circERBB2在卵巢癌中的表达及作用机制研究
2023-09-25包利利王晓黎
赵 达, 包利利, 李 波, 俞 岩, 王晓黎
(1. 海南医学院, 海南 海口, 570216; 2. 海南省妇女儿童医学中心 妇科, 海南 海口, 570100)
卵巢癌是临床常见的妇科肿瘤,也是妇科肿瘤相关死亡的5大原因之一[1-2]。早期诊断并治疗可提高卵巢癌患者的生存率,故临床亟需探寻新的诊断与治疗靶点,以改善卵巢癌患者的预后。环状RNA(circRNA)是一种呈封闭环状结构的RNA分子,不受RNA外切酶影响,表达稳定,不易降解[3]。研究[4]表明,由癌症相关染色体易位产生的融合circRNA与肿瘤发生和治疗耐药性有关。微小RNA(miRNAs)是长度为19~22个核苷酸的短链RNA,其作为许多细胞过程的负调节因子,在癌症、糖尿病、肥胖症和心血管疾病等疾病中大量异常表达[5]。miRNAs通过与信使RNA(mRNA)3′UTR的特定位点结合,调节动物30%的基因,导致转录后抑制[6]。miRNAs在细胞增殖、分化、凋亡等方面的作用已得到证实,且其在发育中也发挥作用[7]。研究[8]发现,微小RNA-187-3p(miR-187-3p)在肺癌、肾细胞癌、前列腺癌、结直肠癌中表达下调,提示miR-187-3p在人类癌症中具有抑瘤作用。原癌基因BCL6是转录主调节因子,在分子水平上能够抑制TP53、CDKN1A和ATR等DNA损伤反应基因,以促进与转型重组和体细胞高突变相关的基因组对不稳定性的耐受性。目前,BCL6在卵巢癌中的作用及其与circERBB2和miR-187-3p的关系尚未明确。本研究基于临床样本分析和体内外实验等探讨circERBB2在卵巢癌中的表达情况及作用机制,以期为卵巢癌的预防及靶向治疗提供一定理论依据。
1 材料与方法
1.1 临床样本
选取2021年6月—2022年12月海南省妇女儿童医学中心收治的20例卵巢癌患者的卵巢癌组织和癌旁正常组织作为研究样本,患者均经病理检查和细胞学检查确诊卵巢癌。20例患者中,年龄≤55岁4例, >55岁16例; 肿瘤直径≤3 cm者13例, >3 cm者7例; 国际妇产科联盟(FIGO)分期Ⅰ~Ⅱ期者14例,Ⅲ~Ⅳ期者6例。本研究方案获得海南省妇女儿童医学中心伦理委员会审核批准,且所有患者签署知情同意书。
1.2 细胞培养方法
人正常卵巢细胞系HOSEpiC和人卵巢癌细胞系OVCAR-3、SKOV-3、3AO、OV90均购自北纳生物细胞公司。在含10%胎牛血清(FBS, 美国Hyclone公司)的90% RPMI-1640培养基中培养HOSEpiC、3AO细胞; 在含20%FBS的90% RPMI-1640培养基中培养OVCAR-3细胞; 在含10%FBS的90% DMEM培养基中培养SKOV-3、OV90细胞。所有细胞均放置于37 ℃、5%CO2环境中培养。
1.3 GEO数据库分析
应用findcirc和circfinder工具分析来自GEO数据库的高通量数据(GSE79572数据集)。以P<0.05和倍数变化值>2.0筛选卵巢癌中差异表达的circRNA, 以P<0.05和倍数变化值>1.0筛选差异表达的miRNA。基于Targetscan 7.1软件和韦恩图,筛选GSE79572数据集中差异表达基因与预测所得miRNA靶向基因的交叉基因。
1.4 实时定量聚合酶链反应(qRT-PCR)
使用TRIZOL试剂(美国Invitrogen公司)和PrimeScript RT试剂盒(日本Takara公司)提取总RNA, 使用SYBRGreenI试剂盒(日本Takara公司)和LightCycler2.0试剂盒(瑞士Roche公司)进行qRT-PCR, 内参基因分别为U6和GAPDH, 引物序列见表1, 采用2-△△Ct法分析基因相对表达量。
表1 引物序列
1.5 细胞转染与分组
pcDNA3.1 hsa_circ_0007766(circERBB2)、miR-187-3p模拟物、miR-187-3p抑制剂和阴性对照(NC)载体(中国GenePharma公司),si-circERBB2-1、si-circERBB2-2和si-BCL6(美国Invitrogen公司), si-BCL6、miR-187-3p模拟物和miR-187-3p抑制剂序列见表2。将细胞按不同转染方法进行分组,分别为对照组(转染NC)、si-circ 1组(转染si-circERBB2-1)、si-circ 2组(转染si-circERBB2-2)、模拟物组(转染miR-187-3p模拟物)、si-BCL6组(转染si-BCL6)、抑制剂组(转染miR-187-3p抑制剂)、si-circ+抑制剂组(转染si-circERBB2-1+miR-187-3p抑制剂)、抑制剂+si-BCL6组(转染miR-187-3p抑制剂+si-BCL6)、p-circ组(转染pcDNA3.1 circERBB),根据转染试剂Lipofectamine2000(美国Invitrogen公司)操作说明书进行转染。培养24 h后,检测各组细胞的转染效率。

表2 BCL6 siRNA、miR-187-3p模拟物及抑制剂序列
1.6 核糖核酸酶R(RNase R)消化试验
使用circERBB2和GAPDH在20 μL反应缓冲液中于37 ℃环境进行RNase R消化反应45 min。通过乙醇沉淀法除去酶和盐,重复消化和沉淀反应2次。通过2.0% TAE-Agarose凝胶电泳或2100生物分析仪(Aglient)对已处理RNA样品进行分析,并与未处理RNase R样品进行比较。所有实验重复3次。
1.7 RNA下拉实验
使用生物素RNA标记混合物(瑞士Roche公司)对circERBB2-野生型(circ-Probe组)和对照组(con-Probe组)探针进行体外转录和生物素标记,用不含RNase的DNase I(瑞士Roche公司)处理(Input组)并使用RNeasy Mini试剂盒(美国Qiagen公司)进行RNA下拉实验。将细胞蛋白提取物(1 mg)与生物素化RNA生物素标记的RNA(50 pmol)混合,用链霉亲和素琼脂糖珠(美国Invitrogen公司)孵育,用室温NaCl/碘化丙啶(PI)洗涤3次。将样品离心后,提取RNA上清液,进行qRT-PCR实验,检测circERBB2和miR-187-3p水平。所有实验重复3次。
1.8 双荧光素酶报告基因实验
将野生型BCL6(BCL6-WT)、突变型BCL6(BCL6-MUT)、miR-187-3p模拟物和模拟物对照3′UTR插入pGL3载体(美国Promega公司)合成荧光素酶报告基因载体的相关结合位点。将293T细胞培养于24孔板,再将miR-187-3p模拟物(miR-对照组)、模拟物对照(miR-模拟物组)和BCL6-WT或BCL6-MUT共同转染至293T细胞,通过Lipofectamine3000(美国Life公司)于转染后48 h测定荧光素酶活性。所有实验重复3次。
1.9 细胞活力检测
采用噻唑蓝(MTT)法测定转染后48 h的细胞增殖水平,每个孔用含100.0 μL新鲜0.5 g/L MTT的无血清培养基代替转染培养基。37 ℃孵育4 h后,吸除MTT培养基,并向每孔加入50.0 μL二甲亚砜(DMSO), 37 ℃孵育10 min, 用分光光度仪检测455 nm处吸光度。所有实验重复3次。
1.10 细胞周期检测
在细胞周期实验中,转染后72 h收集不同的细胞群进行消化,得到细胞悬液。将细胞悬液离心,弃上清液,剩余细胞用预冷磷酸盐缓冲液(PBS)洗涤2次,用75%乙醇固定4 h。将固定好的细胞离心,再用PBS洗涤3次。向固定的细胞中加入40 μg PI和1 mL RNase染色液(美国BD生物科学),于避光区室温孵育15 min。染色后,使用FACS Calibur(美国BD公司)检测细胞周期,并使用FACS Diva(美国BD公司)分析数据。所有实验重复3次。
1.11 细胞划痕实验
采用伤口愈合实验评价人卵巢OVCAR-3细胞活力。将细胞置于含无血清培养基的6孔板中,达到80%~90%的融合度,用200 μL微量移液管进行划痕处理。使用显微镜对不同时点划痕愈合情况进行拍照检测。所有实验重复3次。
1.12 Transwell侵袭实验
将100 μL Tritrigel(美国BD Bioscience公司)和400 μL无血清培养基充分混合,置于Transwell腔室(美国Corning公司)中(40 μL/室)。将无血清培养基中0.5×106~2.5×106个细胞放入上腔室(每个腔室200 μL),同时将含20%FBS的培养基添加至下腔室。培养24~48 h后,用4%多聚甲醛固定细胞, 0.1%结晶紫染色。对每个小孔进行拍照,统计5~10个独立区域,计算统计平均值。所有实验重复3次。
1.13 统计学分析

2 结 果
2.1 circERBB2、miR-187-3p与BCL6的关系
本研究对GSE79572数据集进行findcirc和circfinder分析发现,卵巢癌中仅有1种共同的差异表达circRNA呈高表达,即circERBB2, 见图1A; 基于TargetScan Human 7.1发现circERBB2靶向的miRNA,即miR-187-3p, 见图1B; 从TargetScan 7.1中获得GSE79572数据集差异表达基因与预测所得miR-187-3p靶向基因,韦恩图取交集后得到7个交叉基因,见图1C、图1D; 从TargetScan数据库获取circERBB2与miR-187-3p之间、miR-187-3p与BCL6之间的潜在结合位点,见图1E。

A: 对GSE79572数据集进行findcirc和circfinder分析,卵巢癌中仅1种共同的差异表达circRNA(circERBB2)呈高表达;B: TargetScan Human 7.1分析发现靶标miRNA(miR-187-3p); C: GSE79572数据集中差异表达基因与预测所得miR-187-3p靶向基因的韦恩图; D: 7个交叉基因列表; E: circERBB2与miR-187-3p、miR-187-3p与BCL6的潜在结合位点。
2.2 circERBB2、miR-187-3p、BCL6在卵巢癌组织和癌旁组织中的表达情况
卵巢癌组织中circERBB2、BCL6表达水平高于癌旁组织, miR-187-3p表达水平低于癌旁组织,差异有统计学意义(P<0.05), 见表3。卵巢癌细胞系OVCAR-3、SKOV-3、3AO、OV90中circERBB2表达水平均高于正常卵巢细胞系HOSEpiC,差异有统计学意义(P<0.05), 见表4。因OVCAR-3细胞、SKOV-3细胞 circERBB2表达量高,本研究选用OVCAR-3细胞、SKOV-3细胞进行后续实验。

表3 circERBB2、miR-187-3p、BCL6在卵巢癌组织和癌旁组织中的表达情况比较
表4 circERBB2在不同细胞系中的表达情况比较
2.3 circERBB2与miR-187-3p、miR-187-3p与BCL6的靶向关系
RNA下拉实验结果显示,con-Probe组、circ-Probe组circERBB2、miR-187-3p表达水平均低于Input组,差异有统计学意义(P<0.05), 见表5。
表5 RIP下拉实验验证circERBB2和miR-187-3p的表达情况
OVCAR-3细胞、SKOV-3细胞中, p-circ组circERBB2表达水平均高于对照组, miR-187-3p表达水平均低于对照组, si-circ 1组、si-circ 2组circERBB2表达水平均低于对照组,miR-187-3p表达水平均高于对照组,差异有统计学意义(P<0.05), 表明上调circERBB2可以上调OVCAR-3、SKOV-3细胞系中circERBB2表达并下调miR-187-3p表达,沉默circERBB2可以下调OVCAR-3、SKOV-3细胞系中circERBB2表达并上调miR-187-3p表达,见表6。
表6 各组circERRBB2、miR-187-3p表达情况比较
miR-模拟物组BCL6-WT荧光素酶活性低于BCL6-MUT, 差异有统计学意义(P<0.05), 表明miR-187-3p显著抑制了BCL6-WT的荧光素酶活性,但未抑制BCL6-MUT的荧光素酶活性,见表7。
表7 各组荧光素酶活性比较
OVCAR-3细胞、SKOV-3细胞中,模拟物组miR-187-3p表达水平高于对照组,BCL6表达水平低于对照组,抑制剂组miR-187-3p表达水平低于对照组,BCL6表达水平高于对照组,差异有统计学意义(P<0.05), 表明miR-187-3p模拟物和抑制剂可调节miR-187-3p和BCL6表达水平,见表8。
表8 miR-187-3p模拟物和抑制剂对细胞miR-187-3p、BCL6表达的影响
OVCAR-3细胞、SKOV-3细胞中, p-circ组BCL6表达水平高于对照组, si-circ 1组、si-circ 2组BCL6表达水平低于对照组,差异有统计学意义(P<0.05),表明过表达circERBB2可显著上调BCL6表达,沉默circERBB2可显著下调BCL6表达,见表9。
表9 过表达和沉默circERBB2对BCL6表达的影响
2.4 抑制circERBB2、过表达miR-187-3p、抑制BCL6对卵巢癌细胞增殖、侵袭、迁移和细胞周期的影响
OVCAR-3细胞、SKOV-3细胞中, si-circ组、模拟物组、si-BCL6组G0/G1期细胞占比均高于对照组,S期细胞占比均低于对照组,差异有统计学意义(P<0.05); SKOV-3细胞中,si-circ组、模拟物组、si-BCL6组G2/M期细胞占比低于对照组,差异有统计学意义(P<0.05), 见表10。由此表明,抑制circERBB2、过表达miR-187-3p和抑制BCL6均能够抑制卵巢癌细胞增殖。细胞周期实验结果证实, si-circERBB2、miR-187-3p模拟物和si-BCL6阻断了G1期细胞周期,见图2。
表10 各组细胞周期情况 %
划痕实验结果显示, OVCAR-3细胞、SKOV-3细胞中, si-circ组、模拟物组、si-BCL6组的划痕愈合率均低于对照组,差异有统计学意义(P<0.05), si-circ+抑制剂组、抑制剂+si-BCL6组的划痕愈合率与对照组比较,差异无统计学意义(P>0.05), 表明si-circERBB2、miR-187-3p模拟物和si-BCL6可削弱OVCAR-3细胞、SKOV-3细胞迁移能力,而miR-187-3p抑制剂可恢复由si-circERBB2和si-BCL6引起的细胞迁移能力下降,见表11、图3A。
表11 各组OVCAR-3细胞和SKOV-3细胞的划痕愈合率比较 %
Transwell侵袭实验结果显示, OVCAR-3细胞、SKOV-3细胞中, si-circ组、模拟物组、si-BCL6组的侵袭细胞数量均少于对照组,差异有统计学意义(P<0.05), si-circ+抑制剂组、抑制剂+si-BCL6组的侵袭细胞数量与对照组比较,差异无统计学意义(P>0.05), 表明si-circERBB2、miR-187-3p模拟物和si-BCL6可显著降低卵巢癌细胞的侵袭能力,抑制miR-187-3p则可恢复由si-circERBB2和si-BCL6引起的细胞侵袭能力降低,见表12、图3B。
表12 各组OVCAR-3细胞和SKOV-3细胞的侵袭细胞数量比较 个
3 讨 论
卵巢癌是一种多亚型的异质性疾病,其中90%为上皮性肿瘤,间质细胞肿瘤和生殖细胞肿瘤则仅占少数[9-10]。上皮性卵巢癌是最致命的女性生殖系统恶性肿瘤,早期诊断可提高患者的生存率[11-12]。研究[13]表明, circRNA对细胞凋亡、细胞增殖、癌症发生和发育、衰老等过程具有重要的调控作用。在健康组织和病变组织的大多数细胞过程中,一些circRNA能够通过竞争性吸附miRNA而发挥作用[14-15]。本研究基于数据库分析在卵巢癌中发现circERBB2、BCL6和miR-187-3p,并验证了三者间存在的靶向关系,进一步开展各项实验发现, circERBB2可通过靶向miR-187-3p吸附,促进原癌基因BCL6表达,调控卵巢癌的发展。

图2 各处理组流式细胞术检测细胞周期结果
本研究结果显示, circERBB2在卵巢癌中表达较高,且其能通过靶向miR-187-3p发挥调控肿瘤进程的作用。circRNA广泛存在于哺乳动物细胞中,通过吸附miRNA或与其他分子相互作用,在转录时或转录后调节基因表达水平[16]。本研究还发现, circERBB2与miR-187-3p之间,miR-187-3p与BCL6之间存在潜在结合位点。本研究发现, circERBB2和BCL6在卵巢癌细胞系中的表达水平较高, miR-187-3p在卵巢癌细胞系中的表达水平较低。既往研究[17]发现,长链非编码RNA(lncRNA)和circRNA的生物学功能机制使其更倾向于作为上游调节物质调节miRNA表达,进而调控下游基因发挥作用。
本研究发现, circERBB2与miR-187-3p具有靶向关系,且p-circERBB2可以上调circERBB2表达、下调miR-187-3p表达,此外miR-187-3p对BCL6-WT 3′UTR的荧光素酶活性有显著抑制作用,但对BCL6-MUT的荧光素酶活性无显著抑制作用。miRNA主要通过阻断靶基因表达而介导其在动物细胞中的生物学功能,本研究搜索不同数据库后发现miR-187-3p的预测靶标基因是BCL6[18]。本研究还发现, miR-187-3p模拟物和抑制剂可调节miR-187-3p和BCL6表达,而过表达circERBB2可显著上调BCL6表达。由于复杂的结构重排,BCL6基因与盲肌样蛋白1(MBNL1)融合[19]。BCL6在免疫反应中起着重要作用,是炎症的负性调节因子。本研究结果显示, si-circERBB2、miR-187-3p模拟物和si-BCL6可抑制细胞增殖,阻滞G1期细胞周期,抑制细胞迁移能力和侵袭能力。研究[18]显示,肺癌细胞株A549和SPC-A-1中miR-187-3p能够明显抑制细胞增殖及转移进程。此外, miR-187-3p的生长抑制作用除了抑制细胞增殖外,还有诱导细胞凋亡。
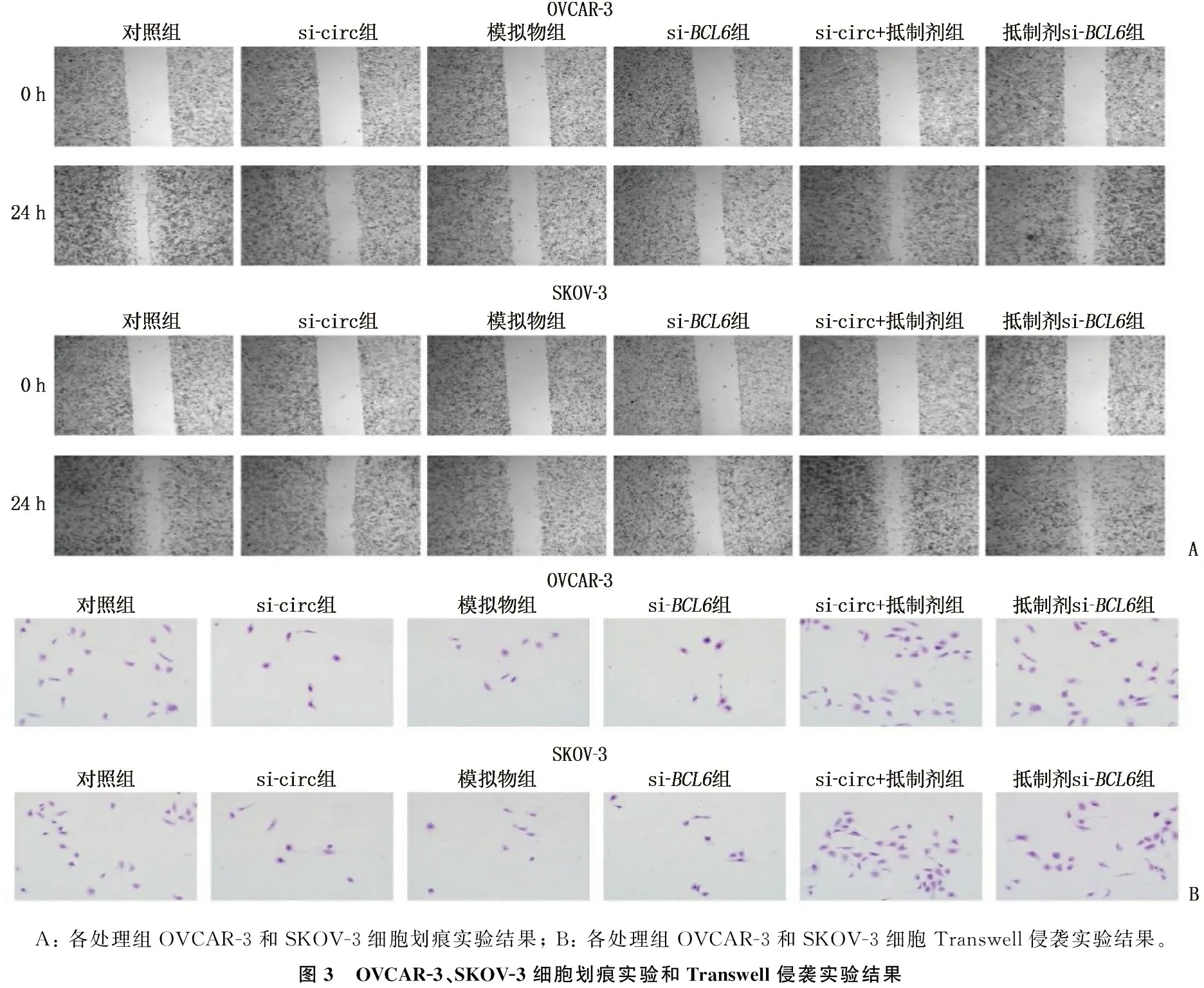
A: 各处理组OVCAR-3和SKOV-3细胞划痕实验结果; B: 各处理组OVCAR-3和SKOV-3细胞Transwell侵袭实验结果。图3 OVCAR-3、SKOV-3细胞划痕实验和Transwell侵袭实验结果
本研究通过细胞实验对circERBB2、BCL6、miR-187-3p三者的调控关系进行初步分析,但仍存在一定局限性,例如仅探讨了circERBB2通过靶向吸附miR-187-3p促进原癌基因BCL6表达进而调节卵巢癌过程,对circBBB2产生这种效应的特殊机制则未深入研究,未来还需进一步深入探讨。
综上所述,本研究基于临床样本分析、细胞实验等发现circERBB2能够通过靶向吸附miR-187-3p促进原癌基因BCL6表达,从而调控卵巢癌过程,这为未来早期诊断标志物的提出及卵巢癌的靶向治疗等提供了一定参考依据。
