温州湾湿地互花米草地下芽功能性状及其影响因素
2023-09-19常宇欢王福京杨小兵
常宇欢 王福京 杨小兵
(1 温州大学生命与环境科学学院,浙江 温州 325035;2 温州大学三垟湿地生态环境研究院,浙江 温州 325035)
互花米草(Spartina alterniflora)因兼具有性和无性两种繁殖而拥有极强的繁殖和扩散能力(Tan et al.,2014),现已成为盐沼优势入侵植物。地下芽作为植物无性(克隆)繁殖的重要组成部分(Zhao et al.,2017),对于维持植物种群的动态稳定与更新具有重要作用。克隆繁殖是互花米草的主要繁殖方式(Zhao et al.,2017)。在营养充足、去叶干扰条件下,互花米草主要以克隆繁殖方式迅速扩展自己的分布区域(汤俊兵,2011)。环境因子的不同影响甚至决定着互花米草地下芽的形成和发展(Tomlinson et al.,2004),因此,掌握互花米草地下芽功能性状及其环境作用机制,对于控制其大范围入侵和科学管控具有重要的理论价值。
当前,国内外关于环境因子对地下芽功能性状的影响开展了大量研究(赵凌平等,2015)。在草地生态系统中,植物地下芽的数量和火烧频率、土壤水分及养分呈正相关(Benson et al.,2004;赵凌平等,2015)。在人为因素干扰下,适度放牧可显著提高高寒植物地下芽密度,但重度放牧会导致地下芽数量的减少(Evette et al.,2009)。然而,也有研究发现放牧强度对植物地下芽密度无显著影响(Qian et al.,2017)。通常情况下,植物地下芽密度会随着海拔和光照强度的增加而增大(Evette et al.,2009)。由此可见,地下芽功能性状与外界环境因子有着极其复杂的关系,但这方面研究主要集中于草地植物地下芽的形态性状,而关于湿地植物地下芽形态性状及化学计量的研究较为匮乏。
温州地处浙南沿海,湿地资源十分丰富,其中温州市湿地面积约占全省湿地面积的19.33%,滨海湿地约占全市湿地总面积的90%(马仁翻等,2014)。互花米草于20 世纪80 年代首次被引入浙江并广泛推广种植,之后迅速扩散,在温州地区分布面积已达4 000 余hm2(谭勇,2015),对沿海生物多样性和生态系统造成严重影响。鉴于目前温州湾湿地互花米草大规模入侵的现状和保护湿地的大背景下,开展互花米草芽功能性状的研究可对其科学管控提供理论依据,也可为滨海湿地保护提供新的研究思路。
1 材料与方法
1.1 研究区概况
温州湾湿地地处浙江省南部,由北向南分别是乐清湾、瓯江口、鳌江口3 处典型的亚热带滨海湿地(图1)。乐清湾西门岛(28°20′—28°22′N,121°11′—121°12′E)位于乐清北部,该地区多年平均气温为18.7℃,全年平均日照时数为1 687.4 h,无霜期约275 d,多年平均降水为2 043.1 mm。瓯江口(27°34′—27°35′N,120°34′—120°35′E)属于强潮河口,受海洋潮流影响较大,该地区多年平均气温为17.7℃,全年平均日照时数为1 638.2 h,无霜期约287 d,多年平均降水为2 271.7 mm。鳌江口(27°58′—27°59′N,120°50′—120°51′E)位于鳌江入海口,鳌江是浙江省独留入海的最小水系,也是全国三大涌潮江之一,该地区多年平均气温为17.9℃,全年平均日照时数为1 594.4 h,无霜期约248 d,多年平均降水为2 271.7 mm。
图1 研究区分布图Fig.1 Distribution map of studying area
1.2 研究方法
1.2.1 野外调查与样品采集2021 年12 月和2022年1 月,在互花米草地下芽的生长旺期,在3 处湿地各设置5 个样方(样方大小1 m×1 m,样方间隔100 m)。分别调查每个样方互花米草的群落密度。在每个样方,用剪刀齐地刈割25 cm×25 cm 面积内的地上植物并装袋(Hai et al.,2011),以测定互花米草地上生物量。滨海湿地植物地下芽主要集中在0~20 cm,用铁铲刈割地上植物后,挖出20 cm 的地下土块(地下植物及土壤),装袋带回室内以测定植物地下生物量。利用土壤环刀(直径37 mm,高度50 mm)采集0~20 cm 的土壤样品,装入密封袋带回室内以测定土壤的理化性质。
1.2.2 室内分析用清水冲洗地下芽样品(避免伤害根茎和芽之间的连接)。依据芽在母株上位置的不同,将地下芽分为水平生长的根茎顶芽、向上生长的根茎顶芽、根茎节芽和分蘖节芽4 种类型(张继涛,2009)。待样品晾干后,用10X 放大镜观察芽,统计芽的类型和数量,并计算地下芽的总密度(芽/m2)和4 种类型芽的分密度(芽/m2)(张继涛,2009)。使用游标卡尺测量互花米草地下芽的长度和直径,并计算芽长度与直径比等形态指标(张继涛,2009)。采用烘干称重法,于105℃杀青15 min 后,放入牛皮纸中以65℃烘干48 h 后用电子天平称重(精度为0.01 g),测定地下芽的生物量。利用H2SO4-H2O2消煮法,测定地下芽的全氮、全磷等化学计量指标。地下芽的全碳含量采用总有机碳分析仪(TOC-L-CPHSSM5000A)进行测定。参照《土壤农化分析》测定土壤含水率、pH值。采用ProCheck 手持多功能读数表(METER,USA)测定土壤电导率。利用H2SO4-H2O2消煮法测定土壤全氮、全磷含量。
1.2.3 数据分析采用SPSS 21.0 的单因素方差分析法(one-way ANOVA)对不同地区互花米草地下芽形态性状及生物量、化学计量及环境因子等指标进行差异显著性分析。通过冗余分析找出影响互花米草地下芽功能性状特征的关键环境因子和解释率。
2 结果与分析
2.1 不同地区互花米草种群密度和土壤理化特征
由表1 可知,3 个湿地土壤含水率、pH 值、土壤全氮和全磷无显著差异。乐清湾互花米草种群密度(101.82±20.41 株/m2)和鳌江口互花米草种群密度(89.86±6.88 株/m2)无显著差异,两者均显著高于瓯江口种群密度(59±9.69 芽/m2)。乐清湾互花米草地上生物量(227.33±38.53 g/m2)最高,显著高于鳌江口和瓯江口地上生物量(142.97±49.15 g/m2,142.07±22.39 g/m2),后 两者无显著差异。乐清湾土壤电导率(15.63±1.79 ms/m)最高,显著高于鳌江口和瓯江口土壤电导率(11.56±4.23 ms/m,10.25±1.47 ms/m),后两者无显著差异。乐清湾和鳌江口土壤碳氮比(1.83±0.08,1.75±0.14)无显著差异,两者均显著高于瓯江口土壤碳氮比(1.57±0.21)。
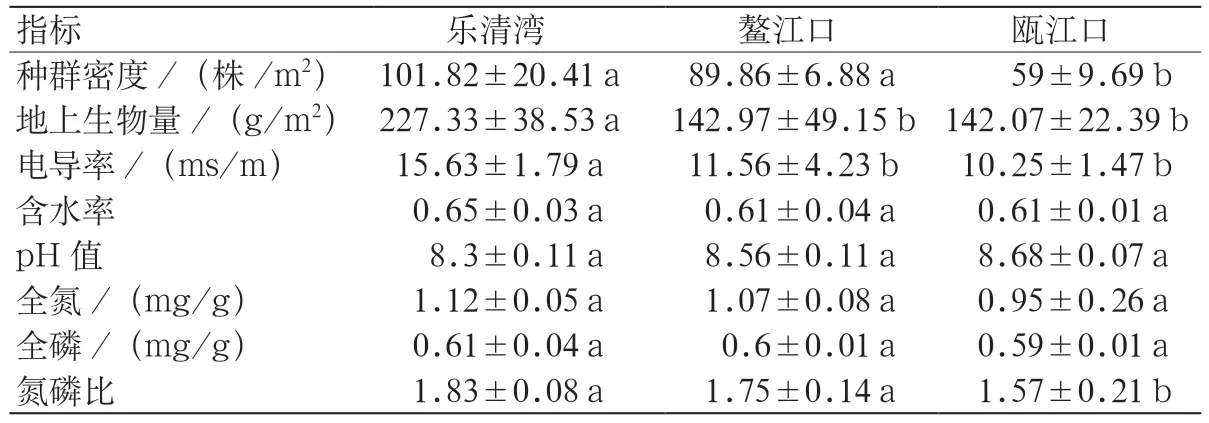
表1 不同地区互花米草种群密度、土壤理化特征比较Table1 Comparison of population density,soil physical and chemical characteristic of S.alterniflora in different regions
2.2 不同地区互花米草地下芽功能性状特征
由图2 可知,3 个地区互花米草分蘖节芽密度、芽总密度、芽生物量、芽长度和芽直径无显著差异。乐清湾和鳌江口水平生长根茎顶芽密度(16±7.84 芽/m2,12±5.72芽/m2)无显著差异,两者均显著高于瓯江口水平生长根茎顶芽密度(3.2±2.86 芽/m2)(图2a)。乐清湾和鳌江口向上生长根茎顶芽密度(12.8±3.46芽/m2,10.9±5.72 芽/m2)无显著差异,两者均显著高于瓯江口向上生长根茎顶芽密度(5.8±5.72 芽/m2)(图2b)。乐清湾根茎节芽密度为16±8.09芽/m2,显著高于鳌江口和瓯江口根茎节芽密度(9.6±5.72 芽/m2,6.4±2.86 芽/m2),后 两者无显著差异(图2c)。乐清湾芽长度与直径比(45.57±7.62)显著高于鳌江口和瓯江口芽长度与直径比(31.4±5.23,27.08±7.37),后两者无显著差异(图2i)。
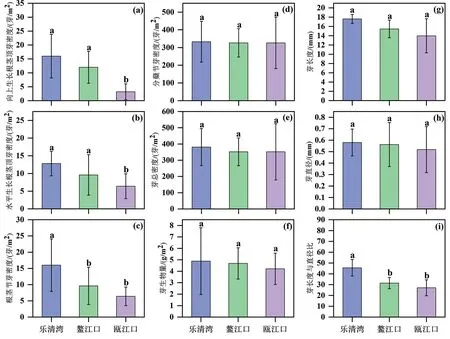
图2 不同地区互花米草地下芽形态性状及生物量比较Fig.2 Comparsion of belowground bud morphological traits and biomass of S.alterniflora in different regions
由表2 可知,3 个地区互花米草地下芽全碳、全磷、碳氮比、碳磷比无显著差异。乐清湾和鳌江口芽全氮(21.68±0.57 mg/g,21.22±2.28 mg/g)无显著差异,两者均显著高于瓯江口芽全氮(18.85 ±0.04 mg/g)。瓯江口芽碳氮比(23.32±0.04)显著高于鳌江口和乐清湾碳氮比(21.04±2.16,20.05 ± 0.25),后两者无显著差异。乐清湾和鳌江口芽氮磷比(7.71±0.63,7.39±0.49)无显著差异,两者均显著高于瓯江口芽氮磷比(6.19± 0.19)。
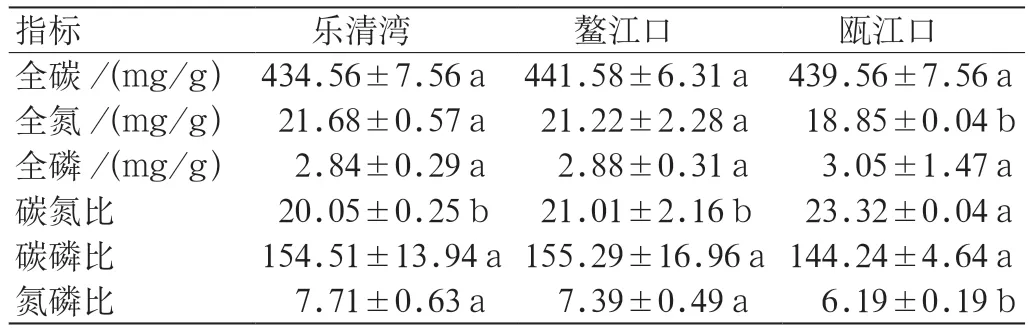
表2 不同地区互花米草地下芽化学计量学比较Table 2 Comparsion of belowground bud chemometrics of S.alterniflora in different regions
2.3 互花米草地下芽功能性状的影响因素
冗余分析表明:土壤含水率对互花米草地下芽形态性状及生物量的解释度最高,其解释度为17.8%(P=0.022),土壤全磷含量对地下芽形态性状及生物量的解释也达到显著水平,解释度为14.7%(P=0.03),其他因子对地下芽形态性状及生物量的解释未达到显著水平(图3a)。在影响地下芽化学计量的因子中,土壤含水率是最重要的变量,解释度为27.4%(P=0.04),种群密度对地下芽化学计量的解释也达到显著水平,其解释度为21.7%(P=0.08),其他因子对地下芽化学计量的解释未达到显著水平(图3b)。
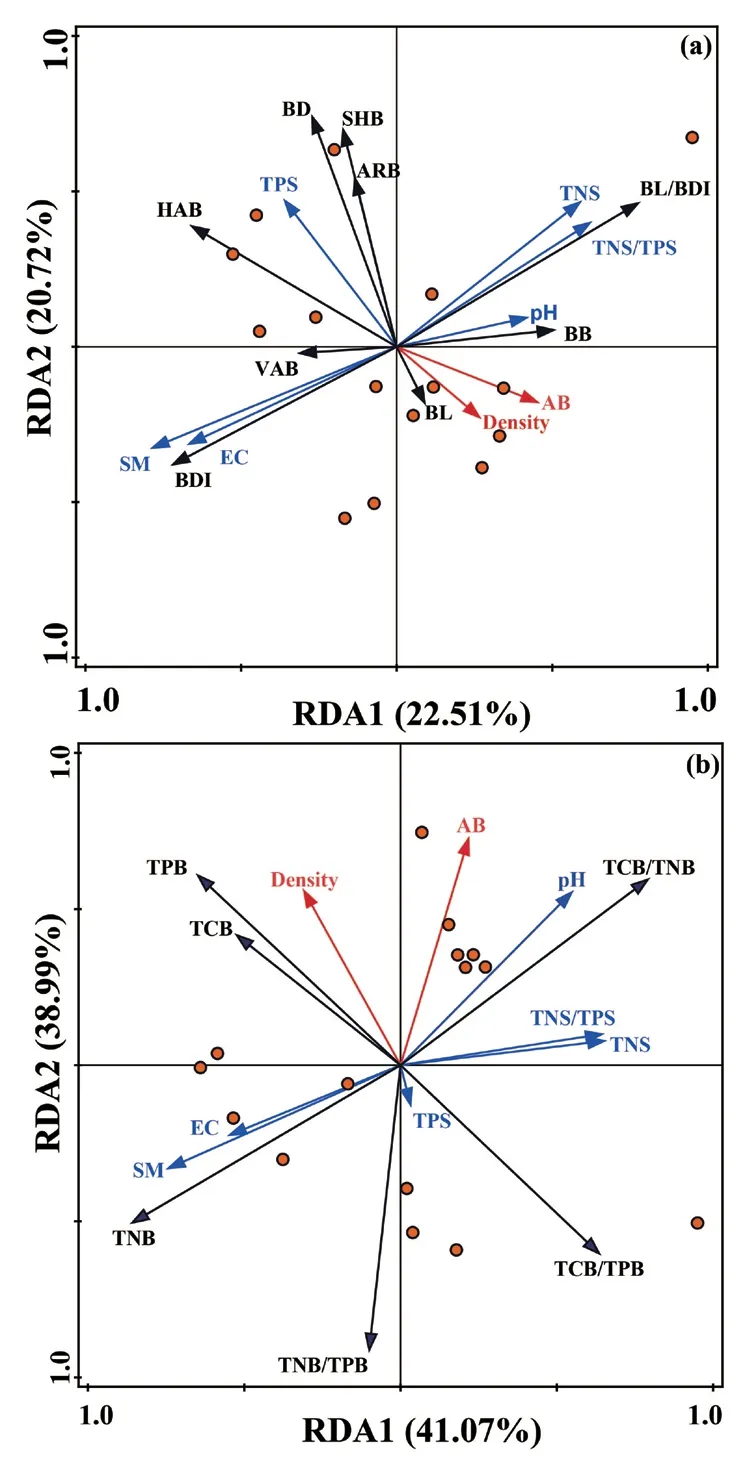
图3 温州湾湿地互花米草地下芽功能性状与环境因子的冗余分析 Fig.3 RDA of belowground bud functional traits of S.alterniflora and environmental factors in Wenzhou Bay Wetland
3 讨论
3.1 不同地区间互花米草地下芽功能性状的差异
温州湾湿地不同地区互花米草地下芽的形态性状及生物量存在一定的差异性(图2),这可能与其所处的土壤含水率和土壤养分差异有关。在本研究中,乐清湾、鳌江口、瓯江口的土壤含水率无显著差异,但乐清湾湿地的土壤含水率略高于鳌江口和瓯江口湿地。乐清湾湿地土壤含水率高,促进了该地区地下芽的生长和萌发,导致该地区拥有最高的地下芽密度、地下芽长度、地下芽生物量。这与叶学华等(2013)对两种根茎型克隆植物地下芽形态特征与水分相互关系的研究结果相一致。相比之下,瓯江口湿地的土壤含水率低,不利于地下芽的生长与发育,因此,瓯江口湿地的地下芽形态性状各指标及生物量相对较低。此外,在湿润的环境中,较高土壤含水率能够促进地上植物的光合作用,产生的有机质会进入到土壤中转变为根茎芽,利于根茎芽的产生和发育(Ding et al.,2019)。相反,在较为干旱缺水的环境中,多数植物会选择产生较多的分蘖节芽来维持地上植被的生长和更新(张继涛,2009)。张继涛等(2015)的研究也证实,在水分匮乏的土壤条件下,分蘖节芽的数量增加,因此,土壤水分的差异也是导致瓯江口湿地分蘖节芽密度较高的原因之一。不同地区地下芽形态性状及生物量的差异也可能与土壤养分的差异有关。植物地下芽的形态性状及生物量与土壤养分密切相关,适宜的土壤养分能够促进地下芽萌发、生长和发育(焦德志等,2015)。土壤中氮、磷元素等是地下芽生长过程中的必须营养元素(Tomlinson et al.,2004)。本研究中乐清湾湿地的土壤氮、磷含量高,可以有效地促进植物细胞生理生化过程,促进植物地下芽的生长和发育(Dalgleish et al.,2009)。相反,较低的土壤氮、磷含量不利于植物地下芽的生长,严重抑制植物的生长和发育,因此,鳌江口湿地和瓯江口湿地的地下芽密度、生物量、长度等形态性状普遍偏低。这与焦德志等(2015)对芦苇地下芽动态与土壤养分相互关系的研究结果相一致。
温州湾湿地不同地区互花米草地下芽全氮、碳氮比及氮磷比存在一定的显著差异性(表2),这可能与其所处的土壤含水率和养分差异有关。乐清湾湿地的土壤含水率和养分较高,湿润且肥沃的环境提高了土壤资源的可利用性,有利于土壤矿化作用、凋落物分解和根系呼吸作用等生理生化过程的进行(李新乐等,2020),提高了植物根系对土壤氮素的吸收能力,导致了植物体地下芽养分的积累。不同地区的地下芽全碳、全磷、芽碳氮比、芽氮磷比则无显著差异,这可能与互花米草对不同环境的响应差异有关。
3.2 影响互花米草地下芽功能性状的环境因子
冗余分析表明,土壤含水率、土壤全磷含量、种群密度是影响互花米草地下芽功能性状的主要环境因子(图3),其中,土壤含水率和地下芽直径、地下芽全氮、向上生长和水平生长的根茎顶芽密度呈正相关。适宜的土壤含水率促进植物对土壤养分的利用率,导致植物体内养分含量的增加,促进了地下芽的生长与繁殖(Klimesová et al.,2007)。但土壤含水率和地下芽碳氮比呈负相关,体现了滨海湿地植物互花米草地下芽化学计量对外界环境因子相应的差异性(周翰文,2016)。土壤全磷和地下芽总密度、地下芽分密度均呈正相关,充足的土壤磷含量利于地下芽的生命活动,导致地下芽生物量和根茎顶芽密度的增加(焦德志等,2015)。此外,种群密度和地下芽长度、地下芽生物量、地下芽长度与直径比、地下芽全碳、地下芽全氮、地下芽全磷呈正相关,这与张海艳等(2014)对羊草植物群落的研究结果不一致。这可能由于本研究在冬季,低温抑制了群落地上凋落物的分解,导致养分在土壤表层聚集(丁文广等,2010),因此,地下芽通过增加对土壤养分的吸收而维持基本的生命活动,以此来维持互花米草种群的扩散与更新。在影响互花米草地下芽功能性状的主要因子中,土壤含水率是影响互花米草形态性状及生物量、化学计量指标最重要的解释变量,这和王娜等(2021)对若尔盖湿地植物地下芽多样性特征的研究结果相一致。作为影响温州湾湿地互花米草地下芽功能性状的最关键环境因子,维护和恢复湿地水环境将成为保护湿地植物多样性和管控互花米草扩散的关键。
4 结论
1)温州湾湿地不同地区互花米草地下芽功能性状存在一定的差异性。其中,乐清湾湿地地下芽形态性状、生物量、全碳及全氮等指标最高,而鳌江口和瓯江口湿地地下芽功能性状相应指标较低。
2)相关性与冗余分析表明,不同地区互花米草地下芽功能性状与其土壤含水率和养分的差异性有关,土壤含水率是调控互花米草地下芽功能性状的最关键环境因子。
3)控制互花米草快速入侵的关键在于对湿地水文环境的控制与管理,维护和恢复湿地水环境是实现滨海湿地植物多样性和互花米草科学管控的有效途径。
