黄芩苷对自噬介导H6N6禽流感病毒致病损伤的作用
2023-09-21单春兰宋旭琴欧德渊
杨 鑫,单春兰,杨 剑,宋旭琴,徐 浩,欧德渊*
(1.贵州大学动物科学学院,贵州贵阳 550025;2.贵州省动物疫病与兽医公共卫生重点实验室,贵州贵阳 550025)
禽流感(Avian influenza virus,AIV)是一种高度接触性传染病,常引起家禽和野鸟等发病,发病率和病死率较高,给家禽养殖造成了巨大经济损失[1]。研究发现,H6N6亚型禽流感病毒可在小鼠体内引发肺组织炎症病变,病毒的感染力较强,证实可以跨越物种差异感染哺乳动物[2]。自噬是免疫病原损伤机体的重要机制,当病毒进入细胞后,自噬体和溶酶体融合,促进病毒颗粒的降解,在病毒感染宿主过程中有防御作用[3]。自噬对流感病毒感染机体产生的炎性因子起着重要的调控作用[4]。甲型流感病毒可诱导细胞自噬,自噬参与病毒的复制和发病机制[5]。中药在调控自噬缓解流感病毒所致细胞损伤方面已经取得一定的研究进展[6-7]。黄芩苷是黄芩提取物中有效的黄酮类成分之一,具有抗炎、抗病毒、抗氧化等广泛的药理活性[8]。黄芩苷可以减少H6N6亚型禽流感病毒感染的小鼠炎症细胞的释放,缓解小鼠肺脏的损伤[9]。黄芩苷通过抑制H3N2病毒诱导的自噬阻止病毒的复制,其药理作用尚不明确,自噬与病毒之间的作用机制将成为研究药物治疗流感病毒的新方向[10]。本研究探讨黄芩苷对H6N6亚型禽流感病毒诱导的小鼠肺炎是否通过调控自噬途径来缓解病理损伤,以阐明黄芩苷抗H6N6亚型禽流感病毒潜在的机制,从而为疫病的防控提供理论依据。
1 材料与方法
1.1 材料
1.1.1 试验毒株 病毒株A/duck/Guizhou/03/2016(H6N6),由贵州省动物疫病与兽医公共卫生重点实验室分离、鉴定并保存在-80℃备用。
1.1.2 试验动物 昆明小鼠40只购自长沙市天勤生物技术有限公司,饲养在12 h的明暗交替的环境中,湿度控制为35%~45%,温度控制为21~23℃。
1.1.3 主要试剂 黄芩苷(纯度90%),上海阿拉丁生化科技股份有限公司产品;3-甲基腺嘌呤(3-MA),美国Selleck公司产品;兔多抗LC3,鼠单抗Beclin-1,鼠单抗ATG7,荧光标记山羊抗兔IgG,武汉三鹰生物技术有限公司产品;鼠单抗ATG5,武汉赛维生物科技有限公司产品;β-Actin小鼠单克隆抗体,山羊抗兔IgG(H+L),山羊抗小鼠IgG(H+L),BCA蛋白浓度测定试剂盒,极超敏ECL化学发光试剂盒,上海碧云天生物科技有限公司产品;免疫组化通用试剂盒,福州迈新生物科技有限公司产品;PrimeScriptTM RT Reagent Kit with gDNA Eraser试剂盒,TB Green® Premix Ex TaqTMⅡ试剂盒,TaKaRa公司产品;小鼠白介素-1β(IL-1β),小鼠白介素-2(IL-2),小鼠白介素-6(IL-6),小鼠肿瘤坏死因子-α(TNF-α),上海酶联生物科技有限公司产品。
1.2 方法
1.2.1 试验分组与处理 选用40只昆明小鼠随机分为对照组(control)、模型组(Model)、黄芩苷治疗组(Baicalin treatmentgroup)、3-MA抑制剂组(3-MA inhibitor group)。参照文献建立H6N6诱导小鼠肺炎模型,通过尾静脉攻毒(0.2 mL/只),攻毒3 d后黄芩苷组进行灌胃给药,黄芩苷给药剂量为每天0.056 mg/只[11],3-MA通过腹腔注射,药剂量为每天15 mg/kg[12],各组每天给予等体积灭菌生理盐水,连续治疗6 d后眼球采血后脊椎处死。该试验严格按照动物福利原则进行,并获得了贵州大学动物伦理委员会的批准(批准号EAE-GZU-2021-T096)。
1.2.2 HE染色观察肺脏组织损伤 小鼠左侧肺组织在室温下用40 mL/L多聚甲醛固定72 h,脱水后将组织包埋制作石蜡连续切片,并进行HE染色封片,在显微镜下观察病理损伤情况。
1.2.3 ELISA法检测各组血清中IL-1β、IL-2、IL-6和TNF-α含量 眼球采血收集各组小鼠血清,充分离心(3000 r/min)收集上清,并严格按照试剂盒说明书进行检测。
1.2.4 小鼠肺脏细胞超微结构观察 小鼠处死后将小鼠的肺脏切成厚度不超过4 mm,在1 min内迅速侵入25 mL/L戊二醛中固定4 h,10 g/L四氧化锇再固定,依次进行丙酮逐级脱水、渗透和包埋、超薄切片,先用醋酸铀染色10~15 min,再用柠檬酸铅染色1~2 min,室温下染色。通过透射电镜对片子进行图像采集。
1.2.5 Western blot法检测肺组织LC3、Beclin-1和ATG7蛋白水平 各组称取等量的小鼠右侧部分肺组织,用BCA试剂盒提取总蛋白并定量,将蛋白煮沸10 min后进行SDS聚丙烯酰胺凝胶电泳,将蛋白条带转移到0.22 μm硝化纤维素滤膜上,50 g/L脱脂奶粉封闭,摇床慢摇2 h,经过TBST连续清洗后分别加一抗LC3、Beclin-1及ATG7和β-Actin抗体4℃孵育过夜,加入相应山羊抗小鼠或抗兔的二抗一起孵育2 h,用ECL溶液化学发光对PVDF膜显色拍照。
1.2.6 免疫荧光检测LC3蛋白水平 取各组左侧肺组织制作石蜡切片,依次进行脱蜡、梯度乙醇复水,修复,血清室温封闭30 min,除去封闭液,滴加LC3一抗工作液4℃孵育过夜,后滴加荧光标记的驴抗兔二抗,室温下孵育避光孵育1 h,DAPI复染细胞核5 min。在荧光显微镜下观察并采集图像。
1.2.7 免疫组织化学法检测LC3、Beclin-1和ATG5蛋白水平 取各组左侧肺组织制作石蜡切片,依次进行脱蜡、梯度乙醇复水,修复,血清室温封闭1 h,分别滴加稀释好的一抗LC3II、Beclin-1和ATG5抗体4℃孵育过夜,后滴加稀释好的山羊抗兔或山羊抗鼠二抗,室温孵育1 h,滴加DAB显色(5 min)。封片后于显微镜下观察并采集图像。
1.2.8 荧光定量PCR检测自噬关键基因mRNA转录表达水平 将各组小鼠右侧部分肺组织液氮研磨至匀浆,采用Trizol法提取肺组织总RNA,依次按PrimeScriptTMRT reagent Kit with gDNA Eraser和TB Green®Premix ExTaqTMⅡ说明书进行反转录与实时荧光定量PCR反应:95℃ 30 s,95℃ 5 s,55~59℃ 20 s,共35个循环。以β-actin作为内参,采用相对定量2-△△Ct法比较LC3、Beclin-1、ATG3、ATG5、ATG7及ATG12 mRNA的相对表达量。查阅GenBank并设计相关引物(表1),引物均由上海生工生物工程有限公司合成。
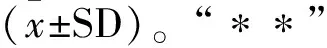
2 结果
2.1 黄芩苷对小鼠肺组织学的影响
对照组小鼠肺泡结构清晰,没有出现炎症性病理变化(图1),与对照组比较,模型组小鼠小支气管周围炎症细胞浸润,肺泡壁毛细血管壁充血,间质变宽,证明H6N6感染的小鼠急性肺损伤造模成功。与模型组相比,黄芩苷治疗组与3-MA抑制剂组可以显著缓解由H6N6亚型禽流感病毒所致的病理损伤,炎性细胞浸润减少。

A.对照组;B.模型组;C.黄芩苷治疗组;D.3-MA抑制剂组A.Control group; B.Model group; C.Baicalin treatment group; D.3-MA inhibitor group
2.2 黄芩苷对小鼠血清中IL-1β、IL-2、IL-6和TNF-α含量的影响
与对照组相比,模型组IL-1β、IL-2、IL-6和TNF-α炎性因子含量显著升高(P<0.01);与模型组比较,黄芩苷治疗组IL-1β、IL-2、IL-6和TNF-α炎性因子含量显著降低(P<0.01);3-MA抑制剂组的IL-1β、IL-2、IL-6和TNF-α炎性因子在也显著降低(P<0.01)(图2)。
A.IL-1β的含量;B.IL-2的含量;C.IL-6的含量;D.TNF-α的含量A.IL-1β content; B.IL-2 content; C.IL-6 content; D.TNF-α content1.对照组;2.模型组;3.黄芩苷治疗组;4.3-MA抑制剂组1.Control group; 2.Model group; 3.Baicalin treatment group; 4.3-MA inhibitor group
2.3 黄芩苷对小鼠肺组织自噬体的影响
透射电镜观察各组小鼠Ⅱ型肺泡上皮细胞,对照组Ⅱ型肺泡上皮细胞形态结构较正常,线粒体和嗜锇性板层小体结构完整清晰(图3);与对照组相比,模型组线粒体发生肿胀(嵴断裂溶解),部分嗜锇性板层小体变性,胞浆内有较多自噬体;与模型组相比;黄芩组肺泡上皮细胞的少数线粒体有轻度肿胀,部分嗜锇性板层小体变性,自噬体减少;3-MA抑制剂组Ⅱ型肺泡上皮细胞的少数线粒体出现轻度肿胀,少数嗜锇性板层小体变性,胞浆内有少量自噬。
2.4 黄芩苷对小鼠肺组织LC3II/I、Beclin-1和ATG7蛋白表达水平的影响
与对照组比较,模型组LC3II/I、Beclin-1及ATG7蛋白水平显著升高(P<0.01);与模型组比较,黄芩苷治疗组LC3II/I、Beclin-1及ATG7蛋白水平显著下调(P<0.01);3-MA抑制剂能够显著下调LC3II/I、Beclin-1及ATG7蛋白水平(P<0.01)(图4)。
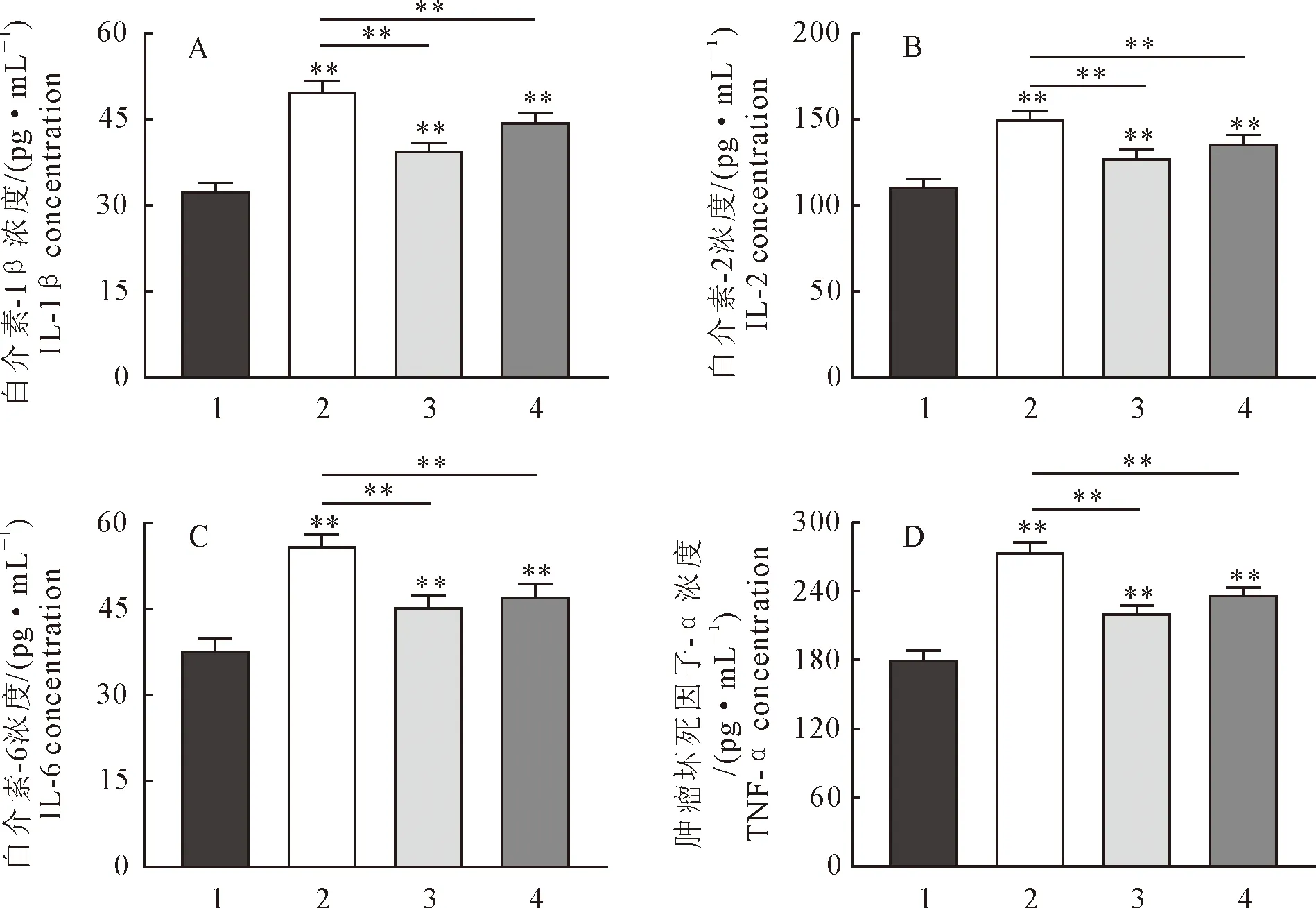
2.5 黄芩苷对小鼠肺组织LC3蛋白表达水平的影响
免疫荧光检测LC3蛋白表达水平结果见图5。与对照组相比,模型组荧光表达强度显著升高(P<0.01);与模型组相比,黄芩苷治疗组的荧光表达强度显著降低(P<0.01);同时3-MA抑制剂组的荧光表达显著降低(P<0.01);此结果与Western blot结果一致。
A.免疫荧光检测LC3蛋白结果;B.LC3蛋白量化结果A.LC3 protein detected by immunofluorescence; B.Quantification results of LC3 protein1.对照组;2.模型组;3.黄芩苷治疗组;4.3-MA抑制剂组1.Control group; 2.Model group; 3.Baicalin treatment group; 4.3-MA inhibitor group
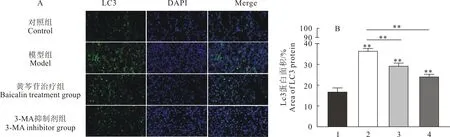
2.6 黄芩苷对小鼠肺组织LC3、Beclin-1和ATG5蛋白表达水平影响
免疫组化检测LC3、Beclin-1及ATG5蛋白结果见图6。与对照组相比,模型组LC3、Beclin-1及ATG5阳性表达升高(P<0.01);与模型组相比,黄芩苷治疗组的LC3、Beclin-1及ATG5阳性表达降低(P<0.01);同时3-MA抑制剂组的LC3、Beclin-1及ATG5阳性表达降低(P<0.01)。
A.免疫组化检测LC3、Beclin-1及ATG5蛋白表达水平;B.LC3蛋白区域量化结果;C.Beclin-1蛋白区域量化结果;E.ATG5蛋白区域量化结果
2.7 黄芩苷对小鼠肺组织自噬关键基因mRNA转录表达水平的影响
RT-qPCR检测自噬关键基因结果见图7,与对照组比较,模型组LC3、Beclin-1、ATG3、ATG5、ATG7及ATG12 mRNA转录水平均不同程度的升高(P<0.01);与模型组相比,黄芩苷组LC3、Beclin-1、ATG3、ATG5、ATG7和ATG12 mRNA转录水平均不同程度下降(P<0.01),同时3-MA抑制剂组的LC3、Beclin-1、ATG12及ATG5 mRNA转录水平均不同程度下调(P<0.01),黄芩苷与3-MA抑制剂作用呈相同调控趋势。
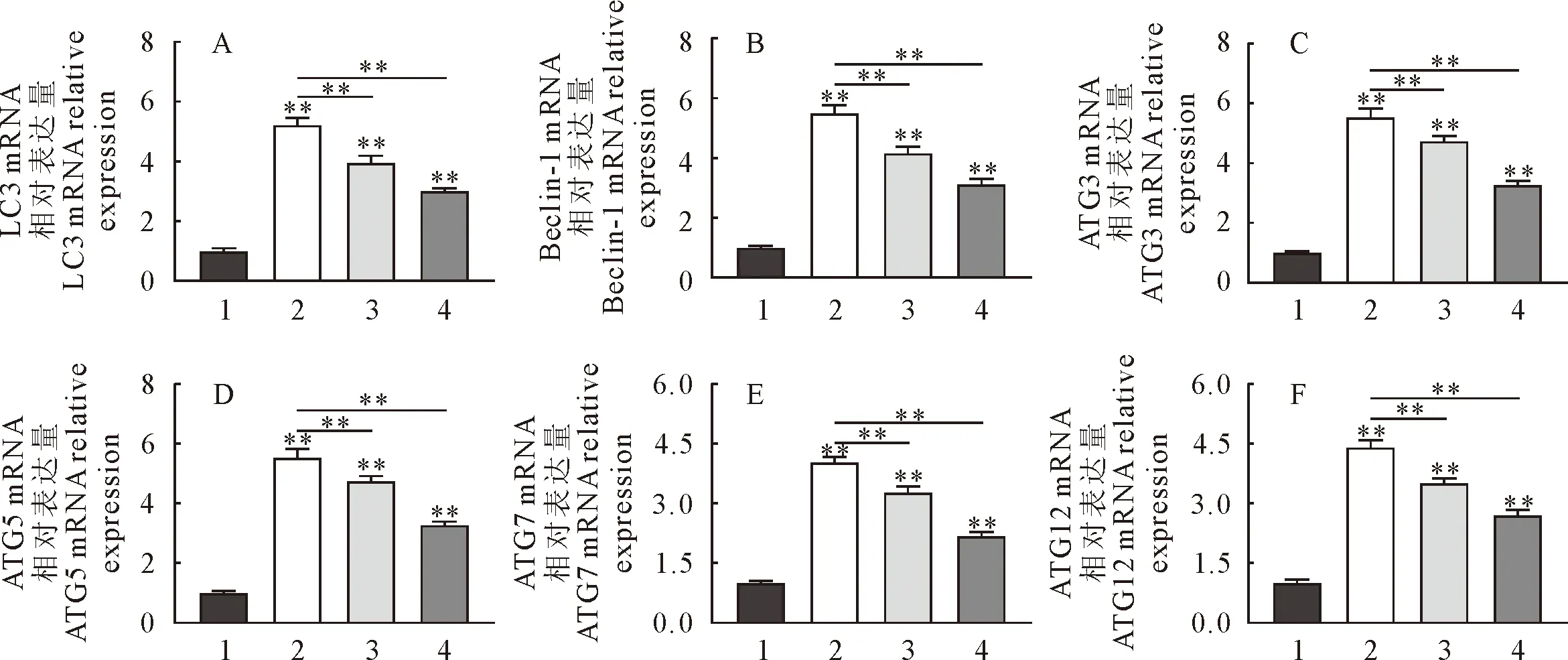
A.LC3 mRNA转录水平;B.Beclin-1 mRNA转录水平;C.ATG3 mRNA转录水平;D.ATG5 mRNA转录水平E.ATG7 mRNA转录水平;F.ATG12 mRNA转录水平
3 讨论
H6N6亚型禽流感病毒可跨越物种屏障感染小鼠和猪等哺乳动物,在家禽中的分离率较高,易与其他亚型流感病毒发生重组,对人类健康构成威胁[13]。流感病毒感染气道上皮细胞之后会释放大量趋化因子,将大量的白细胞募集到感染部位来抵御病毒感染。免疫反应对于清除外来病原体和被感染的细胞是必不可少的,但过度的免疫反应会导致大量炎性细胞因子的累积从而导致肺脏、心脏和肾脏等重要器官的损伤[14-15]。当流感病毒感染机体后,机体内炎性细胞因子过度释放,导致机体细胞及器官组织的炎症损伤,这也是肺炎性实变的原因之一[16-17]。此外,细胞自噬在机体应对外来病原微生物感染的免疫应答中有重要的作用[18]。细胞自噬可以介导流感病毒诱导的炎症反应,在诱导自噬时会产生炎性因子调节细胞自噬[19]。研究表明,黄芩苷可以抑制H1N1流感病毒感染小鼠IL-1β、IL-2、IL-6和TNF-α的分泌,有效的减轻小鼠肺组织的病理损伤[20-21]。抑制细胞自噬可以降低H9N2流感病毒诱导小鼠肺脏病理损伤,降低肺组织炎性细胞的释放及降低IL-1β、TNF-α、IL-8的mRNA转录水平[12]。本研究表明,黄芩苷能够减缓由H6N6亚型禽流感病毒诱导小鼠肺脏的病理损伤及其降低血清中IL-1β、IL-2、IL-6和TNF-α含量,与万巧凤等发现的黄芩苷能减轻小鼠肺内的炎性病变的结果一致[22]。说明黄芩苷能够缓解H6N6亚型禽流感感染小鼠肺组织损伤。在3-MA抑制剂的作用下H6N6亚型禽流感感染小鼠血清中IL-1β、IL-2、IL-6和TNF-α的含量降低。提示黄芩苷改善小鼠肺组织的病理损可能与3-MA抑制自噬的作用机制有关。
不同亚型(H5N1和H9N2)的流感病毒可诱导细胞自噬[23-24]。LC3是唯一在成熟过程中与自噬体保持稳定相关的蛋白,与自噬小体和自噬溶酶体相对特异性相关,LC3的活性的高低是衡量自噬小体的数量的金标准,同时在透射电镜下观察自噬小体和自噬溶酶体也是衡量自噬水平的金标准[25-26]。Beclin-1基因可促进溶酶体和自噬体的融合,是自噬蛋白的标志蛋白,对自噬的活化程度有决定性的作用,ATG家族的基因在自噬泡中的位置与活化程度同样受到Beclin-1基因的调控[27]。ATG5、ATG7和ATG12在自噬小体的延伸过程中起重要作用,ATG7启动ATG5-ATG12结合并加速LC3-Ⅰ脂化反应生成LC3-Ⅱ[27-28]。研究发现,黄芩苷通过下调ATG5、ATG7、ATG12、LC3-Ⅱ、Beclin-1 mRNA和蛋白表达水平抑制LPS诱导的RSC-364细胞的自噬[29]。黄芩苷能降低ATG5-ATG12复合物和LC3-Ⅱ的表达抑制H3N2流感病毒诱导的细胞自噬[10]。本试验结果显示,黄芩苷能降低小鼠肺脏组织中自噬体的数量以及降低LC3、Beclin-1、ATG5和ATG7蛋白表达水平,说明黄芩苷能够抑制H6N6禽流感病毒诱导的细胞自噬,与上述研究结果一致。进一步用RT-qPCR检测自噬关键基因验证,发现LC3、Beclin-1、ATG3、ATG5、ATG7和ATG12 mRNA转录水平显著下降。黄芩苷干预后与3-MA抑制剂效果呈相同调控趋势,提示黄芩苷可能通过抑制H6N6亚型禽流感病毒诱导的细胞自噬以缓解小鼠肺损伤。
综上所述,黄芩苷对H6N6亚型流感病毒致小鼠肺损伤的保护作用可能与抑制细胞自噬有关。明确黄芩苷通过抑制H6N6亚型禽流感病毒触发的自噬和炎症反应之间的关系,可能是治疗流感病毒引起的炎症新的靶向目标,有助于进一步阐释黄芩苷抗流感病毒的药理特性。
