毕节地区6个绵羊品种FecB基因微卫星座位多态性研究
2023-09-28宋德荣吴瑛林如涛单春兰马金萍孙忠鑫熊艳玲刘勇庆胡飞飞
宋德荣, 吴瑛, 林如涛, 单春兰, 马金萍,孙忠鑫, 熊艳玲, 刘勇庆, 胡飞飞
1. 毕节市畜牧兽医科学研究所,贵州 毕节 551700;2. 贵州大学 动物科学学院,贵阳 550025
贵州省位于我国西南地区, 省内绵羊养殖历史悠久[1], 其中绵羊主产区有毕节市威宁县、 纳雍县等. 贵州省养殖的绵羊主要为地方品种威宁绵羊、 小尾寒羊、 湖羊等. 虽品种较多, 有适应性能优良、 抗逆性强等特点, 但大多数地方品种生长速度慢、 繁殖能力低, 已然成为制约地方绵羊养殖产业发展的重要因素. 因此, 对省内地方品种进行改良, 培育更加优良的新品系, 加快发展肉羊产业, 对促进现代畜牧业技术进步、 提高农民收入和改善居民肉类消费结构具有重要意义.
微卫星DNA是一类广泛分布于真核及原核生物基因组中的短串联重复序列(1~5 bp), 又称为简单重复序列(Short tandem repeats, STRs)或短串联重复序列(Simple sequence repeat, SSR), 约占真核基因组的5%, 多存在于基因编码区附近, 也存在于基因组中的间隔区及基因内的外显子、 内含子和调控区域. 由于重复单位的重复数量在个体间呈高度变异性且变异数量丰富, 因此微卫星座位的应用非常广泛, 已经发展成为畜牧业经济动物育种领域中常用的分子标记之一[2]. 王婷等[3]在四川省凉山地区6个绵羊群体的遗传多样性研究中, 利用12个微卫星座位计算绵羊群体的基因频率、 有效等位基因数、 杂合度及多态信息含量, 评估了群体内遗传多样度, 进而通过遗传距离聚类图、 群体结构推测图、 主成分分析及群体间分子方差分析等评估群体间遗传关系. 周建设[4]利用聚丙烯酰胺凝胶电泳技术, 对猪BMP7外显子及内含子区域的6个微卫星座位多态性进行检测, 并对它们的遗传变异特性及与猪繁殖性状的相关性进行了分析, 发现STC,STA与猪繁殖性状密切相关. 这些研究为开展贵州省毕节市羊群体微卫星座位遗传多样性与繁殖性的相关分析提供了参考.
羊FecB(Fecundity Booroola,FecB)基因于1989年被命名, 最明显的生理效应是增加卵巢中的卵泡数量和排卵数量. 前人研究发现, 一个拷贝的FecB基因可增加母羊排卵数1.65个[5]. 在表型上, 该基因纯合的Booroola母羊(BB型)平均排卵数能达到4.65个, 极显著高于对照组[2]. 鉴于FecB基因的主要遗传效应是增加排卵数, 因此国内外学者广泛关注FecB基因研究. Davis等[6]在中国湖羊和小尾寒羊两个品种羊基因组中发现FecB基因突变. 我国学者已对很多地方绵羊品种进行了FecB检测, 发现湖羊、 小尾寒羊、 多浪羊和中国美利奴羊也存在FecB基因[7], 同时至少发现9个品种存在FecB突变[8-9].
关于FecB基因突变与羔羊出生质量之间的关系已有部分研究. 然而关于FecB微卫星座位及与贵州省毕节市羊群实际产子数的关联分析尚不清楚. 因此, 本试验通过检测毕节地区6个绵羊品种FecB基因6个微卫星座位的多态性, 分析它们在6种羊品种中的多态分布情况, 并探讨微卫星座位与羊产羔性能的关系, 以期为FecB基因在贵州省毕节市羊品种的分子标记辅助育种中提供理论依据.
1 材料与方法
1.1 材料与试剂
1.1.1 样品
湖羊与威宁绵羊杂交品系(HW)55头、 威宁绵羊(W)30头、 湖羊(HY)30头、 萨福克羊与威宁绵羊杂交品系(SW)20头、 萨福克羊(SFK)30头、 萨福克羊与半细毛羊杂交品系(SB)20头的血液及耳组织样品采自贵州省毕节市威宁县、 赫章县, 并记录其年产羔羊数; 每只羊无菌采集血液和耳组织样品后低温保存带回实验室, -20 ℃保存备用.
1.1.2 引物设计与合成
根据GenBank中的FecB序列, 选择位于绵羊2号染色体FecB区域可能与繁殖性能相关性较大的6个微卫星座位(LSCV043,GC101,471U,Bulge5,BMS2508,300U), 引物由上海生工生物工程技术服务公司合成(表1).

表1 微卫星引物名称及序列
1.2 方法
1.2.1 基因组的提取
耳组织50 mg或者静脉血200 μL用于抽提DNA, 提取方法参照天根血液/组织/细胞DNA提取试剂盒说明书进行. 提取样品DNA后, TE(Tris和EDTA配置而成)溶解, 用琼脂糖凝胶电泳和紫外分光光度计检测DNA的纯度和体积质量分数, 随后-20 ℃保存备用.
1.2.2 PCR反应体系
PCR反应体系为20 μL, 包含Tiangen PCR mix 2×buffer 10 μL, 两侧引物各0.5 μL, DNA模板1 μL, 用ddH2O补足20 μL. PCR反应程序为: 94 ℃预变性5 min, 94 ℃变形30 s, 退火30 s, 72 ℃延伸30 s, 共计30个循环; 再延伸72 ℃ 5 min; 4 ℃保存备用.
1.2.3 聚丙烯酰胺凝胶电泳
扩增产物用8%的变性聚丙烯酰胺凝胶电泳分离, 电泳条件: 在恒压220 V, 1.0×TBE缓冲液中电泳7~10 h, 硝酸银染色法显色后, 用凝胶成像系统拍照并保存.
1.2.4 微卫星检测方法
将染色后的凝胶用凝胶成像系统复制保存, 并利用Kadak Digitial Science ID Image分析软件根据标准PBR322/MspI Maker计算片段大小.
1.3 统计分析
等位基因频率(Allele frequencies): 微卫星座位呈等显性遗传状态, 其等位基因频率根据电泳图谱统计后获得; 利用Cervus 3.0 软件(Cervus公司)统计6个群体不同位点的等位基因数、 期望杂合度、 观测杂合度和多态信息含量. 利用SPSS 22.0软件的线性回归对不同微卫星座位与繁殖性状进行关联分析, 初步确定与繁殖性状存在显著差异的位点,p<0.05为差异具有统计学意义.
2 结果与分析
2.1 6个羊品种微卫星多态位点的等位基因及频率分析
6对微卫星座位引物均能在6个羊群组中稳定扩增. 微卫星座位的等位基因片段数量及大小如表2所示, 其中微卫星座位LSCV043共检测到4个等位基因, 片段大小为114~160 bp; 微卫星座位BMS2508共检测到6个等位基因, 片段大小为150~190 bp; 微卫星座位300U共检测到5个等位基因, 片段大小为148~178 bp; 微卫星座位GC101共检测到4个等位基因, 片段大小为196~238 bp; 微卫星座位Bulge5共检测到4个等位基因, 片段大小为136~160 bp; 微卫星座位471U共检测到3个等位基因, 片段大小为196~204 bp.

表2 不同微卫星座位的等位基因
表3列出了每个微卫星座位在6个群体中的等位基因频率. 所谓优势等位基因, 就是指某特定品种在特定的基因座位上相对集中的等位基因, 优势等位基因集中可能说明该位点为大多数绵羊所共有, 有可能是起源进化最早、 最原始的基因. 从表3中可以看出, 每个位点都占有一定优势的等位基因.

表3 6个微卫星座位在6个绵羊群体中的等位基因频率
各个微卫星座位等位基因频率在不同绵羊群体中具有明显差异, 对6个绵羊群体6个微卫星座位基因频率的统计结果见表3. 结果显示, 在微卫星座位LSCV043中, 等位基因114 bp在6个绵羊群体中的频率最高(10.00%~70.00%), 160 bp出现的频率最低, 且仅在萨福克羊及萨福克与半细毛羊杂交品系中出现; 在微卫星座位BMS2508中, 等位基因162 bp在6个绵羊群体中出现的频率最高(16.67%~60%), 156 bp出现的频率最低, 且未能在湖羊与威宁绵羊杂交品系、 萨福克与威宁绵羊杂交品系及萨福克羊中检测到; 在微卫星座位300U中, 等位基因160 bp在6个绵羊群体中的频率最高(41.67%~50.00%), 168 bp出现的频率最低, 且仅在萨福克羊与威宁绵羊杂交品系中检测到; 在微卫星座位GC101中, 等位基因200 bp在6个绵羊群体中的频率最高(10.00%~50.00%), 238 bp出现的频率最低, 且未能在威宁绵羊品系中检测到; 在微卫星座位Bulge5中, 等位基因136 bp在6个绵羊群体中的频率最高(55.00%~63.33%), 146 bp出现的频率最低, 且未能在萨福克羊品系中检测到; 在微卫星座位471U中, 等位基因200 bp在6个绵羊群体中的频率最高(60.00%~90.00%), 196 bp出现的频率最低, 且未能在威宁绵羊和湖羊品系中检测到.
2.2 6个羊群组微卫星预期杂合度值(He)、 有效等位基因数(Ne)和多态信息含量(PIC)
6个微卫星座位的遗传指标见表4、 表5、 表6. 微卫星座位在群体内多态性信息含量有一定的差异, 各群体有效等位基因数(Ne)的范围在1.7~4.01之间, 其中湖羊与威宁绵羊杂交品系在微卫星座位BMS2508有最高的有效等位基因数量(4.017 9); 萨福克羊在微卫星座位471U有最低的有效等位基因数量(1.219 5), 表明贵州省毕节市绵羊群体存在等位基因差异, 但差异并不明显.

表4 6个微卫星座位在6个羊群体中的有效等位基因数(Ne)

表5 6个微卫星座位在6个羊群体中的香农指数

表6 6个微卫星座位在6个羊群体中的预期杂合度值(He)
6个微卫星位点的香农指数如表5所示, 其数值越大表明群体间多样性越高. 微卫星座位BMS2508在湖羊品系中最高(1.608 9),LSCV043座位在湖羊与威宁绵羊杂交品系中多态性更好(1.074 0),300U座位在萨福克羊与威宁绵羊杂交品系中最高(1.424 1),GC101座位在萨福克羊与半细毛羊杂交品系中最高(1.329 7).Bulge5座位在湖羊品系中最高(1.165 5),471U座位在萨福克羊与半细毛羊杂交品系中最高(0.848 7), 该结果进一步提示各羊群品系在6个微卫星座位间存在多态性差异.
6个微卫星座位的预期杂合度值(He)如表6所示,LSCV043座位在湖羊与威宁绵羊杂交品系中期最高(0.651 1), 在萨福克羊品系中最低(0.48);BMS2508座位在湖羊品系中最高(0.77), 在萨福克羊品系中最低(0.48);300U座位在萨福克和威宁绵羊杂交品系中最高(0.722 2), 在萨福克羊品系中最低(0.62);GC101座位在萨福克与半细毛羊杂交品系中最高(0.722 2), 在萨福克羊与威宁绵羊杂交品系中最低(0.611 1);Bulge5座位在湖羊品系中最高(0.625), 在萨福克羊品系中最低(0.48);471U座位在萨福克羊与半细毛羊品系中最高(0.493 8), 在萨福克羊品系中最低(0.18). 表6结果提示贵州地方品种及杂交后代遗传多态性较高, 而萨福克羊的遗传多态性较低, 基因保守型较好.
用多态信息含量(PIC)可以描述微卫星座位的变异程度. 当PIC>0.5时为高度多态基因座位; 0.25

表7 6个微卫星座位在6个羊群体中的多态信息含量(PIC)
2.3 6个羊群遗传距离分析
遗传距离是指两个群体间的遗传差异(基因组差异), 但也常用等位基因频率的函数来度量遗传差异. 从表8中可知, 湖羊和威宁绵羊杂交品系与湖羊、 威宁绵羊遗传距离较近, 分别为0.908 8和0.923 5; 萨福克羊、 半细毛羊与湖羊、 湖羊和威宁绵羊杂交品系及威宁绵羊等遗传距离较远.
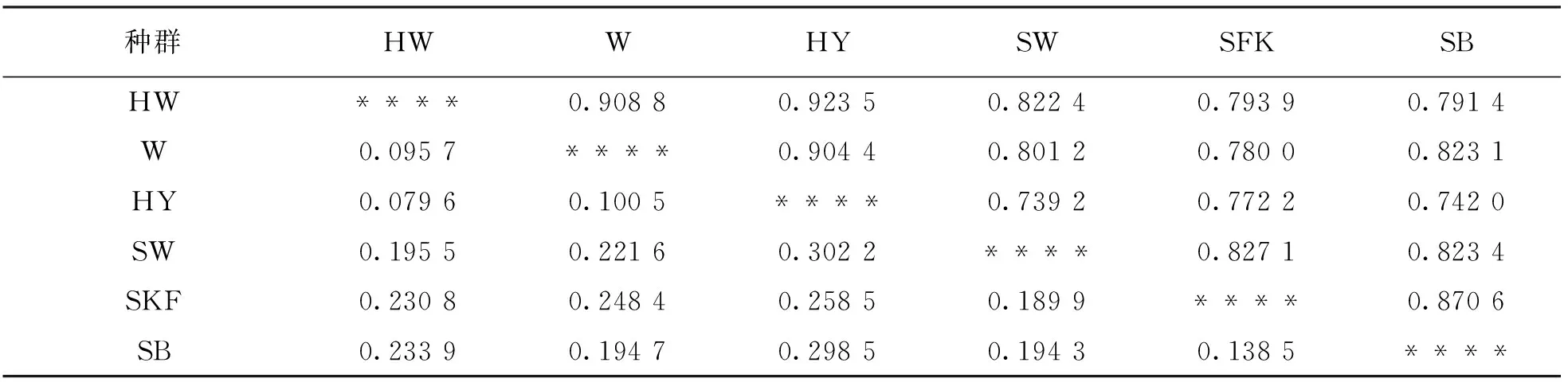
表8 6个种群标准遗传距离结果(Nei)
2.4 微卫星座位300U,GC101的不同基因型与湖羊和威宁绵羊杂交品系(HW)产羔羊数的关系
微卫星座位300U各基因型与湖羊和威宁绵羊杂交品系(HW)产羔羊数的最小二乘平均值±标准误见表9. 从表9中可以看出, 基因型148/160 bp对应的最小二乘平均值最高, 达到了1.81只, 且与其他基因型差异显著(p<0.05), 其次是160/164 bp, 也达到了1.53只, 而基因型160/168 bp对应的最小二乘平均值最低, 为1.17只(p<0.05). 从表10中可以看出, 微卫生座位GC101基因型200/238 bp对应的最小二乘平均值最高, 达到了1.74只, 显著高于196/196 bp、 210/238 bp和200/210 bp(p<0.05), 其次是196/210 bp, 也达到了1.55只, 而基因型200/210 bp对应的最小二乘平均值最低, 为1.27只(p<0.05).

表9 HW微卫星座位300U各基因型与产羔羊数的最小二乘平均值
3 讨论
微卫星DNA广泛分布于真核生物基因组中, 具有随机性、 呈共显性遗传、 遵循孟德尔遗传规律的特点. 同一个体不同年龄段、 不同组织器官、 皮肤、 毛发等产生的DNA指纹图完全一致, 微卫星等位基因的差异主要来自核心区重复单位数目的变化[10]. 本研究涉及的贵州省毕节市6个品种绵羊共检测到13个微卫星等位基因和40种基因型, 其中BMS2508座位、LSCV043座位、Bulge5座位均为中度或高度多态位点, 而471U座位在各组羊群中均为中度和低度多态基因座位, 提示BMS2508座位、LSCV043座位、Bulge5座位可作为筛选羊群基因型的潜在分子标记. 群体多态信息含量(PIC)和预期杂合度值(He)都能反映群体内个体遗传变异的程度, 数值高说明遗传变异大, 反之则群体内的遗传变异小. 在本研究的 6个绵羊群体中湖羊与威宁绵羊、 萨福克与半细毛羊杂交品系的多态信息含量(PIC)、 预期杂合度值(He)较高, 而湖羊、 威宁绵羊、 萨福克羊品系相对较低, 说明贵州省毕节市杂交羊群较本地纯种威宁绵羊的遗传变异大. 近年来, 不断有研究者通过杂交选育手段将FecB基因导入繁殖能力较低的羊群中, 以提高经济效益. 王伟霞等[11]以不含有FecB基因的美利奴羊为父本, 以小尾寒羊公羊与东北细毛羊母羊杂交品系为母本进行杂交, 结果发现FecB基因对提升后代的产羔性能发挥重要作用. 张也等[12]以白头杜泊羊为父本, 以湖羊为母本进行杂交, 结果表明FecB基因对杜湖杂交品系后代母羊的产羔率有影响. 综上所述,FecB基因总体上对绵羊繁殖性能产生良性影响.
FecB基因是绵羊高繁殖力的一个主效基因, 1993年Montgomery等[13]首先报道FecB基因与微卫星基因座位OarAE101和OarHH55紧密连锁, 遗传距离分别为13 cM和20 cM. 等位基因数是群体遗传多样性分析的重要参数之一, 其数量与样本含量和研究涉及的座位数有关. 在基因座位数相同的条件下, 样本量越大, 可检测到的等位基因数越多. 张林等[14]报道,LSCV043微卫星座位98 bp等位基因与小尾寒羊FecB基因B等位基因之间存在一定的连锁不平衡关系, 证明其是与小尾寒羊多羔主效基因紧密连锁的一个遗传标记. 吴舒洁等[15]报道, 小尾寒羊FecB基因B等位基因与471U微卫星座位 200 bp等位基因之间存在一定的连锁不平衡关系. 本研究开展了贵州省毕节市FecB基因微卫星座位多态性和繁殖性状统计, 并就二者的相关性进行了分析. 研究发现, 微卫星座位300U在湖羊与威宁绵羊杂交品系中等位基因数为5个, 其中基因型148/160 bp对应的最小二乘平均值最高, 达到了1.81只; 微卫星座位GC101基因型200/238 bp对应的最小二乘平均值最高, 达到了1.74只, 可用于初步辅助选择, 但这部分基因型个体在群体中的比例并不高, 同时产羔羊数在一定程度上还受到环境、 营养状况和饲养管理水平的影响. 因此, 微卫星座位300U和GC101作为湖羊与威宁绵羊杂交品系繁殖力的分子标记, 尚需要扩大样本进一步研究.
4 结论
本研究检测了6种绵羊群体中FecB基因上6个微卫星座位的多态性,300U和GC101可作为筛选羊群繁殖性状的分子标记, 其中微卫星座位300U基因型148/160 bp, 微卫星座位GC101基因型200/238 bp与湖羊和威宁绵羊杂交品系产羔羊数的关联性最强, 提示其可作为筛选羊群繁殖力的分子标记.
