.(2012)who found that the regeneration of Nothofagus pumilio did not change according to the observed intensity of livestock use.Particularly, considering the increasing risk of(mostly anthropogenically induced;see Mermoz et al.,2005) forest fires in Patagonia, forest grazing might have an important role in fire mitigation, reducing the flammability of forests through the reduction of the combustible load of the forest understorey (McEvoy et al.,2006b;Varela et al.,2018).
Whereas recommendations for the integration of forest grazing in forest management with regard to appropriate stocking levels and grazer species (e.g., Fraser et al., 2001; Papachristou and Platis, 2011) and controlled, rotational or periodical grazing (McEvoy et al., 2006a; Van Uytvanck and Hoffmann, 2009) are available from other forest ecosystems in the world, this has to be investigated under the ecological conditions of Patagonian forests and the given socio-economic situation.Peri et al.(2016,p.944)state that up to now“ranch managers mainly make grazing management decisions based on subjective criteria and previous experience” and“an economic assessment based on realistic production estimates and values is required to encourage the development of silvopastoral systems that could also offer benefits, such as erosion control,timber production and biodiversity conservation” (p.957).
For the regeneration of N.dombeyi, the low to very low dispersal velocity(Table 1)has to be taken into account.This has the consequence that a new establishment of trees after a large-scale disturbance needs time and is initiated by early-successional species, whereas old-growth stands regenerate towards the same forest community.Interestingly,N.dombeyi seems in some respects to combine the characteristics of a pioneer tree species (e.g., light-demanding, very rapid juvenile growth,low reproduction age) with those of a late-successional tree species(competitive strength in the canopy, sun and shade foliage differentiation,longevity).The shade tolerance of N.dombeyi seedlings is assessed as low to medium.Accordingly,open gaps in the forest canopy nurture its regeneration.The seed production highly varies from year to year,with a variation of mean seed mass, depending on the location.Since the dispersal is mainly via gravity, regeneration is found in low distance to the mother tree and thus,the seed source.Consequently,the spreading of this tree species is negatively affected in fragmented landscapes with other forest types or plantations with non-native tree species.
It might be worthwhile to compare the ecological characteristics of the two forest keystone species N.dombeyi (Patagonia) and Central European beech (Fagus sylvatica L.).With regard to certain structural and functional traits and the species’ role as a keystone species in forests,N.dombeyi has some properties in common with Central European beech,notably the ability to form monospecific stands as a climax species on a considerable large forest area.European beech covers an estimated range of ca.910,000 km2(Bohn and Gollub, 2007) and N.dombeyi a range of 156,120 km2(Barstow et al.,2017).
5.Conclusions for forest management
Based on the compilation of literature data on the ecology of N.dombeyi, the known threats of Patagonian forests by biotic (e.g.,invasive exotic conifers)and abiotic hazards(notably fire)as well as our findings on the regeneration of this tree species, we conclude for the management of N.dombeyi forests in the Patagonian Andes:
- All efforts in forestry and environmental policy have to be made to protect and sustainably manage the still existing contiguous,pure or mixed Nothofagus dombeyi forests and to restore those stands which have been degraded through human impact or have been damaged by fire.N.dombeyi may not only be seen as an ecological keystone species in the Patagonian Andes but also has the potential to become a much more important tree species for forestry and timber production.
- Forest grazing of low to moderate intensity favors regeneration through the provision of soil disturbance, thereby enhancing germination and seedling establishment.After the establishment of saplings,however,grazing should be excluded until the young trees have reached a growth height not affected anymore by browsing (>150 cm).The latter recommendation should not counteract sustainable forestry and a clear separation of forestry and agricultural land use like it is practiced in Central Europe.Recommendations for “close-to-nature”management for secondary mixed Nothofagus forests are provided, for example, by Loguercio et al.(2018).Forest grazing might be integrated on the farm or community level on land, where animal husbandry and timber production is combined within agroforestry systems (cp.Zerbe,2022).By favoring Nothofagus regeneration through a moderate grazing intensity which creates safe sites for seedlings and saplings (e.g., Soler et al., 2013) it should also be considered that further grazing might damage young trees and thus, decrease tree growth performance and productivity.Therefore,this integrated land-use approach of forestry and animal husbandry needs careful planning of multifunctionality in time and space and a careful monitoring of regeneration success.
Data availability
Data will be made available on request.
Authors' contribution
Conceptualization: SZ, GeL; Data curation: SS, GeL, AS; Formal analysis:SS,GeL,AS;Funding acquisition:HW,SZ,SS;Investigation:SS,SZ, GeL, NJ; Methodology: SZ, SS, GeL; Software: GeL, AS; Supervision:SZ,GeL,NJ;Validation:GeL,AS,GeL,HW,SH,GaL,JB,CL,MU;Roles/Writing-original draft:SZ,GeL;Writing-review&editing:all authors.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
This study is imbedded in the international and interdisciplinary research project“Sustainable forest management of temperate deciduous forests – Northern hemisphere beech and southern hemisphere beech forests”(KLIMNEM),a cooperation between the universities of Göttingen(Germany), Free University of Bozen-Bolzano (Italy), and the Andean Patagonian Forest Research and Extension Centre (CIEFAP, Argentina).The project is supported by funds of the Federal Ministry of Food and Agriculture(BMEL)based on a decision of the Parliament of the Federal Republic of Germany via the Federal Office for Agriculture and Food(BLE,grant No.28I-042-01).The University of Innsbruck provided travel funds for field work in Patagonia.
Appendix A.Supplementary data
Supplementary data related to this article can be found at https://doi.i.org/10.1016/j.fecs.2023.100126.
杂志排行

Forest Ecosystems的其它文章
- Climate and fire drivers of forest composition and openness in the Changbai Mountains since the Late Glacial
- Complexity responses of Rhododendron species to climate change in China reveal their urgent need for protection
- Differential seed removal,germination and seedling growth as determinants of species suitability for forest restoration by direct seeding – A case study from northern Thailand
- Response of fungal communities to afforestation and its indication for forest restoration
- Words apart: Standardizing forestry terms and definitions across European biodiversity studies
- Spatial niche segregation between bird species in the Białowie˙za primeval forest (NE Poland)