气候因子影响马铃薯致病疫霉的研究进展
2023-09-26王艳平李希静谢婉莹余佳丽冯媛媛刘松青詹家绥
王艳平 李希静 张 娜 罗 亮 陈 佳 谢婉莹 余佳丽 冯媛媛 刘松青 詹家绥
(1 成都师范学院化学与生命科学学院,特色园艺生物资源开发与利用四川省高校重点实验室,四川成都611130;2 瑞典农业大学自然资源与农业科学学院,瑞典乌普萨拉 75007)
马铃薯作为粮菜饲兼用作物,含有丰富的蛋白质、氨基酸、维生素和矿物质等,不仅能够为人体提供丰富的热量,同时还为人类提供丰富的矿物质(曾凡逵 等,2015;Niekerk et al.,2016;杨庆余等,2021;郑军伟 等,2022)。据联合国粮食及农业组织(FAO)统计数据显示,2021 年我国马铃薯种植面积约为578.27 万hm2,占全球总种植面积的31.89%;总产量约为943.62 万t,占全球总产量的25.09%,种植面积和总产量均居世界第一,但我国马铃薯单产(16 317.90 kg · hm-2)显著低于世界平均水平(20 742.60 kg · hm-2)。这不仅与种植管理模式有关,还与田间病虫害的发生及种植环境息息相关。
由致病疫霉(Phytophthorainfestans)引起的马铃薯晚疫病是世界范围内制约马铃薯生产的主要病害。马铃薯遭受致病疫霉侵染时,病斑首先发生在叶片和块茎部位,其次是茎部。叶片发病时,叶尖或者叶缘大多先出现病斑,受害初期为水浸状褪绿斑,而后在气候适宜时病斑扩展迅速,数小时内即可扩展到叶片一半或全部,最后叶片边缘长出一圈白霉,通常在雨后或有露水的早晨叶片背面的霉层最明显,湿度大时在叶片正面也能观察到霉层。遇到干旱天气,病斑干枯成褐色,质脆并且容易断裂,叶片背面无明显白霉,病斑扩展慢(Vayda 和刘光明,1997;Feng et al.,2021)。致病疫霉亦可经叶脉侵入马铃薯叶柄及茎部,在茎部皮层形成长短不一的褐色条形斑,病斑在潮湿的环境中能够长出稀疏的白色霜状霉(柯莉萍 等,2018)。块茎发病时,病斑呈现褐色或紫褐色,发病部位表面轻微下陷,内部薯肉逐渐坏死。马铃薯晚疫病在我国于1950 年首次暴发,对马铃薯产业造成了巨大损失,之后该病害在国内不断流行,常年造成20%~40%的产量损失,严重年份达50%以上,甚至绝收。马铃薯晚疫病的发生、发展和流行与马铃薯种植季节和地区的环境温度、光照、湿度以及二氧化碳浓度等气候条件密切相关(Phukan,2007;柳玲玲 等,2016;淳俊 等,2018;梅海丽,2018;李长印 等,2020;Wu et al.,2022),了解气候因子对致病疫霉的影响,并据此提出病害防控的相关建议,将有助于我国马铃薯产业发展。本文综述了主要气候因子对马铃薯致病疫霉的影响,以期为晚疫病的综合防治提供参考。
1 马铃薯晚疫病致病疫霉及发病因素
致病疫霉属于藻物界(Chromista)卵菌门(Oomycota) 霜霉目(Peronosporales) 腐霉科(Pythiaceae) 疫霉属(Phytophthora)。2009 年Haas 等首次报道了致病疫霉菌的基因组信息,其基因组大小为220~240 Mb,分别是同属栎树疫霉(Phytophthoraramorum)和大豆疫霉(Phytophthora sojae)的3.98 倍和2.77 倍,是其他主要病原真菌的1.62~11.62 倍(表1)(Dean et al.,2012)。与栎树疫霉和大豆疫霉相比,马铃薯致病疫霉基因组中存在大量重复序列(图1),效应子基因和其他与寄主互作的大量基因均具有很快的进化速度,这可能是其适应性强的关键因素之一(Haas et al.,2009)。
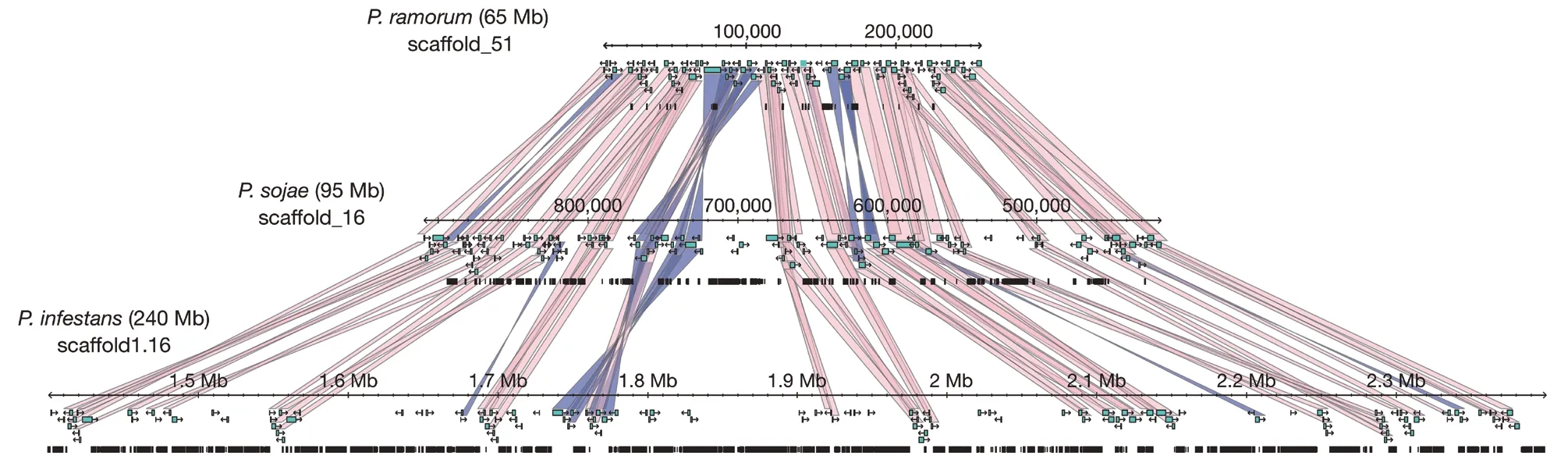
图1 3 个同源疫霉基因支架的保守序列(Haas et al.,2009)
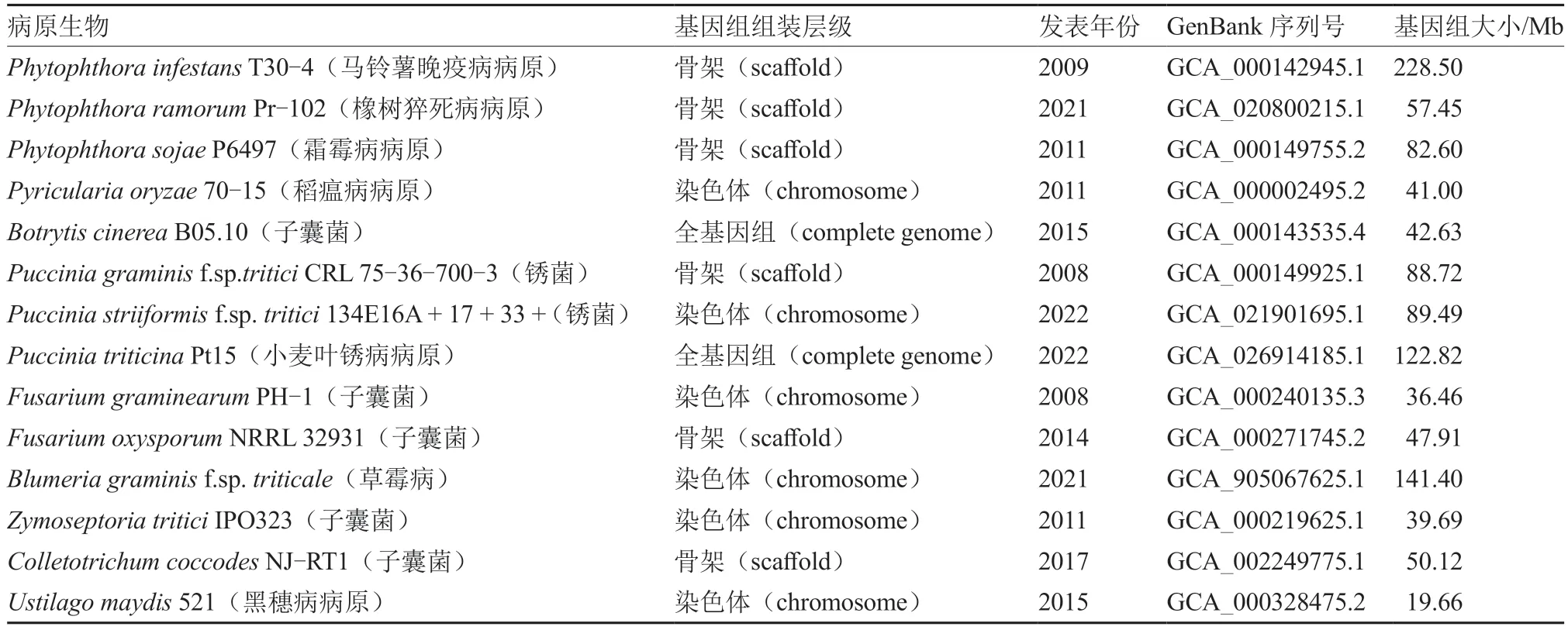
表1 主要卵菌及真菌病原的基因组信息
致病疫霉具有有性生殖和无性生殖两种繁殖方式,无性生殖由菌丝发育形成孢囊梗产生无性孢子,有性生殖是由藏卵器和雄器结合形成卵孢子。卵孢子抗逆性强,可越冬存活,萌发形成初始菌源,但一般卵孢子数量较少,所以晚疫病的危害程度和流行速度与病原菌的再侵染次数和无性孢子数量密切相关。研究表明,生长季节不同,致病疫霉再侵染的次数存在明显差异,在我国云南省大春作马铃薯产区晚疫病每季发生40~130 次再侵染,秋作马铃薯产区晚疫病每季发生25~48 次再侵染,冬作马铃薯产区晚疫病每季发生4~13 次再侵染(李亚红 等,2022)。晚疫病在田间发生后很难快速控制,生产中可采用种植抗病品种、化学防治和生物防治等方法减少损失(刘琼光 等,2010;曲亚英 等,2022),但目前抗病品种较少,国内各马铃薯产区感病品种的种植面积超过70%,若遇适宜气候环境,发病率可达50%~60%(黄冲和刘万才,2016)。化学防治是防治晚疫病最有效的手段,然而已有研究表明,一定范围内的气温升高会使致病疫霉对化学药剂的耐受性增加(Lurwanu et al.,2020,2021)。因此,在初始菌量、气候环境、品种抗性及防治措施中,气候环境是影响马铃薯晚疫病发生和流行不可或缺的因素。温度、光照、湿度以及CO2浓度等气候因子不但能引发非生物性的生理病害,而且是诱导生物性病害侵染和循环、决定生物性病害防控结果的重要因素(图2)。
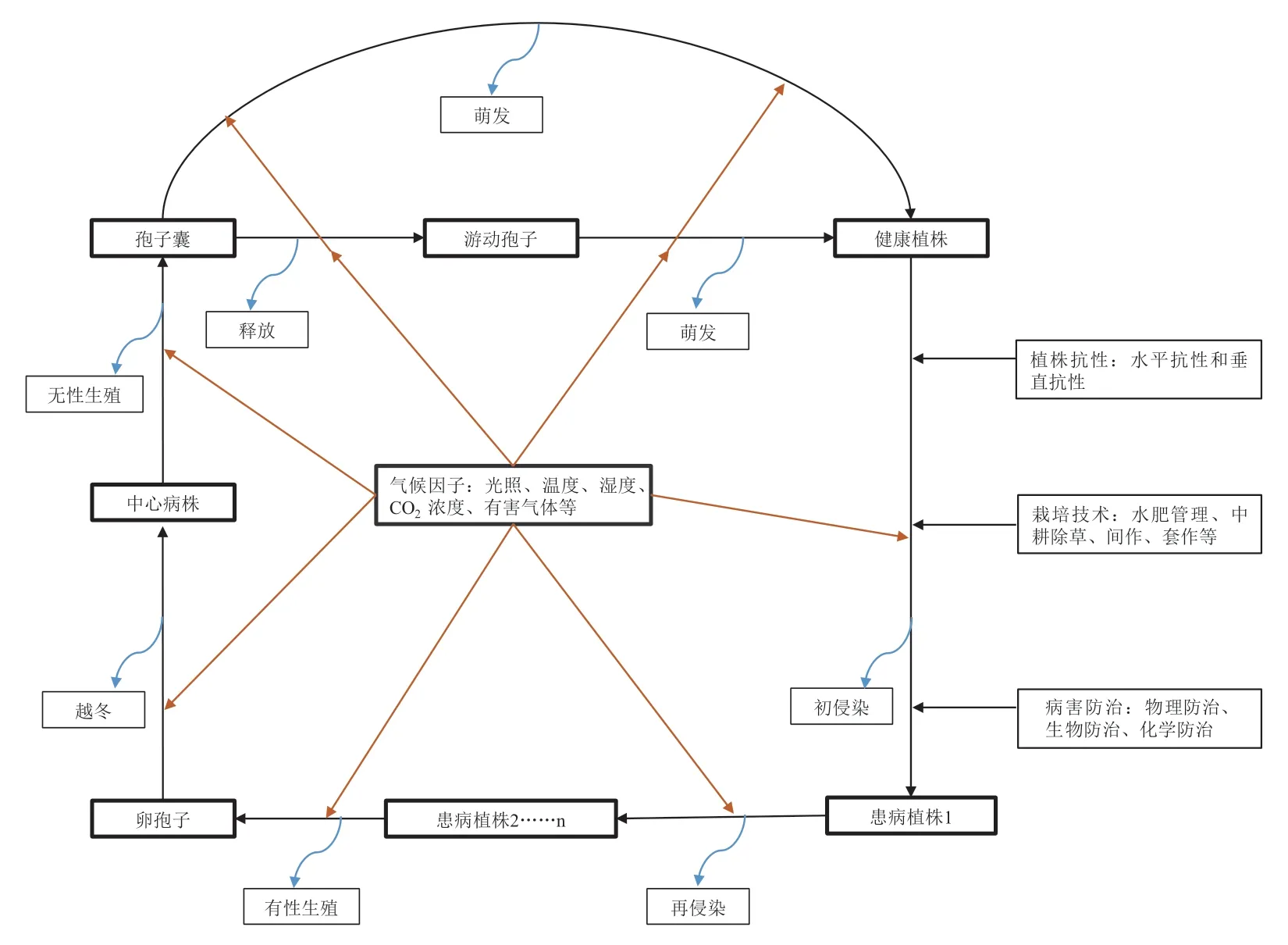
图2 马铃薯致病疫霉为害循环过程
2 温度对马铃薯致病疫霉的影响
温度不仅可以影响致病疫霉的菌丝生长、繁殖模式,而且对其产孢能力、孢子萌发方式、效应子基因的表达、致病力和耐药性等均有较大影响,是影响致病疫霉生存繁殖及其与寄主间互作的关键因素(周阳 等,2014;Eyre et al.,2014)。因此,温度可以很大程度上调控晚疫病的发生与流行(Sparks et al.,2014;胡珍珠,2016)。当前温度对致病疫霉影响的研究主要分为以下几个方面。
2.1 高温对马铃薯致病疫霉的影响
在病害发生和发展过程中,菌丝生长速率和产孢量及孢子萌发对病害的发展均十分重要。贾京珠(2015)分析不同线粒体单倍型致病疫霉在15、18、22、25 ℃和27 ℃ 5 个温度梯度下的菌丝生长率,发现18 ℃下各单倍型菌株的菌丝生长率均最高,随着温度升高菌丝生长速率均降低。Yang 等(2016)对宁夏、甘肃、贵州、云南、福建和广西等6 个马铃薯种植区的140 株致病疫霉株研究发现,致病疫霉丝生长的最适温度为17.69~19.85℃,最高温度为29.33~33.77 ℃。通过测定致病疫霉丝生长速率发现,采自不同地区的菌株对温度的适应性不同,高温培养时来自温暖地区的菌株比寒冷地区的个体明显具有更好的适应性,即致病疫霉的菌丝生长存在明显的本地适应性。Lurwanu 等(2021)对我国内蒙古、福建、宁夏、湖北、广西、甘肃和贵州等地区致病疫霉抗药性机制的相关研究发现,致病疫霉的耐药性和试验温度呈二次回归相关,温度的有限升高可能增加致病疫霉对内吸性杀菌剂嘧菌酯和保护性杀菌剂代森锰锌的抗药性风险,但温度过高则表现为降低致病疫霉对内吸性杀菌剂嘧菌酯和保护性杀菌剂代森锰锌的抗药性风险,然而群体间对农药敏感性的差异与外界气候温度呈直线正相关(Lurwanu et al.,2020,2021)。Wu 等(2020)对内蒙古、福建、宁夏、湖北、广西、甘肃、贵州等地区致病疫霉的研究表明,当温度高于25 ℃时,致病疫霉的菌丝生长变慢,且温度越高这种现象越明显;但当病原菌长期生长于26~29 ℃的高温环境中时,会产生一定的适应性。除了Hsp70、Hsp90 等大量热激蛋白基因参与调控外(表2),致病疫霉还可通过糖代谢、氨基酸代谢、磷酸盐代谢等多种途径来降低高温胁迫造成的自身损伤(Sujkowski,2010;Yang et al.,2016;Shelley et al.,2017)。
致病疫霉的无性繁殖体有两种:孢子囊和游动孢子。在生长发育过程中,菌丝发育形成孢囊梗,孢囊梗顶端膨大形成孢子囊。当田间温度> 15 ℃时,孢子囊可直接萌发,形成芽管侵入寄主植物(Latijnhouwers et al.,2004),而后在寄主细胞的间隙生长,形成极小的丝状吸器伸入寄主细胞内吸取营养(马利松,2003;兰成忠,2009;张小燕等,2021)。Mizubuti 等(2000)研究发现在1.5%水琼脂中,15 ℃培养下致病疫霉单位面积的产孢量较高,孢子囊萌发的速度较快。Andrade-Piedra等(2005)通过田间观察发现,致病疫霉侵入马铃薯引发晚疫病后,周围环境温度在21 ℃左右时孢子囊形成速度最快,在15~21 ℃时单位面积病斑的产孢量最大。20~23 ℃有利于芽管萌发,病害蔓延速度加快;温度继续升高产孢量则下降,当超过26 ℃时较难观察到孢子囊(Harrison,1992;丁海滨 等,2006)。然而,Becktell 等(2005)将致病疫霉接种到番茄上发现,在潜伏期(从接种到病灶形成期间)和病灶形成后,环境温度在23 ℃和28 ℃时孢子囊形成速度最快,18 ℃时单位面积病灶产孢子囊最多。孢子囊在4 ℃低温处理2 h 后再18 ℃培养1 h,可释放大量游动孢子(杨祝强 等,2021)。Mariette 等(2016)对采自北欧、西欧和地中海盆地的致病疫霉在10、14、18 ℃和24 ℃时的致病性研究中发现,10、14、18 ℃条件下其潜伏期逐渐缩短,病斑扩增速率逐渐升高,即致病性不断增强,18 ℃时致病性最强;24 ℃时的致病性较18 ℃略有下降,但显著高于14 ℃时的致病性。在马铃薯现蕾开花期,田间温度较低(< 20 ℃)有利于马铃薯晚疫病的发生,但后期高温有利于病害传播和流行(Adams & Stevenson,1990;Ahmed,2015;池再香 等,2020;张华明,2021)。
2.2 低温对马铃薯致病疫霉的影响
致病疫霉有性生殖产生的卵孢子具有较强的耐低温能力。与4 ℃和21 ℃相比,卵孢子在-20 ℃下保存的活力最好,可以延长休眠期(张铉哲 等,2020),且低温冷冻处理(-20 ℃或-12 ℃)可诱导卵孢子萌发,50%~90%的卵孢子可抵御不良环境、越冬存活,作为下一季田间马铃薯晚疫病的初侵染源(Smirnov et al.,2008;王伟伟 等,2019)。Cohen 等(1997)测定8、11、15、18 ℃和23 ℃下致病疫霉在马铃薯和番茄叶片上形成卵孢子的能力,发现温度越低卵孢子形成能力越强,8 ℃时形成的卵孢子数量最多。杨绪旺等(2005)发现致病疫霉卵孢子形成的最佳温度在10~15 ℃,高于或低于此范围的温度均不利于有性繁殖和卵孢子的形成。温度低于10 ℃时,病原体生长发育缓慢,卵孢子形成延迟。相反,温度高于15 ℃则会促进无性孢子而非有性孢子的产生,从而导致卵孢子数量减少。在4 ℃转至中间温度(5、10、15、18 ℃)再转至18 ℃的变温处理过程中,中间温度为10 ℃有利于卵孢子形成,且卵孢子数量最多。5 ℃和15℃变温处理对卵孢子的影响不大,且卵孢子数量有下降的趋势,变温处理时间以4 h 为佳。笔者比较了140 个致病疫霉在5 个温度梯度(13、16、19、22、25 ℃)下的有性繁殖情况,发现在22 ℃时卵孢子形成最多,13 ℃几乎不形成卵孢子(数据未发表)。不同研究结果存在较大差异可能与使用的试验材料遗传背景有关。由于致病疫霉群体间时空差异很大,不同群体甚至同一群体不同基因型之间有性繁殖的能力也不同(Zhu et al.,2015;Qin et al.,2016;Yang et al.,2016;Wu et al.,2019;Shen et al.,2022;Yang et al.,2022)。在我国南方地区如云南,原有异宗交配为主的致病疫霉群体逐渐被以同宗交配的群体取代(Zhu et al.,2016)。前人研究采用的是同宗交配致病疫霉,而笔者采用的是异宗交配致病疫霉。
2.3 变温对马铃薯致病疫霉的影响
与实验室不同,田间温度会随着季节和日夜交替发生剧烈的变化,所以了解病原菌受变温的影响比受恒温的影响更为重要。Shakya 等(2015)研究7 个恒定温度(10、12、15、17、20、23、27 ℃)和相同平均温度下的昼夜温度变化(变幅 ± 5 ℃和 ± 10 ℃)对致病疫霉侵染率、潜伏期、病斑扩增速率和产孢量的影响,发现侵染率随着低平均温度下温度变化的加快而加快,同时潜伏期随着低平均温度下温度变化的加快而缩短,但在高平均温度下随着温度变化的增加呈相反趋势。与恒定温度相比,小幅度温度变化下致病疫霉侵染率、病斑扩增速率和产孢量升高,而当温度变化幅度较大时则降低。即较小的温度变幅( ± 5 ℃)将增加病害的扩展速度和严重程度,较高的温度变幅( ± 10 ℃)则反之;在低温下变温处理有助于提高侵染率、缩短病害潜伏期,在高温下变温处理则反之。较大的温度变幅( ± 10 ℃)有助于缩短病害的潜伏期,提高发病率(Hossein et al.,2020)。
3 光照对马铃薯致病疫霉的影响
马铃薯是喜光植物,长时间的光照一方面有利于植株生长势健壮、促进块茎生长,另一方面可加快雨、露、雾等天气造成的叶面水膜和水滴的挥发速度,降低环境空气湿度,减轻致病疫霉的危害(张厚桐,2002)。在阴天〔太阳辐射强度(SI)<300 W · m-2〕暴露3 h 致病疫霉的孢子活力略微下降;而在晴天(SI > 600 W · m-2)暴露1 h 可以降低95%致病疫霉的孢子活力,暴露3 h 可显著降低致病疫霉的孢子萌发能力,在其他变量相同的情况下,孢子囊在阴天的存活时间比在晴天长数小时(Mizubuti et al.,2000)。日照时间缩短(< 16 h ·d-1)会加快致病疫霉的无性繁殖,当日照时间不足5 h · d-1时会增加致病疫霉的侵染速率,晚疫病的发病率显著升高(池再香 等,2009)。
在寄主与病原菌的互作中,紫外辐射不仅影响病原菌的侵入、定殖、潜育和致病,还可改变寄主的形态、生理和生化过程,从而影响马铃薯晚疫病的危害程度。紫外辐射还可影响马铃薯的免疫能力和致病疫霉孢子囊的成活率,在紫外辐射处理下,马铃薯由于植株变矮、叶片细胞壁变厚等原因而增强免疫力,而致病疫霉的孢子囊则会很快失去活力,孢子囊萌发率显著降低(Marie et al.,2008)。因此,紫外辐照可降低致病疫霉的侵染速度,延长晚疫病的潜伏期(Braga et al.,2015)。但有研究发现,致病疫霉较强的表型可塑性有利于增加其紫外耐受性,且其十分保守的紫外损伤修复通路(核苷酸切除修复)可保护其遗传DNA 免受紫外损伤(表3)(Andrew et al.,2014;Wu et al.,2019;Wang et al.,2021;Wang et al.,2022)。
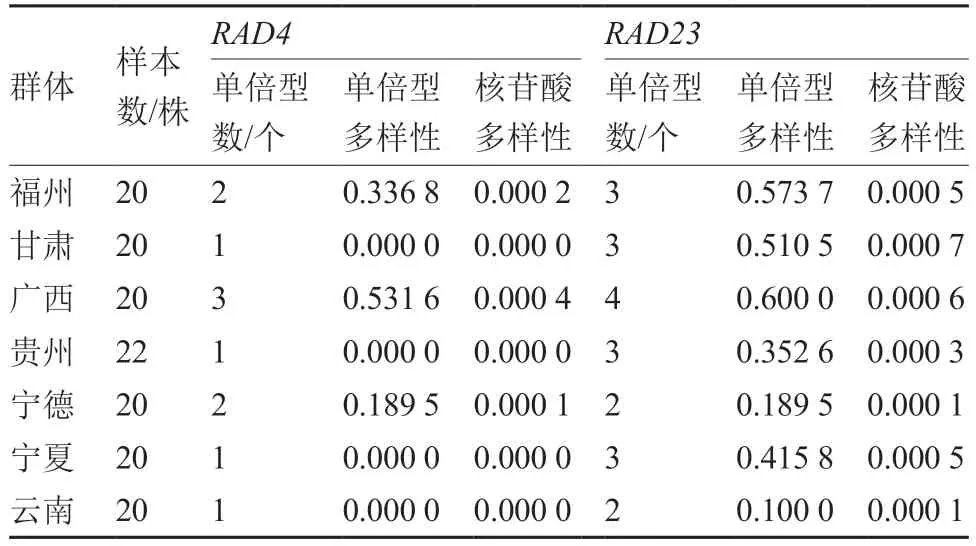
表3 核苷酸切除修复通路关键基因(RAD4 和RAD23)在7 个群体马铃薯致病疫霉中的样本数和遗传多样性
4 湿度对马铃薯致病疫霉的影响
马铃薯晚疫病中心病株的出现与相对湿度和较大降雨量出现的时间密切相关。降雨天气的空气相对湿度大于80%时,马铃薯叶片上易形成水膜和水滴,为致病疫霉的生长和传播提供了有利条件。杨远恒等(2015)研究表明,马铃薯致病疫霉侵染发病的时间与年降雨量最大值出现的时间一致,并且降雨量越大致病疫霉侵染马铃薯的程度就越重。相对湿度和降雨量分别在63%~71%和1.50~3.75 mm 的范围内均有利于马铃薯晚疫病的发生(曹艳秋 等,2008),当田间持续5~7 d 湿度大于85%即可引起晚疫病大发生(刘玉华和张振远,2007)。
由于致病疫霉孢子囊和游动孢子的萌发都必须有水参与(马师祥,2021),所以晚疫病发病期的空气相对湿度通常要达到75%以上,并且致病疫霉的孢囊梗形成要求空气相对湿度在85%~97%范围内,相对湿度越大越有利于马铃薯晚疫病菌的生长繁殖(陈娟娟,2021)。在其他条件适宜时,通常连续3~4 d 空气相对湿度≥ 72%时,10 d 之内就可能在田间发现中心病株。降水量≥ 70 mm、相对湿度≥ 80%的天数超过18 d 时,田间较易发现马铃薯晚疫病的中心病株,持续的高湿度会加快致病疫霉侵染,最终导致马铃薯晚疫病的大面积发生(谭宗九 等,2001)。
5 CO2 浓度对马铃薯致病疫霉的影响
研究发现,植物光合作用能力、水分利用效率、生长和产量、抗病性都受到CO2浓度升高的影响(Mitchell & Zentmyer,1971a)。当CO2浓度升高时,光合作用效率提高,为植物提供碳水化合物,从而使淀粉和糖含量增加(Mitchell & Zentmyer,1971b;Lal et al.,2017)。高浓度CO2使植物叶片数、叶面积、冠层大小和密度增加,从而改变冠层温度和小气候湿度,有利于植物病害的发生(Gautam et al.,2013)。但高浓度CO2又能使植物气孔关闭,从而增加植物抗性、减少病原物入侵率。且部分研究表明,高浓度CO2可以诱导植物产生水杨酸和茉莉酸等物质,从而增加植物对病原菌物和细菌的抗性。
Panke-Buisse 等(2020)研究发现,CO2浓度升高可以抵消内生菌对高羊茅生长和病原菌抗性的有益影响。Kobayashi 等(2006)研究发现,CO2浓度升高水稻感染稻瘟病和叶枯病的风险均会增加。然而Matros 等(2006)研究发现,高浓度CO2使烟草老叶和未感染的幼叶对马铃薯Y 病毒的抗性都有所增加,推测这可能导致寄主植物对病原菌物和病原细菌的易感性增加。Mhamdi 和Noctor(2016)研究发现高浓度CO2会影响易感寄主植物对病原菌的抗性,这与高浓度CO2可诱导植物抗病基因β-碳酸酐酶3(β-CARBONICANHYDRASE 3,βCA3)表达有关。Plessl 等(2007)研究发现,CO2浓度升高降低了由致病疫霉引起的马铃薯叶片坏死率,且致病疫霉孢子形成数量和速率均显著降低,这与CO2浓度升高下叶片中抗病相关酶(β-1,3-葡聚糖酶)的活性升高有关。相反,笔者试验结果揭示,CO2浓度在400~1 200 mg · L-1之间,致病疫霉的致病性、耐药性、抗逆性、无性孢子的形成数量及生长速度都随着CO2浓度的升高而增加(相关数据未发表)。更重要的是,由于人类活动引起的CO2浓度升高很可能会通过调节植物生理特征、营养成分和免疫能力以及其他气候因子如大气温度和湿度间接影响未来晚疫病的流行和防控(Yang et al.,2023)。相较于其他作物气候因素,有关CO2浓度对马铃薯晚疫病发生和流行直接影响的研究很少,未来值得加强。
6 小结
近年来,马铃薯产业不断发展,其在粮食产业上的地位也在不断提升(李国景 等,2023),但致病疫霉仍是影响马铃薯产量的最大威胁之一。温度、光照、湿度等气候因子是影响致病疫霉发生和扩散的重要因素。在未来马铃薯生产和病害防治中,做好致病疫霉群体结构和生长研究对于马铃薯晚疫病防控工作有着重要的意义。据此提出以下建议:首先,采用高垄双行栽培方式,既能够增加光照,又可以避免田间积水,减少因温度低和湿度过大而导致的病害侵染。其次,在马铃薯生长期间增加磷钾肥和微肥用量,尽量避免单一施肥,使植株营养均衡,从而提高马铃薯植株的抗病性,降低马铃薯晚疫病的发生率;加强田间巡查和气候预警,在发现晚疫病病株后立即拔出,装入密封袋中带出田间焚毁,并对病株周围进行灭菌处理,以防止病菌传播侵染其他健康植株。利用环境检测系统,当预测将出现有益于病害大流行的气候条件时,及时告知种植户做好病害防控措施,从而降低危害系数。最后,在马铃薯采收后进行科学轮作,从而达到降低土壤中病原菌数量的目的。同时加强致病疫霉对气候变迁适应性和田间预测预报研究,为有效、持久防控马铃薯晚疫病提供理论基础。
