不同生长年限固阳黄芪根际土壤微生物多样性及差异
2023-09-19刘鸽赵博刘扬李俊达孙旭红付树志杨可青郝佳瑞刘惠荣
刘鸽,赵博,刘扬,李俊达,孙旭红,付树志,杨可青,郝佳瑞,刘惠荣
(内蒙古农业大学 生命科学学院,内蒙古呼和浩特 010018)
黄芪(Astragalus membranaceus)属豆科多年生草本植物,是我国大宗常用滋补类中药材,具有固表补气、托毒生肌、利水消肿、降血糖及抗病毒等功效[1]。现代药理学研究及临床应用表明,黄芪中的活性成分可起到抗肿瘤、抗衰老、多靶点免疫调节等作用,已被广泛用于相关疾病治疗。
根际是土壤-根系-微生物三者紧密结合且相互影响的场所,不同植物都有其特定的根际微生物组群。相关研究表明,根际微生物群落中的不同微生物能够与植物进行互作,在促进植物生长、抑制土传病害、平衡土壤养分及提升养分利用率等方面具有重要意义[2]。目前科研工作人员对黄芪的研究侧重于黄芪的活性成分,仅有少量学者对黄芪根际微生物群落做了相关研究。此外,前人未曾对内蒙古固阳县不同生长年限黄芪根际土壤微生物群落结构进行研究。固阳县位于内蒙古中西部,典型的半干旱大陆性气候有利于黄芪有效成分的积累。调查发现,内蒙古黄芪生态种植主要集中在固阳县及其周边,且“固阳黄芪”已被国家核准注册为中国地理标志商标。目前,由于黄芪市场需求增加、多年生黄芪种植成本高及易感病,当地黄芪种植年限缩短至2 年。因此本试验采用高通量测序技术,对内蒙古固阳县不同种植年限黄芪的根际微生物进行多样性分析,以期为黄芪种植及开发黄芪功能微生物提供参考依据。
1 材料与方法
1.1 试剂与仪器
PowerSoil DNA Kit 试剂盒,美国QIAGEN 公司;LSC-373G 型4 ℃冰箱,浙江星星冷链股份有限公司;DW-HL398S 型-80 ℃低温冷冻箱,中科美菱低温科技股份有限公司;SX-500 高温高压灭菌锅,日本TOMY 公司;SW-CJ-1FD 型超净无菌工作台,苏州净化公司;ABI GeneAmp®9700 型PCR 仪,美国ABI 公司;DYY-6C 电泳仪,北京六一生物科技有限公司。
1.2 采样区概况及样品采集
供试土样采于2021 年7 月,来源于固阳县二年生、三年生黄芪根际。在每个采样点随机布设3 个1.5 m×1.5 m 样方,每样方随机选取5 株植株。采用抖根法收集土样,用毛刷将紧贴附于根系的土壤装入无菌自封袋,编号。两组黄芪根际土壤样品分别命名为Ⅱ-TYB(二年生黄芪根际土壤,取样地海拔1 550 m,位于北纬41°06′59″、东经110°18′54″)、Ⅲ-TYA(三年生黄芪根际土壤,取样地海拔1 540 m,位于北纬41°06′56″、东经110°18′25″)。将样品迅速放入4 ℃冰盒保存,及时带回实验室,将采集的土样分别放入4 ℃与-80 ℃冰箱保存。
1.3 土壤微生物基因组DNA 的提取、PCR 扩增及测序
利用PowerSoil DNA Kit 试剂盒提取根际土壤微生物总DNA,使用NanoDrop 2000 检测DNA 浓度和纯度。用引物338 F(5'-ACTCCTACGGGAGGCAGCAG-3')和806 R(5'-GGACTACHVGGGTWTCTAAT-3') 扩增细菌16S rDNA 序列的V3-V4 区,用引物ITS1F(5'-CTTGGTCATTTAGAGGAAGTAA-3')和引物ITSIF2R(5'-GCTGCGTTCTTCATCGATGC-3')扩增真菌ITS1区。PCR 反应体系:10×Buffer 2 μL,2.5 mmol/L dNTPs 2 μL,rTaq 聚合酶0.2 μL,两条引物(5 μmol/L)各0.8 μL,DNA 模 版10 ng,BSA 0.2 μL,ddH2O 补 足至20 μL。反应条件为95 ℃预变性3 min,95 ℃变性30 s、55 ℃退火30 s、72 ℃延伸45 s(细菌27 个循环、真菌35 个循环),72 ℃延伸10 min,4 ℃保存。用2%琼脂糖凝胶电泳检测PCR 产物的片段及长度,并对合格产物进行纯化、定量,对纯化后的PCR 产物建库,质检合格后使用Illumina Miseq PE300 平台进行测序。
1.4 数据分析与统计
对所得原始测序序列进行质控拼接,根据97%相似度对序列进行OTU 聚类并剔除嵌合体。利用RDP classifier 对每条序列进行物种分类注释,比对Silva 16S rRNA 数据库(Release 138)及Unite 真菌数据库(Release 8.0),设置置信度阈值为70%。并统计每个样本在不同物种分类水平下的群落组成。基于测序的数据,采用mothur 软件计算Alpha 多样性、Chao、Shannon指数等,采用Wilxocon 秩和检验进行Alpha 多样性的组间差异分析;采用Wilxocon 秩和检验在属水平进行组间差异显著性检验,确定丰度显著差异的微生物。
2 结果与分析
2.1 不同年份黄芪根际土壤微生物测序数据统计
本次测序共得到细菌有效序列290 249 条,平均长度约为415 bp,测序质量值Q30 为93.38%~93.40%,Q20 为97.77%~97.81%;真菌有效序列321 885 条,平均长度约为235 bp,测序质量值Q30 为99.00%~99.43%,Q20 为99.50%~99.72%。细菌、真菌根际样本经过最小样本序列数标准均一化处理后,分别得到185 538、259 554 条有效序列,在97%的序列相似度水平上,获得细菌、真菌OTU 数分别为3 875 个、1 082 个。
2.2 不同年份黄芪根际土壤微生物Alpha 多样性指数分析
细菌多样性指数中,Ⅱ-TYB 的Chao、Shannon、Simpson 指数分别为2 883.2、6.186 8、0.008 3,Ⅲ-TYA的Chao、Shannon、Simpson 指数分别为2 996.5、6.191 0、0.008 9,Ⅲ-TYA 的3 个指数均高于Ⅱ-TYB,说明细菌群落丰度随着生长年限的增加而增加,但不同年份黄芪根际细菌多样性指数无显著差异。
真菌多样性指数中,不同年份黄芪根际真菌Chao指数存在显著差异,随着时间推移,Chao 指数升高,真菌群落多样性增加。总体来说,随着黄芪种植时间的增加,根际微生物丰富度升高。不同年份黄芪根际土壤细菌和真菌覆盖度均在0.97 以上,说明本研究中测序量和测序深度合理。
2.3 不同生长年限黄芪根际土壤微生物群落组成
2.3.1 细菌群落组成分析
从门水平上看,2 组样品优势门类主要是放线菌门(Actinobacteriota)、变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、酸杆菌门(Acidobacteriota),合计占比在80%以上,其中放线菌门与变形菌门所占比例最高,如图1(a)所示。随着黄芪年限的增加,土壤中不同细菌门类的相对丰度发生变化,放线菌门、变形菌门占比相对增加,绿弯菌门与酸杆菌门占比相对降低。属水平上,主要优势菌属为节杆菌属(Arthrobacter)、芽生球菌属(Blastococcus)、norank_f__JG30-KF-CM45、微枝形杆菌属(Microvirga)与类诺卡氏属(Nocardioides),如图1(b)所示。各属在不同生长年限中丰度不同,其中节杆菌属与芽生球菌属在Ⅲ-TYA 样品中相对丰度最高,分别占5.58%、5.39%,在Ⅱ-TYB 中分别占比5.37%、3.93%。对比可知,随着黄芪年限增长,节杆菌属、芽生球菌属与微枝形杆菌属在黄芪根际土壤中比例增加,可能是这3个属菌株与黄芪植株形成了相对长久稳定的关系。

图1 黄芪根际细菌在门属水平上的相对丰度
2.3.2 真菌群落组成分析
从门水平上看,根际土壤真菌优势门类有子囊菌门(Ascomycota)、被孢霉门(Mortierellomycota)、担子菌门(Basidiomycota),如图2(a)所示。子囊菌门在二年生与三年生黄芪根际土壤真菌中占比分别为82.87%、92.73%,被孢霉门占比分别为11.18%、1.67%,担子菌门占比分别为3.97%、4.41%。对比可知,子囊菌门与担子菌门丰度呈逐年递增趋势,被孢霉门丰度变化则相反。属水平上,Ⅲ-TYA 中相对丰度较大的物种分别为刺孢属(Phaeomycocentrospora)、新刺毛茎点霉属(Neosetophoma)、链格孢属(Alternaria)、赤霉属(Gibberella),相对丰度依次为24.93%、9.93%、9.08%、7.89%,Ⅱ-TYB 中排名较前的为赤霉属(Gibberella)、刺孢属(Phaeomycocentrospora),被孢霉属(Mortierella)、毛壳霉属(Chaetomium),相对丰度依次为18.14%、11.67%、11.05%、6.81%,如图2(b)所示。由此可知,两组样品真菌丰度存在较大差异,随着黄芪的生长,土壤根际真菌的群落结构发生一定改变。
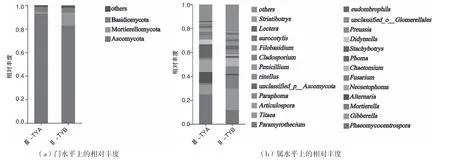
图2 黄芪根际真菌在门属水平上的相对丰度
2.3.3 组间差异显著性检验
对不同年份黄芪根际土壤样本中细菌和真菌在属水平进行组间差异分析(置信区间为95%,P<0.05)。如图3 所示,就细菌而言,未命名属(norank_f__norank_o__211ds20)仅存在于三年生黄芪根际土壤样本,存在显著性差异;二年生黄芪根际土壤中异短链球孢菌属(Allocatelliglobosispora)的丰度显著高于三年生。真菌方面,三年生黄芪根际土壤组中,Udeniomyces丰度显著高于二年生同属的丰度,为三年生黄芪根际土壤独有,但多孢囊霉属(Diversispora)的丰度则相反。
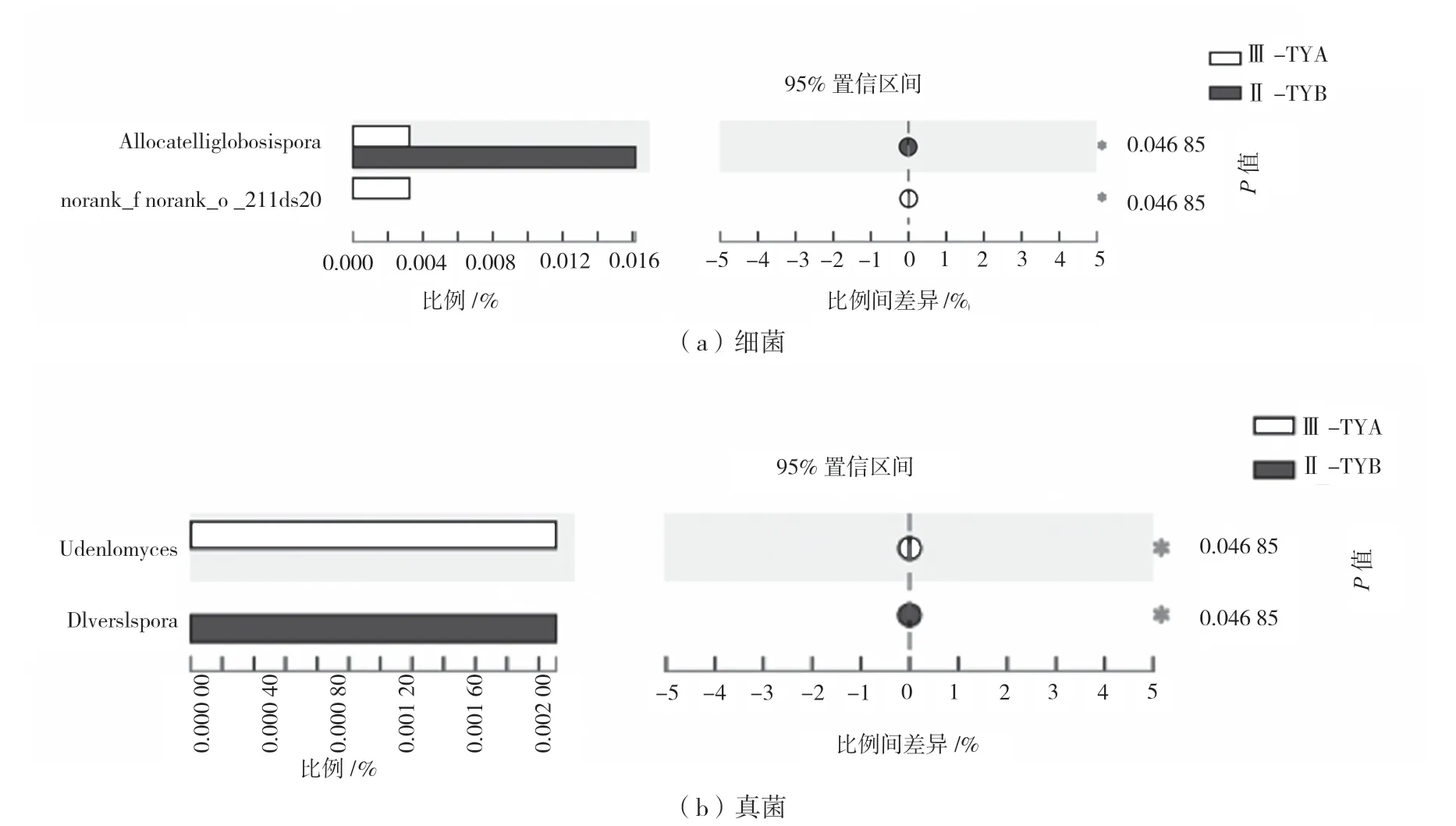
图3 黄芪根际微生物组间差异(属水平)
3 讨论
土壤微生物是陆地生态系统中植物生产力的关键驱动因素,研究黄芪根际土壤微生物群落结构和丰度变化,对黄芪的种植具有重要意义。本文首次研究了内蒙古包头市固阳县不同生长年限黄芪根际土壤微生物多样性的变化规律,结果表明,随着黄芪生长年份的增加,黄芪根际细菌与真菌多样性均呈增加趋势,这与康宝玲等[1]研究结果相似。本研究发现,不同年限黄芪根际土壤细菌优势门类主要为放线菌门、变形菌门。真菌优势门最丰富的是子囊菌门,其次为被孢霉门、担子菌门。牟建平等[3]在研究甘肃的6 个黄芪种植区根际土中也发现了同样的规律,由此可见不同产地的黄芪根际土壤微生物优势菌门组成具有共性。LI 等[4]在研究陕西省5 个不同种植年限的黄芪根际土中发现,除变形菌门外,拟杆菌门也为根际主要优势细菌门,且拟杆菌门丰度在1 ~4 年生黄芪根际土中表现出增加趋势。本研究中放线菌门、变形菌门的占比随着黄芪生长年限增加均发生小幅度增加。以往研究表明,放线菌门可参与有机质的分解,在植物生长、土壤代谢中发挥重要作用,变形菌门为药用植物根际微生物主要类群,且有助于提升药用植物质量和产量,可见此类菌门根际微生物显著富集可能有利于该地区黄芪的生长及有效成分积累。本研究发现子囊菌门、被孢霉门、担子菌门是黄芪根际土壤中丰度最大的真菌类群,子囊菌门与担子菌门丰度在土壤中呈逐年递增趋势,被孢霉门丰度变化则相反。LI 等[4]的研究也发现子囊菌门在1 ~4 年生的黄芪根际土中呈现出逐年增加趋势,不同的是,本试验还发现担子菌门的丰度也会随年份增加,被孢霉门丰度随年份减少。众多研究显示[5],子囊菌门与担子菌门菌株为关键的真菌分解者,二者比例可影响土壤抗性,抑制枯萎病。可见这些微生物随年份发生波动变化,对本地区黄芪的生长发育可能具有较大影响。
本研究中,两样品根际土壤细菌的主要优势细菌属均为节杆菌属(Arthrobacter)、芽生球菌属(Blastococcus)、未命名属(norank_f__JG30-KF-CM45)、微枝形杆菌属(Microvirga)与类诺卡氏属(Nocardioides),随着黄芪种植年限增加,节杆菌属、芽生球菌属与微枝形杆菌属在黄芪根际土壤中比例增加。前人大量研究已表明[6-7],节杆菌属、类诺卡氏属与微枝形杆菌属的菌株均为有益菌。节杆菌属具有较强的环境适应性,在环境污染物的分解代谢中扮演重要角色,具有固氮促生的能力;微枝形杆菌属为根瘤菌,与植物共生可结瘤,具有显著的固氮活性[6];类诺卡氏属可用于降解原油、污水治理、生物防护。此外,本研究中,不同样品存在显著差异的还有多孢囊霉属,其在二年生黄芪根际土壤中丰度显著高于三年生。已有研究发现[8],多孢囊霉属是一类丛枝菌根真菌,菌根真菌可与植物形成菌根互惠共生体,诱导激发植物体内的防御机制,减轻病害对植物造成的伤害。肖龙敏等[9]研究发现,该菌属还存在于种植10 年及24 年的枸杞根际土壤中,由此可见此类微生物与植物生长发育过程有密切联系,但对其是否可与固阳黄芪建立共生体系及对黄芪生长和药用成分的积累有无促进作用仍需要深入研究。
综上可以看出,固阳黄芪根际微生物中,一些微生物与黄芪的生长发育可能密切相关,这些微生物群组具有潜在研究价值,未来可深入研究此类微生物的功能及其影响黄芪根内有效成分积累的作用机制。
4 结论
本文对内蒙古包头市固阳县不同生长年限黄芪根际土壤微生物多样性进行研究,发现随着黄芪生长年份推移,黄芪根际细菌与真菌多样性均呈增加趋势,且两组样品真菌丰度存在较大差异。不同生长年限黄芪根际土壤中主要优势细菌均为放线菌门、变形菌门;优势真菌为子囊菌门、被孢霉门、担子菌门。放线菌门、变形菌门、子囊菌门与担子菌门丰度随着黄芪生长年限增加均发生增大。不同生长年限黄芪根际土壤的主要优势细菌属为节杆菌属、芽生球菌属、未命名属、微枝形杆菌属与类诺卡氏属。二年生黄芪根际土壤优势真菌属为赤霉属、刺孢属、被孢霉属、毛壳霉属,三年生优势真菌属包括刺孢属、新刺毛茎点霉属、链格孢属、赤霉属。刺孢属丰度随黄芪生长年份的增加而增加,赤霉属变化却相反。
