‘沙糖橘’S蛋白同源基因CrSPH的克隆与表达分析
2023-09-21吴秀兰唐文武徐呈祥李桂花
吴秀兰,唐文武,徐呈祥,李桂花
(1 肇庆学院 食品与制药工程学院,广东肇庆 526061;2 肇庆学院 生命科学学院,广东肇庆 526061;3. 广东省农业科学院蔬菜研究所,广州 510640)
在显花植物中,自交不亲和性(self-incompatibility,SI)被认为是防止自花授粉、促进异花授粉及产生新基因型的重要遗传机制。SI是由1个单一的复等位基因S位点所控制,以实现接受还是排斥自我的花粉[1]。SI系统可分为孢子体SI(sporophytic SI,SSI)和配子体SI(gametophytic SI,GSI)。SSI和GSI由不同的S位点基因控制,以十字花科为代表的SSI 主要由S位点的SRK(S-receptor kinase)和SCR(S-locus protein 11)基因控制[2]。这2个紧密连锁的S位点基因在柱头的乳突细胞中特异性表达,并在SI中控制柱头的功能[3]。以茄科为代表的GSI主要由雌蕊中的S-RNase和花粉中SLF(S-locus F-box)/SFB(S-haplotype-spectific F-box)基因控制[4]。有关S-RNase研究较多,玄参科[5]、蔷薇科[6]、芸香科[7]、在茄科[8]等均有报道,不同S-RNase之间存在高度的多态性,其氨基酸序列的同源性介于38%~98%之间[9]。在花粉S位点基因研究方面,Lai等[10]从金鱼草的BAC文库中筛选到1个AhSLF-S2基因,该基因与S2-RNase紧密连锁。Sassa等[11]在梨中鉴定了5个F-box基因,均位于Sn-RNase基因附近。Beecher等[12]报道除了关键S因子外,其他非关键因子也参与自交不亲和反应,如PCD[13]、SSK1[14]、CrSKP1-e[15]、BrCML49[16]等。
柑橘属于S-RNase介导的GSI,由花柱S-RNase和花粉SLF相互作用,通过抑制花粉管的生长实现自交不亲和性[17]。当花柱S-RNase进入花粉管后,非自我S-RNase会被SLF识别并被泛素化及降解,花粉管能正常延伸至子房受精。当自我S-RNase释放到花粉管中,会降解花粉管中的RNA,使得花粉管无法在花柱中生长,表现出自交不亲和反应[18]。沙糖橘是中国重要的柑橘品种,属于自交亲和品种。‘无籽沙糖橘’是‘沙糖橘’的芽变体,属于GSI[19-20],是研究自交不亲和的理想材料。课题组前期从无籽沙糖橘SSH文库中筛选到1个自交不亲和相关的EST序列,序列比对为S-蛋白同源基因(S-protein homologous gene,SPH),柑橘SPH基因的相关研究尚未见报道。本研究以甜橙(Citrussinensis)的SPH基因序列(Cs5g34580.1)为参照,克隆了‘沙糖橘’CrSPH基因的CDS和DNA序列,并分析其在花器官不同部位、授粉后不同时段的表达量,同时研究该蛋白对‘无籽沙糖橘’、‘沙糖橘’花粉萌发率的影响,以期为研究CrSPH基因功能及在柑橘自交不亲和反应中的作用机制提供依据。
1 材料和方法
1.1 试验材料
试验用‘无籽沙糖橘’及‘沙糖橘’果树由肇庆市德庆县马圩镇果园提供,为7年生果树。于2019年春季收集含苞待放花蕾,用镊子剥离出花瓣、花丝、柱头、花柱、子房、花丝和花药,液氮速冻后置于-80 ℃冰箱,用于不同组织表达分析。收集自交(‘无籽沙糖橘’ב无籽沙糖橘’)、异交(‘无籽沙糖橘’ב沙糖橘’)后0,1,2,3,4,5,6,7 d的雌蕊,液氮速冻后置于-80 ℃冰箱,用于授粉后不同时段的表达分析。收集‘无籽沙糖橘’和‘沙糖橘’花粉,置于-20 ℃冰箱保存,用于体外花粉萌发实验。
1.2 试验方法
1.2.1CrSPH基因克隆
采用RNA提取试剂盒提取总RNA,采用CTAB法[21]提取基因组DNA,采用Life Technology公司生产的M-MLV反转录试剂盒合成cDNA。参考甜橙SPH基因序列(Cs5g34580.1)设计1对包含起始密码子和终止密码子的引物CrSPH-F/R(表1),扩增CrSPH基因的CDS和DNA序列,扩增产物经1.2%琼脂糖凝胶电泳并回收,获得目的基因片段。将目的基因与pMD19-T克隆载体连接并转化感受态细胞DH5α,经37 ℃过夜培养后,挑取3个阳性菌落经PCR鉴定正确后,送基因测序公司测序验证。

表1 所用引物序列
利用 NCBI的blastp 对CrSPH基因序列和蛋白序列进行分析,采用ProtParam在线工具分析蛋白质的等电点及分子量。利用在线软件SignalP 6.0对CrSPH蛋白进行信号肽分析。
1.2.2 qPCR表达分析
为分析CrSPH基因在花器官的不同部位及授粉后不同时段的表达量,利用qPCR技术检测花瓣、子房、花柱、柱头、花丝和花药等花器官,以及自交(‘无籽沙糖橘’ב无籽沙糖橘’)、异交(‘无籽沙糖橘’ב沙糖橘’)授粉后不同时段的基因表达量。根据CrSPH基因和柑橘Actin基因序列,分别设计符合qPCR要求的特异引物(qCrSPH-F/R)和内参引物(Actin-F/R)(表1),按照qPCR试剂盒(SYBR Green)说明书进行qPCR反应,反应体系为20.0 μL,包括10.0 μL SYBR ExTaq,10 μmol/L引物各1.0 μL,6.0 μL ddH2O和2.0 μL(80 ng) cDNA,扩增反应在ABI 7500实时定量PCR仪进行。设置3次重复,每次用ddH2O做阴性对照,数据分析采用2-ΔΔCT法计算[22]。
1.2.3 Southern杂交分析
以10.0 μg基因组DNA为模板,分别用限制性内切酶EcoR Ⅰ、ScaⅠ、ClaⅠ于37 ℃过夜消化,全部酶切产物于0.8%琼脂糖凝胶电泳3~5 h,待全部产物分离后凝胶成像系统拍照,将其转入Hybond N+尼龙杂交膜。根据PCR DIG探针合成试剂盒说明书合成带有DIG标记的探针,于42 ℃预杂交2 h后,加入制作好的探针过夜杂交;然后用2×柠檬酸钠(stand saline citrate,SSC)溶液(含0.1% SDS),再用0.5×SSC(含0.1% SDS)于65 ℃洗脱2次,每次洗脱10 min;最后于暗室中将X-ray胶片平铺到杂交膜并置于暗匣中,于37 ℃曝光5~30 min。
1.2.4 原核表达载体构建及CrSPH蛋白诱导
设计含有BamH I和SalI酶切位点的引物pET32a-CrSPH-F/R(表1),以pMD19-CrSPH为模板扩增目的基因,获得带有黏性末端的目的基因。用BamH I和SalI双酶切线性化原核表达载体pET32a,将带有黏性末端的目的基因与线性化的表达载体用T4DNA酶连接,经转化、鉴定后获得pET32a-CrSPH重组原核表达载体。将上述重组质粒转化至大肠表达菌株BL21(DE3),以空质粒为对照。用1 mmol/L IPTG诱导菌株表达获得pET32a-CrSPH融合蛋白,并经12% SDS聚丙烯酰胺凝胶电泳。
1.2.5 花粉萌发实验
将原核诱导的CrSPH蛋白稀释成0.4,0.8,1.6,3.2,6.4 μg/μL 5个浓度。花粉培养基配制按照Miao等[26]的方法,分别取上述各浓度的‘无籽沙糖橘’和‘沙糖橘’CrSPH蛋白200 μL,加入到上述花粉培养基后分成2份:一份用于‘无籽沙糖橘’花粉培养,一份用于‘沙糖橘’花粉培养。相应花粉撒播至培养基后于恒温培养箱中28 ℃培养10 h,以花粉管长度超过花粉粒直径作为萌发标准,在光学显微镜下观察并统计各处理的花粉萌发率,设置3次生物学重复,以未添加CrSPH蛋白组作为对照。
2 结果与分析
2.1 CrSPH基因克隆
以‘无籽沙糖橘’和‘沙糖橘’的花蕾 cDNA 为模板,利用引物CrSPH-F/R进行PCR扩增,均得到约400 bp特异条带(图1),经回收、克隆后测序表明‘无籽沙糖橘’和‘沙糖橘’CDS均为417 bp,编码138个氨基酸。两者间序列存在3个碱基差异,‘沙糖橘’第93位C突变为T、第190位G突变为A、第324位C突变G,从而导致‘沙糖橘’第64位缬氨酸(V)突变为异亮氨酸(I),108位天冬酰胺(N)突变为赖氨酸(K)(图2)。利用CrSPH-F/R引物对2个材料的基因组DNA进行扩增及测序,其序列长度也是417 bp,表明‘沙糖橘’和‘无籽沙糖橘’基因组DNA均不含内含子。利用ProtParam预测‘沙糖橘’CrSPH蛋白显示其分子量为16.05 kD,理论等电点为8.48,SignalP 6.0分析显示该蛋白不存在信号肽。

M. Marker; 1. Wuzishatangju; 2. Shatangju.

STJ. Shatangju; WZSTJ. Wuzishatangju.
2.2 不同部位花器官的基因表达分析
采用qPCR技术对花瓣、花丝、花药、柱头、花柱和子房等花器官部位的CrSPH基因表达量进行检测,结果如图3所示,CrSPH基因在‘沙糖橘’花器官不同部位的表达量差异较大,其中子房组织的表达量最高,且与其他部位表达量差异均达到显著水平;‘无籽沙糖橘’不同部位的表达量差异均未达到显著水平。在高表达的子房中,‘沙糖橘’表达量是‘无籽沙糖橘’的16倍,表明CrSPH基因在‘沙糖橘’中具有高度的组织表达特异性。

柱状图上不同小写字母表示花器官不同部位表达量差异显著(P<0.05)。
2.3 自交和异交授粉后不同时段的基因表达分析
利用qPCR检测了CrSPH基因在自交(‘无籽沙糖橘’ב无籽沙糖橘’)和异交(‘无籽沙糖橘’ב沙糖橘’)授粉后0,1,2,3,4,5,6,7 d的雌蕊组织表达量,相关结果见图4。由图4可知,异交授粉后第6,7天的表达量最高,与其他时段的差异均达到显著水平;异交授粉后第2~4天的表达量较高,与表达量较低的第1和第5天的差异达到显著水平,异交授粉第6天表达量是第1天表达量的12.4倍,表明CrSPH基因在异交授粉后不同时段的表达量差异显著。
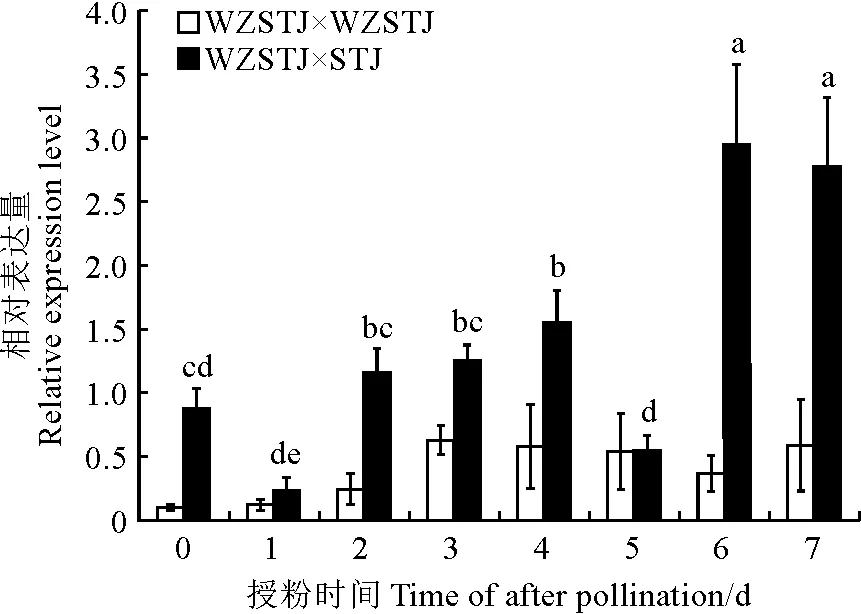
柱状图上不同小写字母表示授粉后不同时间的表达量差异显著(P<0.05)。
由图4可知,自交授粉后各时段的基因表达水平较低,其表达量呈现出先上升后下降的表达规律,以第3天的表达量最高,但不同时段的基因表达量差异未达到显著水平。
2.4 Southern 杂交分析
利用CrSPH基因内部不存在酶切位点的EcoR I和SacI酶以及存在1个酶切位点的ClaI酶对基因组DNA消化后,与DIG标记CrSPH基因的探针进行杂交实验,结果见图5。由图5可知,经EcoR I和SacI消化的DNA杂交出1条杂交带,而经ClaI酶消化的DNA出现2条杂交带,表明CrSPH基因在‘无籽沙糖橘’和‘沙糖橘’基因组中均以单拷贝形式存在。
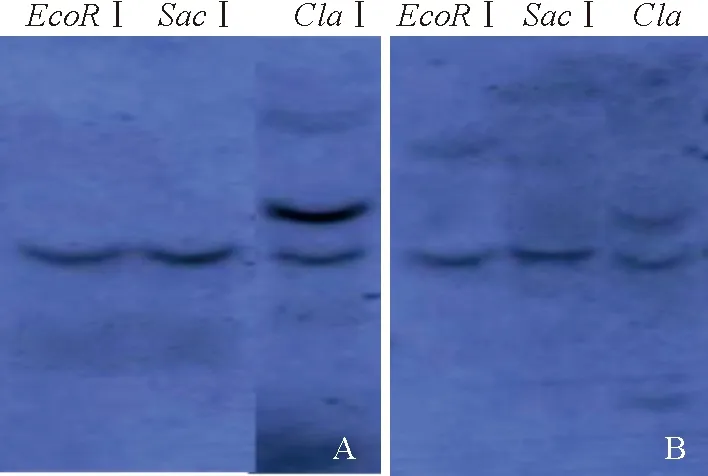
A.‘Wuzishatangju’, B. ‘Shatangju’.
2.5 CrSPH蛋白诱导表达及对离体花粉萌发影响
将线性化的pET32a表达载体与带有BamH I和SalI酶切位点的目的基因连接后,经转化、培养、测序鉴定后,成功构建含有CrSPH基因的原核表达载体pET32a-CrSPH。将该重组质粒转化大肠杆菌BL21(DE3),经IPTG诱导表达及SDS-PAGE电泳结果见图6。由图6可知,原核诱导表达的融合蛋白条带约37 kD,其中CK中的标签蛋白为21 kD,诱导获得的CrSPH蛋白约为16 kD,这与生物信息学预测的16.05 kD结果相吻合,表明该原核表达系统成功诱导表达出CrSPH蛋白。

M. Marker; CK. His tags protein; 1. ‘Wuzishatangju’;2. ‘Shtangju’.
将上述异源表达的CrSPH蛋白稀释成0.4,0.8,1.6,3.2,6.4 μg/μL浓度后添加到花粉培养基,观察其对花粉萌发率的影响,并以不添加外源CrSPH蛋白组为对照,结果见图7。由图7可知,‘无籽沙糖橘’CrSPH蛋白对2种花粉萌发率有一定影响,随着异源‘无籽沙糖橘’CrSPH蛋白浓度增加,‘无籽沙糖橘’花粉萌发率显著性降低;但‘沙糖橘’花粉在0.4,1.6 μg/μL的低浓度处理时萌发率较高,而在3.2,6.4 μg/μL的高浓度处理下萌发率反而下降。‘沙糖橘’CrSPH蛋白对‘无籽沙糖橘’和‘沙糖橘’的花粉萌发均无明显影响。

图柱上不同字母表示不同浓度处理下的差异显著性(P<0.05),*表示两材料间差异达到显著水平(P<0.05),**表示两材料间差异达到极显著水平(P<0.01)。
3 讨 论
多数果树属于配子体自交不亲和,其自交不亲和性是雌蕊关键因子S-RNase基因和花粉关键因子SLF/SFB基因相互作用结果[7,17,23-25]。Lai等[10]在研究金鱼草花粉自交不亲和反应中,将金鱼草花粉特异表达AhSLF-S2基因和雌蕊特异表达的AhS2-RNase基因转入矮牵牛(Petuniahybrida),结果分别在转基因矮牵牛的花粉和雌蕊中检测这两个基因的表达量,Qiao等[26]进一步通过花粉实验证明转了AhSLF-S2和AhS2-RNase基因的矮牵牛由SI转变成SC。然而蔷薇科花粉自交不亲和反应并非由于基因表达量差异导致,而是花粉S基因编码区发生了碱基的替换、缺失,导致S蛋白发生改变,从而导致由SI向SC转变[27]。Sassa等[4]发现SC突变体‘Osa-Nijisseiki’是由于SI日本梨‘Nijisseiki’的S4-RNase基因发生了突变,使得S4-RNase基因不能在花柱表达,导致SI向SC突变。Ushijima等[28]在梅(Prunusmume)中获得花粉突变造成的自交亲和单体型Sf,Sf基因结构中有6.8 kbp碱基插入,造成编码的F-box蛋白缺少了C-端序列,从而造成亲和突变。本研究中,CrSPH基因在‘沙糖橘’和‘无籽沙糖橘’CDS序列存3个碱基的差异(C→T,G→A,C→G),导致2个氨基酸残基发生改变(V→I,N→K)。在CrSPH基因在特异性表达的子房部位,‘沙糖橘’表达量是‘无籽沙糖橘’的16倍,推测‘无籽沙糖橘’的CrSPH基因突变和子房中低水平表达可能与其自交不亲和表型相关。
植物自交不亲和主要由1个单一的复等位基因S位点控制,该基因座上存在2个紧密连锁且分别在花柱和花粉中特异表达的基因,分别是雌蕊决定因子S-RNase和花粉决定因子SLF/SFB。柑橘属于S-RNase介导的GSI,主要通过抑制花粉管的生长实现自交不亲和性,因此S位点蛋白在花粉萌发和抑制花粉管生长中起着重要的作用[17]。Zhang等[18]提出泛素蛋白降解和液泡分拣途径假说,认为S-RNase通过细胞内吞作用从细胞外基质进入到花粉管后被运输到花粉的液泡中,在异花授粉的花粉管中,液泡膜未被破坏,S-RNase会被SCFCLF泛素化而降解,花粉管正常延伸;而自花授粉后,液泡膜由于某种机制会被破坏,S-RNase便释放到细胞质中,产生细胞毒素效应从而抑制自花花粉管的生长。SI除了雌蕊决定因子S-RNase和花粉决定因子SLF外,其他非S因子也参与自交不亲和反应。Beecher等[12]等发现通过转化单一的S-RNase基因不一定能够使自交亲和的烟草品种恢复拒斥自花花粉的能力,说明SI除了花粉和花柱S决定因子外,其他非关键因子也参与自交不亲和反应。Goldraij等[29]报道了HT-B作为非S因子可使不亲和花粉管中的液泡破裂,使得S-RNase酶从液泡中释放出来,从而抑制了花粉管生长。Thomas等[13]报道了PCD等非S因子不参与花粉管生长尖端的生长抑制,可能参与SI下游事件来保证花粉管生长抑制的不可逆性。Ren等[15]从无籽沙糖橘中克隆到1个CrSKP1-e基因,在花粉中特异表达,是SSK1基因的同源基因并参与SI反应。本研究中,CrSPH作为S位点蛋白的同源基因,CrSPH蛋白对花粉萌发表型效应也有显著影响,随着‘无籽沙糖橘’CrSPH蛋白浓度增加,‘无籽沙糖橘’花粉萌发率显著性降低;而‘沙糖橘’花粉在低浓度CrSPH蛋白处理时萌发率较高,在高浓度处理时萌发率下降,因此,笔者推测CrSPH蛋白作为非关键因子,可能通过参与花粉管液泡破裂或其他下游代谢过程参与柑橘SI反应过程,但具体如何作用,需要进一步遗传研究和功能验证。
