木荷优树自由授粉家系生长性状分析与早期选择
2023-09-28郑如德钱国财潘昕汤行昊范辉华
郑如德,钱国财,潘昕,汤行昊,范辉华
(1.福建省南平葫芦山国有林场,福建 南平 353016;2.福建省林木种苗总站,福建 福州 350003;3.福建省林业科学研究院,福建 福州 350012)
近年来,我国高度重视乡土珍贵树种种质资源的发掘和保护,加强珍贵用材林资源培育。木荷(Schimasuperba)属山茶科(Theaceae)木荷属(Schima)常绿阔叶乔木[1, 2],广泛分布于我国南方各省区[3],因其具有适应性强、速生丰产性高、材质优异且造林成效好等特点,成为我国重要的高效生态树种和生物防火林带建设的主栽树种[4, 5],也是我国南方重点发展的珍贵优质阔叶用材造林树种之一[6]。我国木荷育种始于2001年,但至今仍未完全实现良种化造林[7]。长期的突变、隔离及自然选择,导致木荷种内变异极其丰富[8, 9],由于现有优良育种材料的缺乏,在一定程度上导致了当前木荷种植规模偏小和推广应用受限[7]。因此,木荷的良种选育一直是人们关注的问题之一[10]。优良种源和家系的选择是林木遗传改良的有效手段和良种选育的基础[11]。因此,进行木荷优良种源和家系选择对加快良种的繁育进程、促进木荷人工林发展、提高木荷资源开发利用水平具有重要的理论意义和实用价值。本研究以营建在福建省南平葫芦山国有林场樟湖分场的5年生34个木荷优良单株半同胞家系试验林为材料,开展了树高、胸径、冠幅等9个林木生长性状指标的调查,以分析生长性状的家系间遗传变异和育种值的综合评分,初步筛选出优良速生家系,同时为育种亲本的选配及逆向选择提供了参考依据。
1 材料与方法
1.1 试验材料与试验设计
试验林位于福建省南平葫芦山国有林场樟湖分场黄龙岗工区(东坑山头077 林班 54大班 011小班),樟湖林场地处118°27′ E、 26°26′ N,属武夷山戴云山山间立地亚区,海拔200~300 m,为丘陵地带,地形平缓,坡度为20~30°,年降水量1 600~1 900 mm,年平均湿度 79%;最高气温为40 ℃,最低气温为-3 ℃,霜期短或无霜期。土壤为花岗岩发育的山地红壤,土层中等厚度,为90~100 cm,土壤肥力为Ⅱ级。植被以檵木(Loropetalumchinense)、苦竹(Pleioblastusamarus)、五节芒(Miscanthusfloridulus)、芒萁(Gleichenialinearis)为主,高度50~150 cm,盖度约95%。
试验材料来自福建省建瓯、延平、古田和江西省崇义、龙南等5个县市34个初选木荷优树的半同胞自由授粉家系,2017年采用1年生轻基质容器苗营建家系测定试验林,完全随机区组设计, 10株顺坡单列小区、3次重复,株行距为2 m×2 m,造林地前茬为1代杉木纯林,采伐迹地全面劈草和耙带清理,挖明穴,规格为50 cm×40 cm×30 cm,区组之间种植枫香隔离行,试验地外围种植杉木纯林,造林后1~3年每年分别于5—6月、9—10月各除草扩穴培土1次,第2年5—6月结合抚育沟施复合肥100 g·株-1,并开展除萌1次。
1.2 林分调查和数据处理
2021年底,对5年生木荷家系测定林开展全林每木调查,测定指标包括树高、胸径、冠幅、树干通直度、侧枝性状(最大侧枝长、最大侧枝粗和分叉干数)等。采用标尺及围尺测定树高和胸径;冠幅取东西与南北的平均值;树干通直度按通直、较通直、一般、弯曲、严重弯曲划分为5级,分别量化为5、4、3、2、1,数值越大越通直。
(1)单株材积的采用木荷人工林二元材积计算[12]:
V=6.801×10-5D1.865 613H0.918 129
式中:V为单株材积(m3);D为单株胸径(cm);H为单株树高(m)。
(2)家系遗传力计算[13]:
式中:F为方差分析值。
(3)表型变异系数[14]:
(4)遗传变异系数:
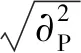
(5)一般配合力:

(6)使用表型观测值计算方法统计家系各性状育种值[15]:
(7)遗传增益计算公式:
2 结果与分析
2.1 木荷家系生长性状方差分析
方差分析结果显示(表1),34个木荷家系间各性状差异均达极显著水平(P<0.01),表明不同家系间木荷生长具有明显的家系遗传差异性,这为优良家系的选择提供了丰富的变异基础。本研究对木荷家系进行评价选择具有重要意义。
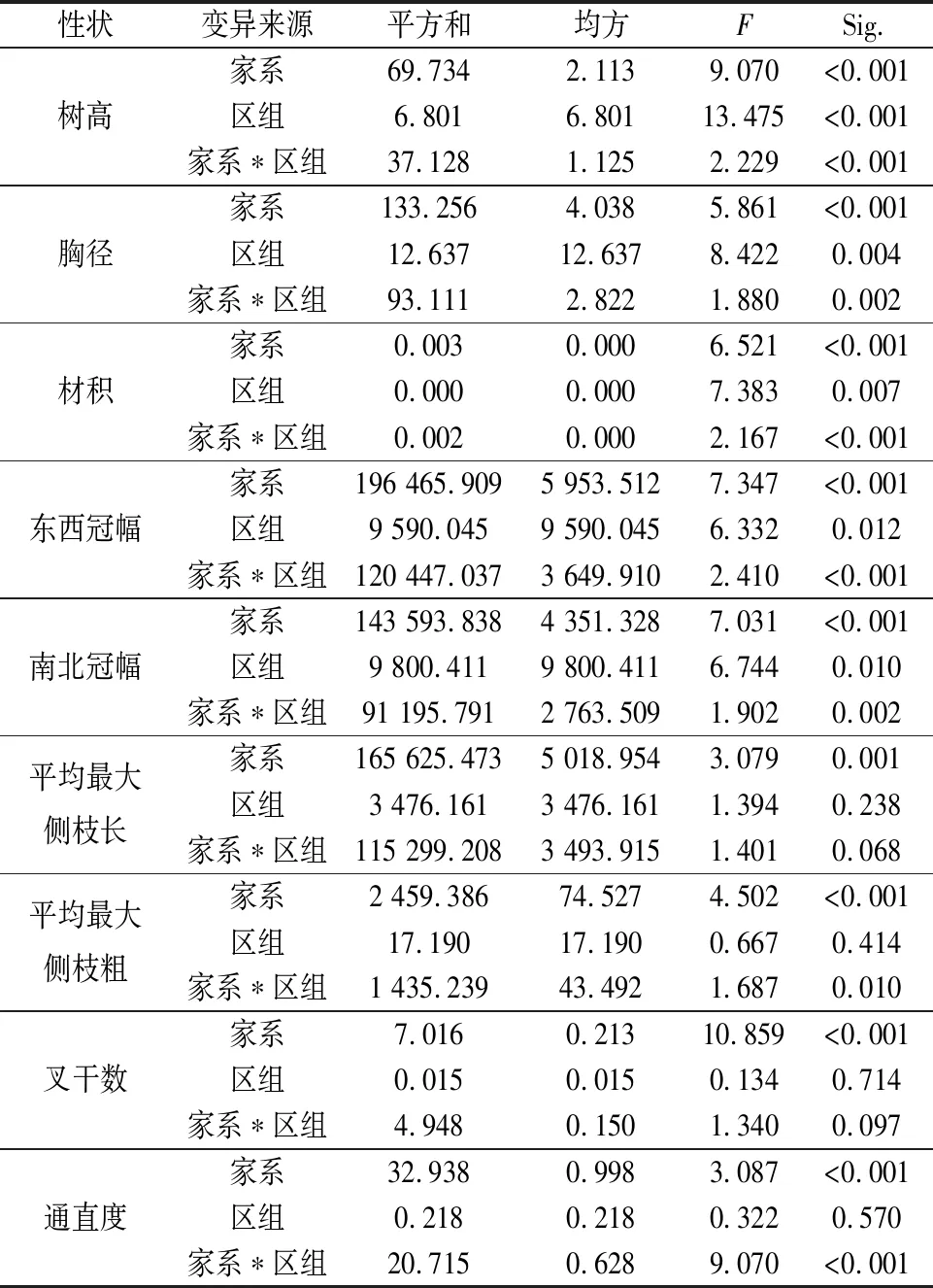
表1 木荷各家系生长性状方差分析
2.2 木荷家系间生长性状的变异分析
34个木荷家系树高、胸径、冠幅、侧枝性状、叉干数和通直度等变异参数分析结果见表2。
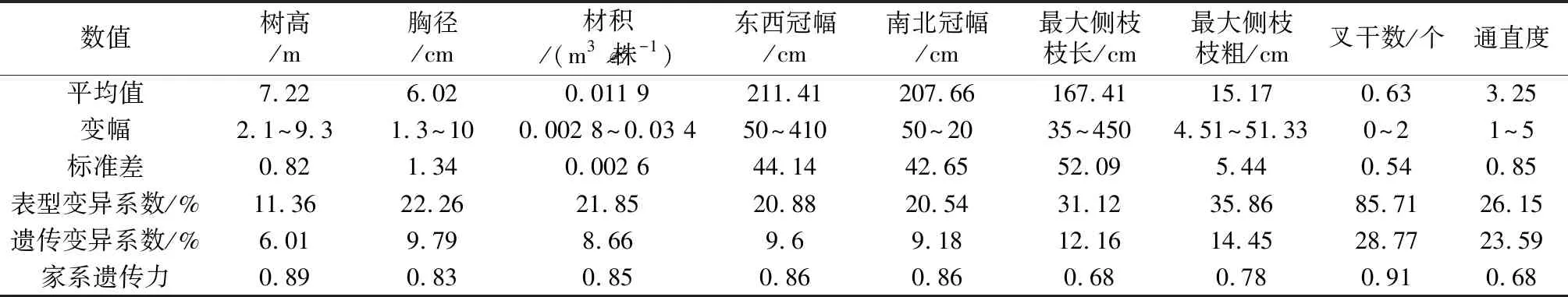
表2 34个木荷半同胞家系各性状平均值及变异系数
变异系数可以反映不同家系间各指标的变异情况,变异系数越大,表明变异越大,越有利于优良家系的评价选择[16, 17]。本研究中,34个木荷半同胞家系不同性状表型变异系数变化范围为11.36%~85.71%,其中,表型变异系数最低的是树高,为11.36%,最高的为平均叉干数,达到了85.71%;而胸径和单株材积的表型变异系数分别为22.26%和21.85%。遗传变异系数能反映由遗传因素造成的性状观测值变异的程度[18]。34个木荷半同胞家系各性状的遗传变异系数变化范围为6.01%~28.77%,树高、胸径和单株材积相对较低,分别为6.01%、9.79%和8.66%,而叉干数和通直度则相对较高,分别为28.77%和23.59%。木荷生长各性状遗传变异系数约占表型变异系数的43.98%~90.2%,表明变异不同程度地受到了遗传和生长环境的控制。
遗传力是林木性状的重要遗传参数,是亲本某一性状遗传给子代的能力,反映不同性状表型变异中遗传因素和环境因素作用的大小关系,其值越大则性状越稳定,受环境作用越小[13, 17]。34个木荷半同胞家系各生长指标的遗传力均较高,均超过了0.65,为高遗传。其中,树高、胸径、单株材积、叉干数及冠幅生长均超过了0.80。这表明木荷家系幼林生长受较强的遗传控制,各性状在不同家系内均能稳定遗传,受环境影响小,故而进行家系试验及家系选择研究,对提高木荷的经济效益有着重要作用。
2.3 木荷家系不同生长性状的相关性分析
由表3可知,树高与除平均叉干数以外的所有性状均达极显著正相关,其中与材积之间的相关系数最高,为0.711,胸径与除平均叉干数以外的所有性状均呈极显著正相关,其中与材积之间的相关系数高达0.976,但与平均叉干数呈极显著负相关。材积与除平均叉干数以外的所有性状均呈极显著正相关性,但与平均叉干数呈极显著负相关。除树高、胸径和单株材积外,冠幅生长也与最大侧枝长和侧枝粗呈极显著正相关性,冠幅与胸径和材积间的相关系数均在0.60以上,但冠幅与平均叉干数和通直度分别呈极显著负相关和显著正相关。除此之外,木荷平均最大侧枝长和侧枝粗均与平均叉干数呈极显著负相关,与通直度间无显著相关性。但平均叉干数与通直度间达极显著负相关,表明叉干数较多的家系单株整体通直度较小。

表3 34个木荷半同胞家系各性状相关性
综上数据表明,木荷家系生长性状之间均有一定的显著或极显著的相关性关系,表明生长指标之间存在极显著相关性和制约性,尤其是材积和胸径、树高之间,两两相关系数均在0.70以上。因此,在选择优良家系时,应密切关注材积、胸径和树高生长量。
2.4 木荷家系材积一般配合力分析
一般配合力(general combining ability,GCA)是评价亲本利用价值的重要指标,在林木遗传改良实践中发挥着十分重要的作用[11, 19]。通过子代测定数据可以选择出优良的亲本,在作物育种中配合力被广泛应用,已经取得了较好的效果。利用一般配合力较高的亲本进行杂交能够提高得到杂种优势的概率[16]。木荷家系材积一般配合力计算结果见表4,34个木荷半同胞家系中,材积一般配合力变化范围为-0.006 623(家系JLS24)~0.005 75(家系GT34),其中具有正效应的家系有15个,占参试家系的44.11%,家系GT34、家系YP26和家系GT23的一般配合力相对较高,分别为0.005 75、0.005 33和0.003 99,这一结果表明,利用以上3个家系的亲本作为杂交母本,子代材积遗传改良潜力较大。

表4 34个木荷半同胞家系材积一般配合力
2.5 木荷优良家系初选
优良家系的选择应从具体培育目标出发。本次选择以用材林为目的,以生长性状为主,同时兼顾形质性状,以选出胸径、树高、材积等生长迅速,干形通直且冠幅较小以便合理密植的优良家系种质。因此,为能科学合理,避免单纯考虑某一性状而导致筛选家系的局限性,且在家系间树高、胸径和单材材积性状显著差别的前提下对各参试家系进行多性状的综合评定,选出的家系更稳定[14, 20]。
育种值是林木遗传育种中的重要参数,由群体平均值加上遗传力与选择差的积组成。利用木荷家系不同性状的遗传力估算结果,进一步计算出不同性状的育种值,再根据育种值大小进行排序,每个性状的育种值排列第 1 位赋予 34 分,育种值排列第 2 位赋予 33 分,依次类推,育种值排列最后 1 位赋予 1 分。最后将不同性状得分值相加作为每一个家系的总分值,再根据总分值对34个木荷家系进行排序[14, 15]。结果表明(表5),34个家系的总得分为3~97分,平均分值为52.8分,总得分超过群体得分均值的优良家系共有19个。其中,家系GT34得分最高,为97分;其次为GT23,总得分为94分;而家系JLS24的总得分最低,仅有3分。这表明,GT34家系的性状综合表现相对最好,而JLS家系表现最差。排在第一位的CT34家系总得分为家系群体平均得分的1.84倍。
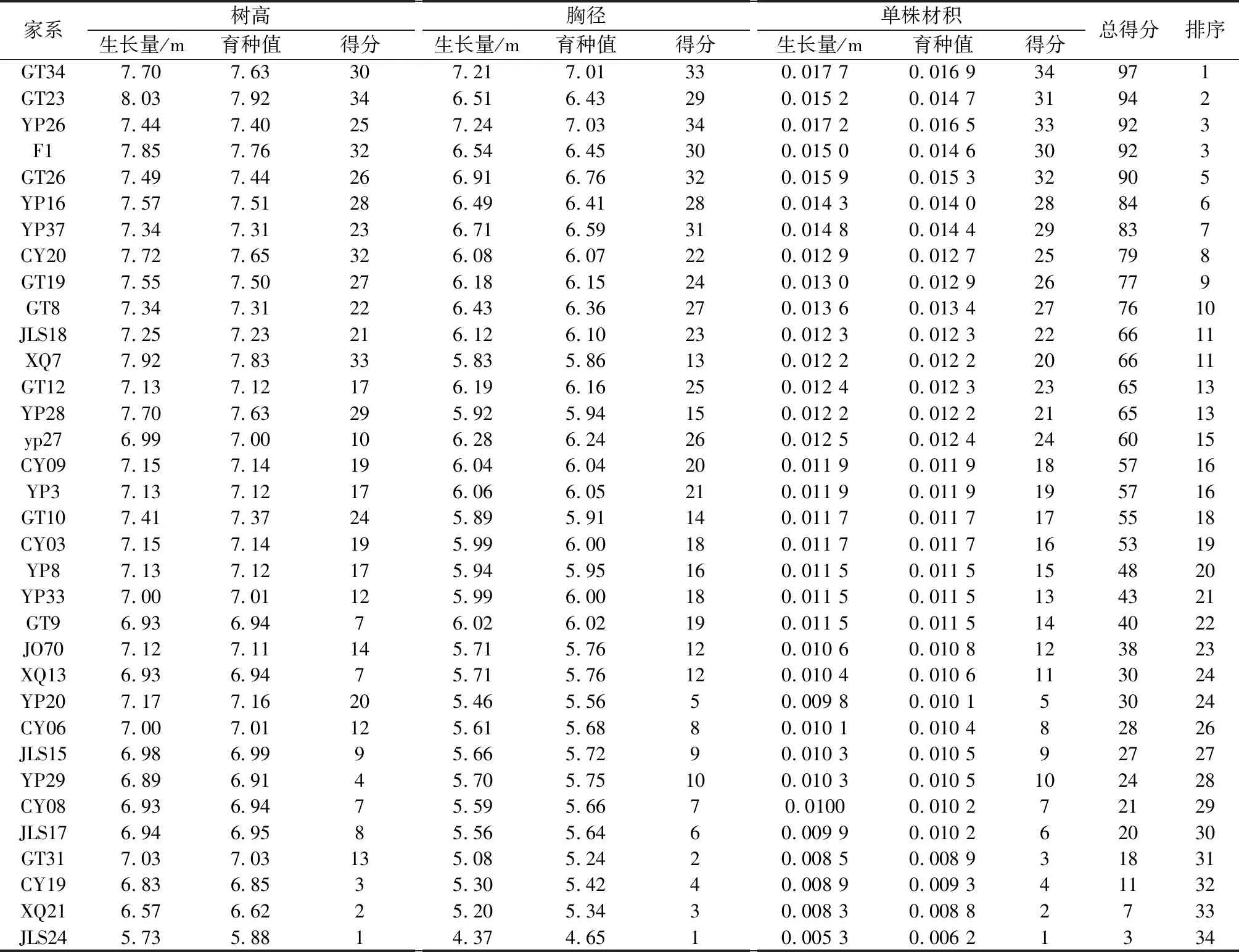
表5 木荷34 个半同胞家系综合性状育种值分析
按10%的入选率进行选择,家系总得分位列前4名的GT34、YP26、 F1和 GT23初选为最优家系,其树高平均值为7.755 m,比群体总平均值提高了7.41%,遗传增益为6.60%;胸径平均值为6.875 cm,比群体总平均值增加了14.2%,遗传增益为11.79%;单株材积的平均值为0.016 m3,比群体总平均值提高了36.63%,遗传增益为31.47%。
3 结论与讨论
3.1 遗传和变异是林木育种研究的主要内容,选择优良的家系和单株用于营建种子园生产良种并加以推广应用,是良种选育和生产相结合的首选方式。本研究中,34个半同胞家系间各性状方差分析均达到显著或极显著水平,这表明,不同木荷家系间具有显著性差异,木荷家系间生长性状受高强度的遗传控制,具有较强的家系选择潜力。
3.2 本研究筛选出了生长和稳定性表现最优的4个半同胞家系GT34、YP26、 F1和 GT23,入选家系的平均树高、胸径和单株材积分别为7.755 m、6.875 cm和0.016 m3,入选的家系对不同性状改良程度存在差异,树高和胸径平均遗传增益分别为6.60%和11.79%,而材积的遗传增益最高,为31.47%。这与覃敏等[14]对米老排、梁德洋等[21]对红松、徐悦丽等[13]对红松的研究结果相似。这4个家系可以作为优良家系直接应用于生产。同时,通过综合评价,家系YP26、GT34和GT23的一般配合力明显更高,一般配合力高的亲本可以产生相应性状表现好的子代,故可作为木荷后期遗传改良的育种亲本。筛选出的速生性显著的优良家系既可作为木荷种子园建园材料进行遗传改良[22, 23],也可通过无性繁殖手段发挥生产潜力[24]。
3.3 相关性分析可以体现各性状之间的关系,树高、胸径和单株材积间两两相关的相关性系数均超过0.7,尤其是胸径与材积之间的相关系数高达0.976,表明材积与胸径存在极强的相关性。
冠幅与胸径和材积间的相关系数均在0.60以上,同时也与最大侧枝长和侧枝粗呈极显著正相关,树高和胸径生长量大的家系通常具有更大的冠幅和较粗的分枝,这可能是由于冠幅越大,接受光照越多,光合作用越强,使树木生长发育旺盛[25, 26]。树高、胸径、材积与除平均叉干数以外的所有性状均呈极显著正相关性,但均与平均叉干数呈极显著负相关。同时,平均叉干数与通直度极显著负相关,表明叉干数较少的单株生长更快,且叉干数较多的家系单株整体通直度较小。对速生的木荷家系,需加强早期修枝,以培育优质的干材[7]。
3.4 众多研究表明,速生树种适宜进行早期选择时间在2~8年[27-30],如刘宇等[27]对6年生的白桦杂交子代为研究对象进行优良杂交组合的早期选择,王婧等[31]对4年生黄樟优良种源和家系的早期选择,覃敏等[14]对6年生米老排种源及家系进行的综合评价,初步筛选出优良的种源和家系,都取得了良好的效果。因此,本研究针对5年生的木荷半同胞家系为研究对象进行优良家系的早期选择是可行的。通过评价筛选,去劣留优,从而提高种子园的遗传增益,这是林木遗传改良的主要环节之一。但苗木生长的稳定性及生长适应性也同样是评价林木家系优劣的重要指标[27]。本研究虽然初步筛选出适宜本地生长的优良家系,但对其后期的生长表现和适应性仍有待于跟踪和观测。
此前研究发现,木荷苗木生长具有明显的种源变异,且与立地间存在明显的互作效应,故在一个试点筛选出的优良家系仍无法直接地推广应用于与本试点立地生境迥异的其他区域,同时,单点造林试验结果也无法为家系生长稳定性的适应范围及木荷家系基因型和生长环境的互相效应提供更有力支撑[7, 31-33],因此,开展多点联合的木荷家系生长跟踪尤为重要,为今后不同生态区优良家系的推广提供可靠的保证。
