青海高寒草地水热因子对土壤微生物生物量碳、氮空间变异的贡献
——基于增强回归树模型
2023-10-07陈懂懂霍莉莉赵亮陈昕舒敏贺福全张煜坤张莉李奇
陈懂懂,霍莉莉,赵亮,陈昕,舒敏,4,贺福全,张煜坤,张莉,李奇*
1.青海三江源草地生态系统国家野外科学观测研究站/中国科学院三江源国家公园研究院/高原生物适应与进化重点实验室/中国科学院西北高原生物研究所,青海 西宁 810008;2.青海省青海湖景区保护利用管理局,青海 西宁 810000;3.中央民族大学,北京 100081;4.中国科学院大学,北京 100049
土壤微生物生物量是衡量土壤养分变化的指标,也是土壤有效养分储存库,并且是植物生长可利用和吸收的重要养分来源,反映了土壤微生物群落的相对大小、土地生产力和土壤健康状况,因其对环境具有较强的敏感性,所以能对土壤理化性状的演变做出快速响应(Bardgett et al.,1999)。虽然土壤微生物生物量只占土壤有机碳的1%-5%,却直接或间接参与几乎所有的土壤生物化学过程,在推动土壤物质转换、能量平衡和生物地球化学循环中起着重要作用(何亚婷等,2010)。土壤微生物生物量碳作为土壤有机碳中最活跃和最易变化的部分,直接参与养分循环转化等生物化学过程,同时,又与土壤C、N、P、S等元素的生物化学循环密切相关,是反映土壤微生物活性强度及有机质分解过程的重要指标,能很好地反映出土壤状况(管海英等,2014)。土壤微生物生物量N是土壤N素的一个重要储备库,并且具有较快的周转速率,因此在调控土壤氮对植物有效性和N素循环方面起着重要的调节作用(王国兵等,2011)。
土壤微生物作为土壤生态系统中物质循环和能量流动的核心指标,对环境因素的变化非常敏感(Fan et al.,2021)。土壤微生物生物量的变化往往受到土壤温度、pH、水分以及有机质含量等因素的影响(Alvare et al.,1995;Fan et al.,2021)。但是由于环境因子之间的互相干扰,土壤微生物生物量碳氮及其影响因子之间的关系研究还存在着不同的结论(蔡家艳,2017),亟需在不同的生态系统中加强研究。目前,对土壤微生物生物量碳、氮动态变化中的季节分布报道较多(李世清等,2004;何容等,2009;王国兵等,2011;管海英等,2015;刘放等,2020),且对其影响因素的分析也多集中在土壤理化性质、植被类型和土地利用等方面(何亚婷等,2010;蔡家艳,2017;仲波等,2017;邬嘉华等,2018;李雪萍等,2020;喻岚晖等,2020);而对其空间分布方面的研究较少(管海英等,2014;张稳,2015;覃乾等,2019),特别是在不同生境下水热因子对其影响的贡献如何更少见报道。而土壤微生物活动是在一定时空范围进行的(管海英等,2014),有必要从时间动态变化以及空间分布共同着手了解土壤微生物的活动特征。因此,本文主要研究青海高寒草地土壤微生物生物量在不同区域季节变异,以及土壤含水量、降水量、实际蒸散发、气温、土壤温度等水热因子对土壤微生物生物量碳、氮含量的变异的相对贡献,以期为研究青海高寒草地土壤微生物的季节特征提供依据,为系统认识区域土壤碳氮循环规律及其影响因素提供参考。
1 材料与方法
1.1 研究区域
青海省地处89°35′-103°04′E和31°30′-39°19′N,是青藏高原重要组成部分,平均海拔在3000 m以上;气候为典型的高原大陆性气候,全省年均气温在-5.1-8.5 ℃,年均降水量为50-550 mm(马昊翔等,2018),绝大部分地区的年均降水量低于400 mm(王亚晖等,2022)。境内河流众多,是黄河、长江主要水系的发源地,素有“中华水塔”和“江河源头”之称。作为中国五大牧区之一,青海省草地资源丰富,其中可利用面积约占全省天然草地面积的87%。草地类型主要包括高寒草甸类、高寒草原类、温性草原类、温性荒漠类、高寒荒漠类、低地草甸类、山地草甸类等(马昊翔等,2018)。该实验选取青海三江源草地生态系统国家野外科学观测研究站(简称“三江源站”)建立的5个长期监测样地,分别为海晏(HY)、同德(TD)、玛多(MD)、曲麻莱(QML)和可可西里(KKXL)(表1)进行相应研究。
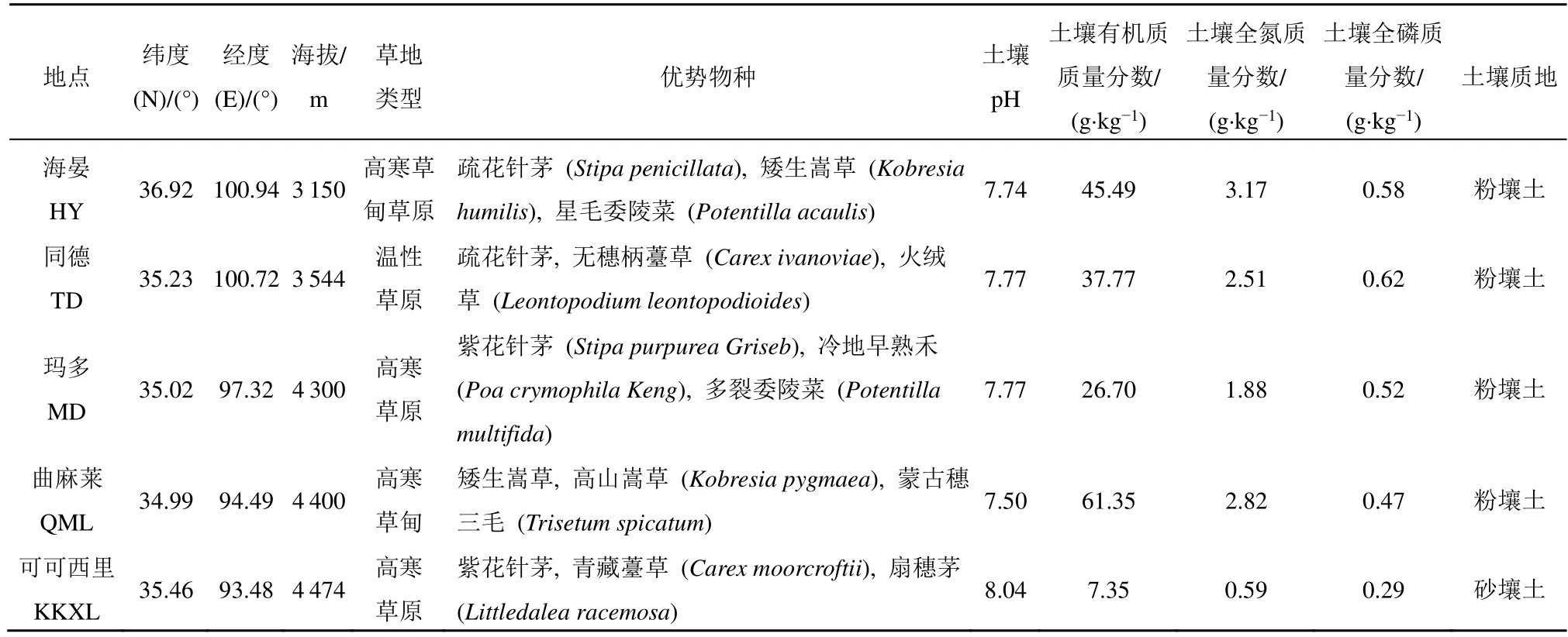
表1 研究区信息Table 1 Information of the observation stations
1.2 样品采集及数据收集
1.2.1 土壤样品采集
于2021年生长季(5-9月)的每个月中旬,在每个研究区的监测样地上,按照随机取样法设置5个采样点,每个采样点尺度约5 m×5 m,在该尺度内再随机分4层(0-10、10-20、20-30、30-40 cm)采取5点,同层土样进行混合得到该采样点的混合土样。连续5个月均在相同的区域内进行同样的土样采集。采集的土壤样品立刻带回实验室去除根系、石块等杂质后,先进行土壤含水量(Soil water content,SWC,%)的测定(于室内采用烘干法进行)(鲍士旦,2000);进行过筛处理后,一部分土样4 ℃保存用于土壤微生物生物量碳、氮的测定,一部分进行后续处理后用于测定其他指标。
1.2.2 水热因子采集
该研究中气温、降雨量等气象因子数据来自三江源站在5个研究区域架设的自动气象站和蒸渗仪(LYS-20PL)。自动气象站每半小时观测1次并通过北斗接受端(CR1000X)进行数据传输,降雨量(Precipitation,P,mm)、空气相对湿度(Relative humidity,RH,%)、气温(Temperature,t,℃)、土壤温度(Soil temperature,tS,℃)和净辐射量(Net radiation,Rn,MJ·m-2)均来自自动气象站的数据,渗漏量数据则来自蒸渗仪。选取2021年自动气象站观测的阶段储水量(W)、阶段降雨量(P)、阶段渗漏量(PD)日观测数据进行蒸散发(Evapotranspiration,ET,mm)的计算。对异常和缺失数据进行空缺值插补-K近邻算法插补进行修正(霍莉莉等,2022)。
1.3 指标测定及计算
1.3.1 土壤微生物生物量测定
土壤微生物生物量的测定采用氯仿熏蒸浸提法(Brookes et al.,1985;Vance et al.,1987):称取过2 mm筛的新鲜土样15.0 g(3份),放入 100 mL烧杯后放入干燥器,干燥器底部放入几张蒸馏水浸湿的滤纸、两个分别装有氢氧化钠溶液和无乙醇氯仿的小烧杯,并用凡士林封口,用真空泵抽取干燥器中空气直至氯仿沸腾并保持至少2 min,关闭干燥器阀门。将干燥器在25 ℃黑暗环境中放置24 h。培养结束后,将熏蒸土样转移至振荡瓶并加入60 mL 0.5 mol·L-1的硫酸钾(K2SO4)溶液,于150-200 r·min-1频率下振荡60 min,静置后过滤上清液。同时用0.5 mol·L-1K2SO4提取另外3份新鲜土样(土壤:溶液比例为1:4)作为对照样品。通过分别测定熏蒸土样与对照土样中的全氮、有机碳含量获得微生物生物量氮(Soil microbial biomass nitrogen,Nmic)、碳(Soil microbial biomass carbon,Cmic)。熏蒸和对照样品中的全氮用凯氏定氮法测定,有机碳则用重铬酸钾-硫酸氧化法测定。
EC——熏蒸土样与对照土样有机碳质量分数之差,mg·kg-1;
KC——计算Cmic的转化系数,为0.38(Vance et al.,1987)。
EN——熏蒸土样与对照土样全氮质量分数之差,mg·kg-1;
KN——计算Nmic的转化系数,为0.54(Brookes et al.,1985)。
1.3.2 蒸散发计算方法
水量平衡法:通过称重系统获取蒸渗仪总质量,根据前后的质量差计算每天实际蒸散量。由于实验地为地势平坦的区域,且降雨强度较小,因此不考虑生长季内地表径流量、地下水补给量的影响,根据水量平衡方程计算ET(霍莉莉等,2022),计算式为:
式中:
ET——实际蒸散发(mm);
WΔ——阶段储水量差值(mm);
P——阶段降雨量(mm);
PD——阶段渗漏量(mm)。
1.4 数据处理
增强回归树(Boosted regression tree,BRT)是基于分类回归树的一种机器学习算法。该算法在运算过程中通过多次随机抽取一定量的数据,分析自变量对因变量的影响程度,剩余数据用于结果的校验,对生成的多重回归树取均值并输出;可以计算在其他自变量取均值或不变的情况下,某自变量与因变量的相互关系,从而得出自变量对因变量的影响大小。BRT可以提高计算结果的稳定性和精度,并且在处理不同数据格式时具有很大的灵活性,输出结果较为直观(尹才等,2016)。同时,它通过生态因子对响应变量离差平方和的减少量以度量生态因子的相对重要性(张法伟等,2022)。因此适合用来定量分析微生物生物量及其影响因子之间的复杂关系。
本研究采用BRT模型定量评估水(SWC、RH、P、ET)、热(tS、t、Rn)因子对土壤微生物生物量变化的相对贡献。参与分析的气象数据为采集土壤数据当月该指标的平均值,与土壤微生物量数据一一对应。BRT模型利用R语言(R 4.0.2)的Dismo软件包实现,设置学习速率为0.001,树的复杂度为5,每次抽取90%的数据进行分析,并进行10次交叉验证。BRT模型依据预测变量在分类回归树中对响应变量离差平方和的减少量以度量因子的重要性,并对结果进行规范化处理,使所有预测因子的相对重要性之和为1.0,以百分数来表示单个预测因子的相对重要性(Elith et al.,2008)。所有图均在Origin 2019b软件中绘制(Origin Pro Inc.,USA)。
采用单因素方差分析(One-way ANOVA)评估微生物生物量在不同区域间的差异,分析有显著性(P<0.05)则进行LSD(Least significance difference)多重比较。利用Pearson correlation对整体的Cmic、Nmic与水热因子进行相关性分析,利用多元线性回归分析多因子对微生物生物量碳、氮的共同影响。分析均在SPSS 26.0中完成。
2 结果
2.1 研究区的水热因子变化
研究区地表年均气温变化为HY>TD>MD>KKXL>QML,其中最冷月均在1月,最热月均出现在7月。HY地区7月温度最高(13.42 ℃),KKXL最低(8.38 ℃)。研究区相对湿度(年均值)在区域间相差不大,生长季(56.88%-68.21%)高于非生长季(46.04%-52.04%)。研究区生长季年降雨量
MD (347.36 mm)>HY (344.57 mm)>TD (343.29 mm)>QML (333.54 mm)>KKXL (266.66 mm)。日净辐射量KKXL地区最高(11.10 MJ·m-2·d-1),MD地区最低(2.50 MJ·m-2·d-1),HY和TD相近,为5.7-5.8 MJ·m-2·d-1。研究区域实际日蒸散发在7-9月达到最大值,生长季其变化为HY (2.04 mm·d-1)> TD(1.63 mm·d-1)=QML (1.63 mm·d-1)>MD (1.57 mm·d-1)>KKXL (1.52 mm·d-1)(图1)。
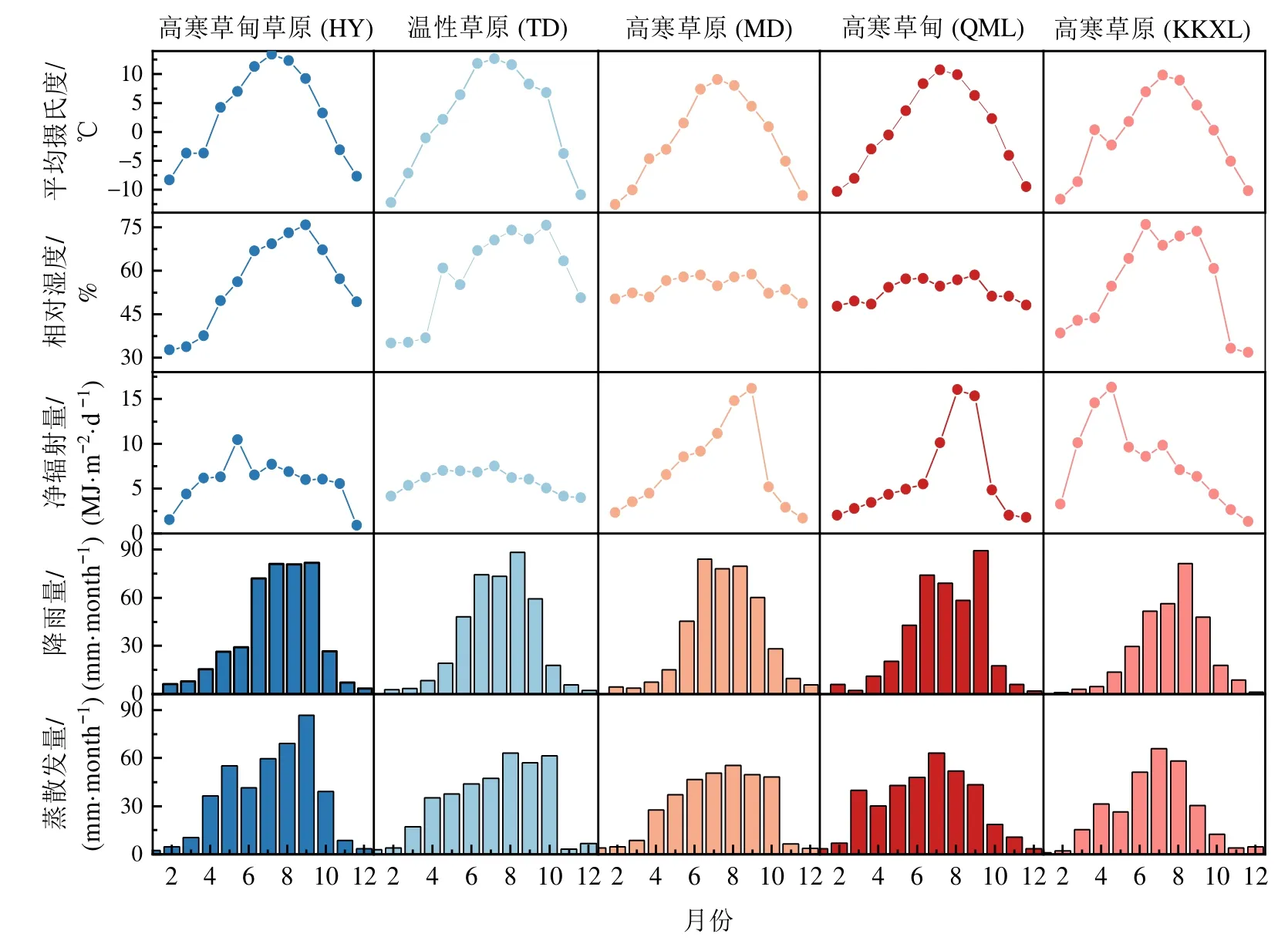
图1 研究区气象因子月动态Figure 1 Monthly dynamics of meteorological factors in study areas
SWC在QML最高,其次为HY和TD,MD和KKXL土壤含水量最低,具体变化为,第1层QML>TD>HY>MD>KKXL;第2层QML>HY>TD>KKXL>MD;第3层QML>HY>TD>KKXL>MD;第4层QML>HY>TD>MD(表2)。
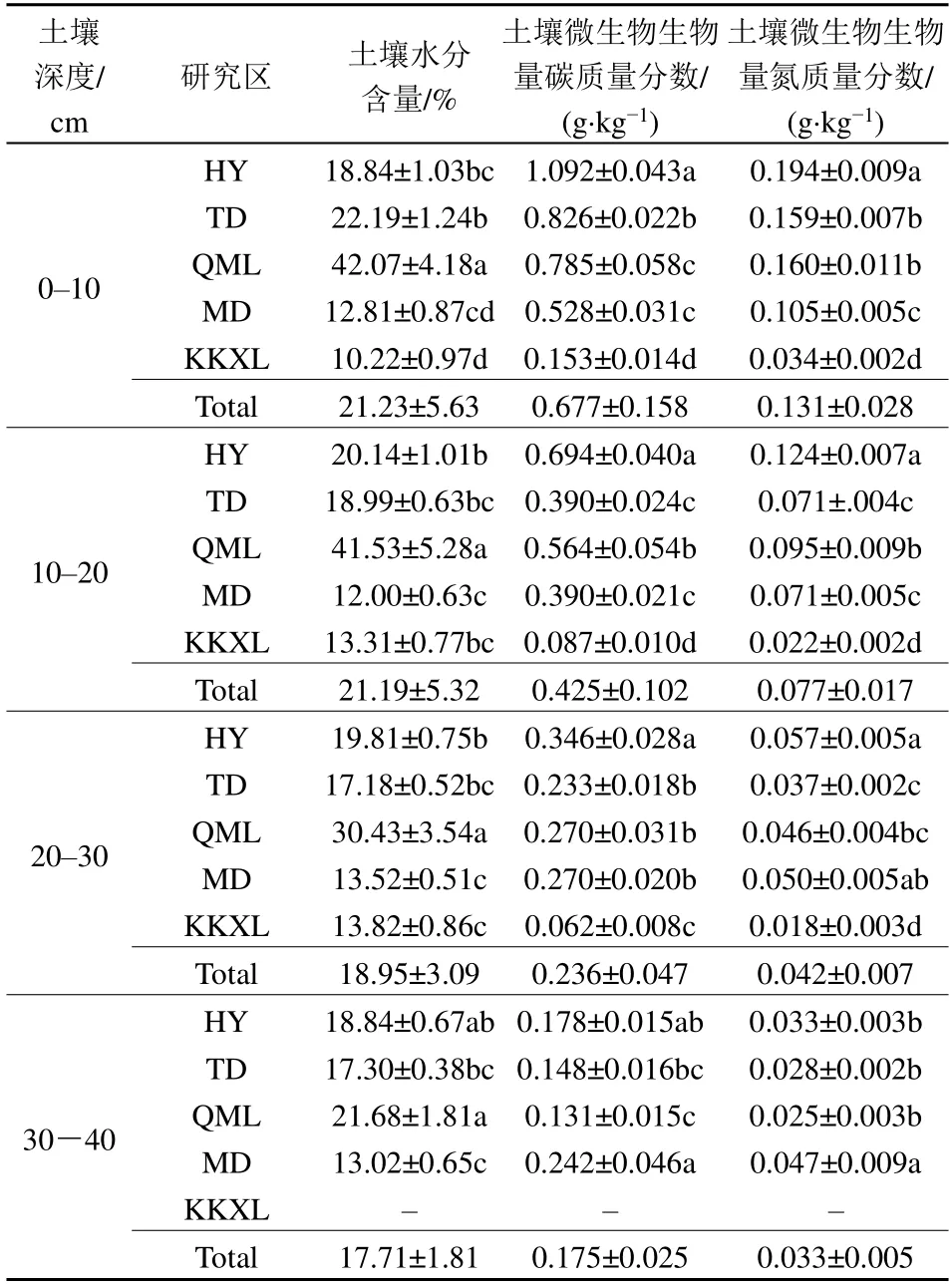
表2 土壤微生物生物量碳、氮特征Table 2 Characteristics of soil microbial biomass carbon and nitrogen
2.2 土壤微生物生物量碳的变化
研究区内Cmic均呈随土壤深度增加而下降的趋势。在0-10 cm土层,不同区域Cmic均值变化为HY>TD>QML>MD>KKXL;在10-20 cm土层,其变化为HY>QML>TD≈MD>KKXL;在20-30 cm,则HY>MD≈QML≈TD>KKXL;在30-40 cm,MD≈HY>TD≈QML。随着土层加深,Cmic从表层的温性草原>高寒草甸>高寒草原,到第4层则高寒草原>温性草原>高寒草甸。全区0-30 cm土层Cmic范围为0.105-0.710 g·kg-1,其中HY 0.710 g·kg-1,QML 0.540 g·kg-1,TD 0.483 g·kg-1,MD 0.398 g·kg-1,KKXL地区最低,为0.105 g·kg-1。从整个生长季看,处于青海省东部的HY和TD样地,Cmic最高值在5月,最低值出现在8月,呈现出随生长季而先降低后升高的变化规律;而位于中西部的QML、MD和KKXL样地,则最高值出现于7月的盛草期(表2,图2)。
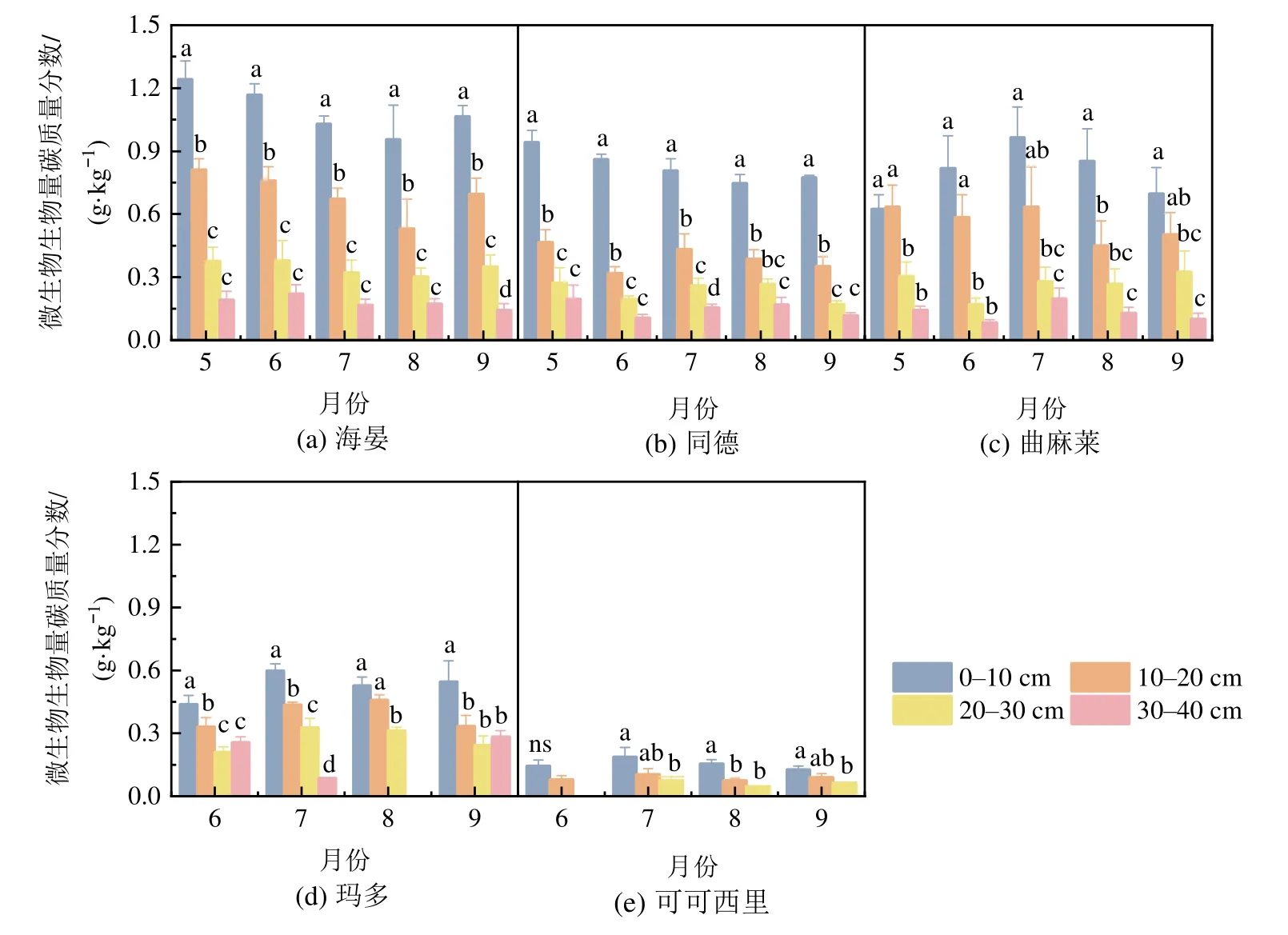
图2 生长季土壤微生物生物量碳动态Figure 2 Dynamics of soil microbial biomass carbon during the growth season
2.3 土壤微生物生物量氮的变化
与Cmic趋势一致的是,Nmic亦存在随土壤深度增加而下降的趋势。在0-10 cm土层,不同区域Nmic均值变化为HY>QML≈TD>MD>KKXL;在10-20 cm土层,其变化为HY>QML>MD≈TD>KKXL;在20-30 cm,则HY≈MD≈QML>TD> KKXL;在30-40 cm,MD>HY>TD≈QML。全区0-30 cm土层Nmic范围为0.024-0.125 g·kg-1,其中HY最高,KKXL区最低;QML 0.100 g·kg-1,TD 0.089 g·kg-1,MD 0.076 g·kg-1。在生长季,位于中西部的QML、MD和KKXL样地,Nmic最高值出现于9月,为随生长季逐渐升高的趋势。而位于青海省东部的HY样地,Nmic在5月最高,随后下降,到8月再一次升高;TD样地Nmic则在生长季先呈缓慢增加趋势而后又下降,最低值出现在9月(表2,图3)。
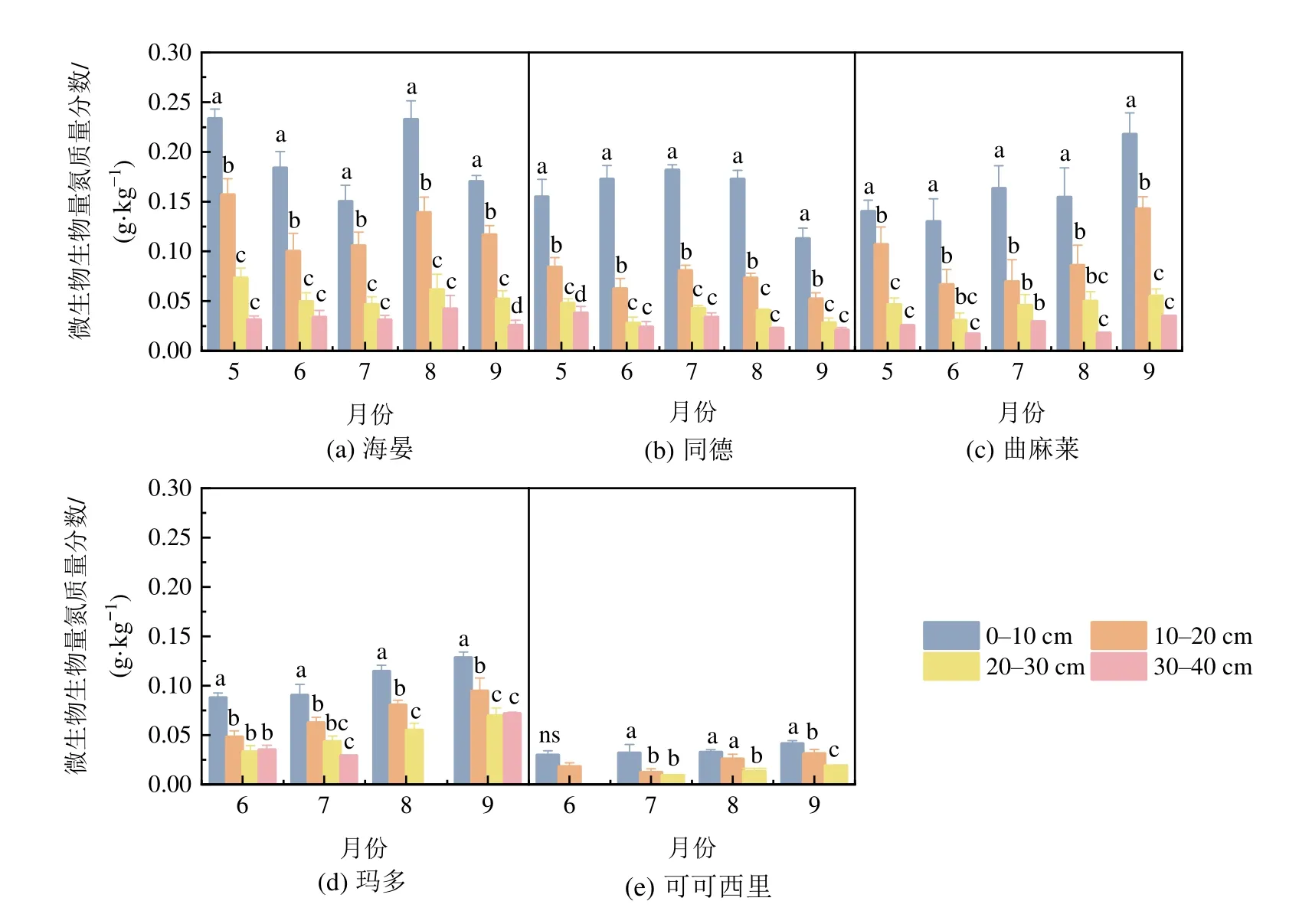
图3 生长季土壤微生物生物量氮动态Figure 3 Dynamics of soil microbial biomass nitrogen during the growth season
2.4 水热因子与微生物生物量的相关性
利用Pearson correlation对整体的微生物生物量碳、氮与水热因子进行相关分析,结果显示与Nmic显著相关的因子为SWC(r=0.337,t=7.356,P=0.000)和tS(r=0.222,t=4.535,P=0.000);与Cmic显著相关的因子除SWC(r=0.370,t=8.182,P=0.000)和tS(r=0.269,t=5.557,P=0.000)外,还有t(r=0.146,t=3.040,P=0.003)和Rn(r= -0.134,t= -2.768,P=0.006)(图4)。同时,多元线性回归分析的结果也显示,与Nmic相比,Cmic变化更容易受到多因子的共同影响,其中tS和SWC贡献最大(表3)。

图4 土壤微生物生物量与水热因子相关性Figure 4 Correlation between soil microbial biomass and hydrothermal factors

表3 土壤微生物生物量与水热因子的回归模型Table 3 Regression model between soil microbial biomass and hydrothermal factors
2.5 水热因子对微生物生物量空间变化的相对贡献
利用增强回归树模型定量评估水(SWC、RH、P、ET)、热(tS、t、Rn)因子对土壤微生物生物量变化的相对贡献。在土壤垂直梯度上,对Cmic和Nmic空间变化的影响,虽然水分因子对其垂直梯度变化的贡献均随土层加深而呈下降的趋势,热量因子的贡献率呈上升趋势,但不管在哪个土壤深度,水分因子对其影响均大于热量因子;尤其在0-30 cm,土壤含水量的贡献率是最大的。水热因子对Nmic的影响大于对Cmic的影响(图5)。
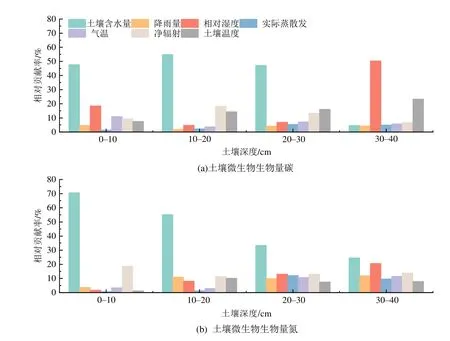
图5 不同土层水热因子对微生物生物量变化的相对贡献Figure 5 Relative contributions of water and heat factors in different soil layers to the variation of soil microbial biomass
就不同区域而言,在HY样地,决定Cmic、Nmic变化的是热量因子,其中tS的影响最大,相对贡献分别为45.23%和52.74%。在TD样地,对Cmic、Nmic变化贡献前两位的是SWC(分别为37.49%,62.23%)和tS(38.01%,27.19%)。在QML区域,SWC对Cmic、Nmic变化的相对贡献分别为92.37%,88.75%。在MD,tS决定了Cmic的变化,贡献为70.31%,而对Nmic变化的影响则ET最大(84.41%)。KKXL地区,对微生物生物量变化影响最大的也是SWC和tS,对Cmic,SWC贡献为40.56%,tS为30.85%;对Nmic,则SWC贡献最大为78.59%(图6)。综合区域来看,从东部到西部,对微生物生物量变异的影响,呈现从热量因子(主要为tS)向水分因子(主要为SWC)的过度,尤其是在对Nmic的影响上趋势更为明显。
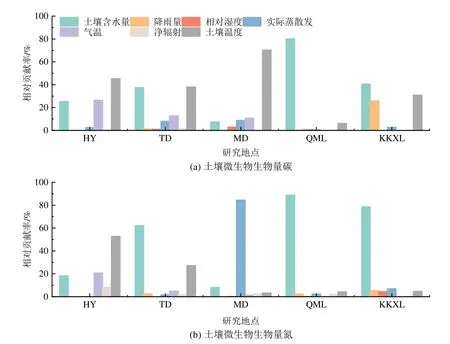
图6 不同区域水热因子对土壤微生物生物量变化的相对贡献Figure 6 Relative contributions of water and heat factors in different regions to the variation of soil microbial biomass
3 讨论
位于青海省中西部的高寒草原,尤其是可可西里地区土壤微生物生物量最低;而高寒草甸、高寒草甸草原和温性草原则相对较高。这是由于土壤微生物生物量碳、氮直接来源于土壤有机质的转化,且它们对可变的土壤条件非常敏感(Wang et al.,2018;Fan et al.,2021),若土壤质量差,土壤资源与土壤微生物之间的元素组成不平衡,土壤微生物无法支持持续的化合物合成,其生长、矿化和生态定居活动受到土壤养分和水资源有限的限制,微生物的生长和对养分的利用效率则降低(van Gestel et al.,2016;Fan et al.,2021),从而影响微生物对碳、氮的固定。而在环境条件较好的区域,微生物的分解和矿化作用会增强,微生物可以促进土壤养分的周转及养分的吸收(Jilling et al.,2018;Fan et al.,2021)。这与我们前期对土壤养分的研究结果一致(Chen et al.,2022)。另外,不同土壤深度间土壤的养分含量,水热条件,通气状况等的异质性往往会导致土壤微生物生物量在不同深度之间存在差异。表层的凋落物、密集分布的根系以及根系分泌物,提供了大量的有机物质;同时,表层土壤中好氧菌较多,活动较强,随着土层的加深,微生物的生境条件变差,深层土壤中生长代谢活动不强的厌氧菌占主导地位,微生物生物量的分布发生变化,因而土壤微生物生物量碳、氮明显低于上层(严登华等,2010;许华等,2020)。
土壤微生物生物量碳、氮在不同区域的季节变异存在差异,并没有在所有区域表现出春季高秋季低(许华等,2020),或者微生物生物量碳从春季到秋季的单峰变化而微生物生物量氮的“V”字型变化趋势(刘放等,2020)。这主要与不同生态环境能源的供应有关(Joergensen et al.,1994)。位于中西部研究区(QML、MD和KKXL)的微生物量季节动态基本一致,且明显区别于东部研究区。Cmic在生长季呈单峰变化,在盛草期达到最大值,这是因为夏季适宜的土壤温湿度为微生物的繁殖提供了良好的代谢环境;而Nmic则随生长季延长呈增加趋势,这可能是与在整个生长季研究区土壤氨氮含量的增加有关(谢梅珍等,2022)。东部区域(HY、TD)Cmic在返青期高,盛草期降低,生长季末期又上升的原因,可能是返青后随着气温上升,土壤微生物数量及活性的提高,对可利用残留物和土壤可溶性有机物的利用分解使得更多营养物质被微生物固定;另外,植物返青后根系分泌物的增加也为微生物提供了能源。植物进入生长旺季,需从土壤中摄取大量营养物质,从而限制了土壤微生物对养分的可利用性。而在生长季末,植物凋落物的归还以及根系的衰老等,增加了土壤碳源的输入,从而导致微生物量的增加。Nmic在HY和TD的波动幅度差异较大,可能与研究区土壤有机质含量差异有关(谢梅珍等,2022)。在水热条件相似的情况下,有机质丰富的土壤中(HY)具有更多可供分解周转的底物,而有机质含量较低的区域(TD)可供微生物利用的底物也有限,使得两区Nmic变化幅度产生差异。综合而言,微生物体与植物之间对营养物质的竞争,是影响土壤微生物生物量季节变异的主要因子。这种竞争关系在不同的生境中,随环境温度、湿度、植被组成以及可利用残留物的季节变化而存在差异(Garcîa et al.,1994;Alvarez et al.,1995;何容等,2009;王国兵等,2011;van Gestel et al.,2016;许华等,2020)。当微生物在竞争中获得更多营养时,或者当微生物群落产生较高的碳、氮利用效率时,因减少了“能量溢出”,而降低了能源维护成本,从而增加了用于增长的能源比例,更多物质被其固定,使其生物量增加(van Gestel et al.,2016)。而当植物的快速生长减少了碳水化合物和氮含量时,能量的不足使得微生物活动减弱,从而限制其生长繁殖,只有当土壤得到外源能量供应或植物根系旺盛生长能够提供较多有机物时才得到扭转(何容等,2009);但也可能因大量的根系分泌物的渗出而增加微生物活性(van Gestel et al.,2016)。
研究显示,陆地生态系统中的微生物对水分状况和温度等气候变化因素均有强烈的响应(Xu et al.,2016;van Gestel et al.,2016)。我们的研究也显示同样的结果:在土壤垂直分布上,对微生物生物量碳、氮影响最大和最直接的因子都是土壤含水量(P<0.001),且都是正向的,贡献较大的热量因子为净辐射和土壤温度(P<0.05)。随着土层加深,水分因子影响呈减弱趋势,可能的原因是深层土壤紧实度的增加和土壤孔隙的减少阻碍了水分的入渗,而增加了土壤热导率和热容量(杨晓娟等,2008)。而在不同区域上,虽然水分和热量因子对微生物生物量变异的贡献差异较大,土壤温度和土壤含水量仍是最主要的两个因子。但在区域上由东向西,影响土壤微生物量的因子由热量因子向水分因子过度,这可能是因为东部区域降水较多气候湿润,水分对微生物而言并不是限制因子,而西部区域往往雨水少而干旱,水分更容易限制微生物的活性(李红英等,2022)。前人的很多研究都显示土壤含水量和土壤温度对于土壤微生物生物量的形成是优势因素(张崇邦等,1996;管海英等,2014;蔡家艳,2017)。因为土壤水分是维持土壤微生物正常代谢活性不可缺少的物质,一方面水是微生物原生质的重要组成部分(约占微生物组成的90%左右),任何时候水分的缺失,都将严重影响到微生物自身的基本特征(Hu et al.,2023);另一方面水分能够通过影响土壤的通气状况、土壤温度、酸碱度及含盐量等生态因子间接地作用于微生物(何亚婷等,2010)。另外,由于微生物的生命活动都是由一系列的生物化学反应组成的,而这些生化反应又受到温度的强烈影响,因此,温度也是影响微生物数量与活性的重要因子(何亚婷等,2010)。研究显示,在低于35-45 ℃时,土壤微生物活性是随温度升高而上升的(张稳,2015),这与我们测得的温度较高区域土壤微生物生物量高而温度较低地区微生物生物量低的结果是相对应。可能的原因是适宜的土壤温度促进了土壤微生物的活性及碳氮利用效率(覃乾等,2019)。
4 结论
青海省高寒草甸、高寒草甸草原和温性草原的土壤微生物生物量明显高于高寒草原。不同区域高寒草地土壤微生物生物量表现出不同的季节动态,尤其是中西部区域表现出与东部区域明显不同的趋势。在青海高寒草地,土壤含水量和土壤温度是控制土壤微生物生物量变异的主要因子。从青海东部到西部,对微生物生物量变异的影响,呈现出从热量因子(主要为土壤温度)向水分因子(主要为土壤含水量)的过渡。
