基于ISSR分子标记的中华鳖种群遗传多样性分析
2023-09-19丁淑荃吴梦晓崔鑫炎谢俊杰
丁淑荃,吴梦晓,尤 昆,袁 娟,崔鑫炎,黄 梅,谢俊杰,米 笛
(1.安徽农业大学动物科技学院,安徽合肥 230000;2.肥东县水产技术推广站,安徽合肥 231600 ;3.定远县绿色亲情种养殖专业合作社,安徽滁州 233200)
中华鳖(Pelodiscussinensis)因其蛋白质含量丰富,药用价值高,成为我国经济价值最高,养殖规模最大的龟鳖类,2021年鳖类的养殖总产量可达364 878 t[1]。我国中华鳖种质资源丰富,但缺乏有效的亚种分化,常把不同地理种群的名作为中华鳖的同物异名;尽管在分类学中它们同属于中华鳖,但不同水系和地理居群的品系之间存在巨大的生理和性状差异。具有良好经济性状的各品系鳖可以为种质研究和良种选育提供丰富的选择性培养材料。但近年来外来鳖种的引进以及持续近交繁殖,导致中华鳖品种纯度下降与质量下滑,降低了种质间遗传多样性[2]。目前,种质资源的严重退化已成为我国中华鳖产业可持续发展的重大障碍,因此现有中华鳖种质资源的遗传多样性的统计分析工作迫在眉睫。
此前,中华鳖常基于形态学进行分类,由于性状受发育阶段和环境因素的影响,仅显示有限的变异,因此难以固定具体性状并得到稳定的良种资源[3]。近年来,分子标记技术作为种群遗传和系统发育研究中的有效手段,已成为遗传多样性评估中的重要方法,不仅有助于研究种群结构、分类问题,还能帮助种质鉴定,确定种质保护的优先级[4]。分子标记技术以聚合酶链式反应(PCR)为基础,主要包括简单重复序列间扩增(ISSR)、随机扩增多态DNA(RAPD)、简单重复序列(SSR)和扩增片段长度多态性(AFLP)[5]。ISSR作为一种基于SSR基础的分子标记技术,已被广泛应用于品种鉴定、基因定位、遗传多样性、进化及分子生态学研究[6]。谭舜等[7]建立了红尾仙女虾ISSR的反应体系。ISSR分析表明,崇明白山羊的公羊具备更高的遗传水平[8];丹东和秦皇岛松江鲈鱼群体分化程度较低[9];分布于广东从化的平胸龟在物种水平和居群水平的遗传多样性水平均要高于福建寿宁地区[10]。与SSR标记相比,ISSR不需要基因组序列,更加省时且经济。与RAPD和RFLP相比,ISSR除了可评估密切相关的个体之间的遗传多样性之外,还具备可靠性、多态性高、成本低及高效等特性[11-12]。
遗传多样性和变异性是生物对环境变化作出适应性反应的基础,拥有足够丰富的遗传多样性的物种在应对不断变化的环境和竞争者时具有更大的优势[13]。因此,对中华鳖种群遗传多样性以及种内、种间、种群结构的研究,为中华鳖种质资源保护提供一定的研究基础,并为新品种选育改良提供数据参考。分子标记技术以其先进的计算方法从基因层面为分析遗传多样性和了解种群系统发育关系提供了新的切入点。笔者利用ISSR技术对4个地方品系以及1个杂交品系的中华鳖进行分子标记,评估了不同品系中华鳖遗传多样性、各种群间亲缘关系和遗传结构,为辅助现有中华鳖品种选育、品种分类以及品种表征鉴定等提供基础资料,同时为中华鳖遗传资源的有效评价和利用提供重要信息。
1 材料与方法
1.1 试验材料试验中华鳖来自5个不同居群,均由安徽省滁州市定远县绿色亲情种养殖专业合作社提供,具体特征和信息见表1。活体中华鳖运至实验室暂养3 d后取样。纱布擦拭体表,并用乙醇体表消毒后取腿部肌肉,液氮冷冻后置于-80 ℃保存备用。
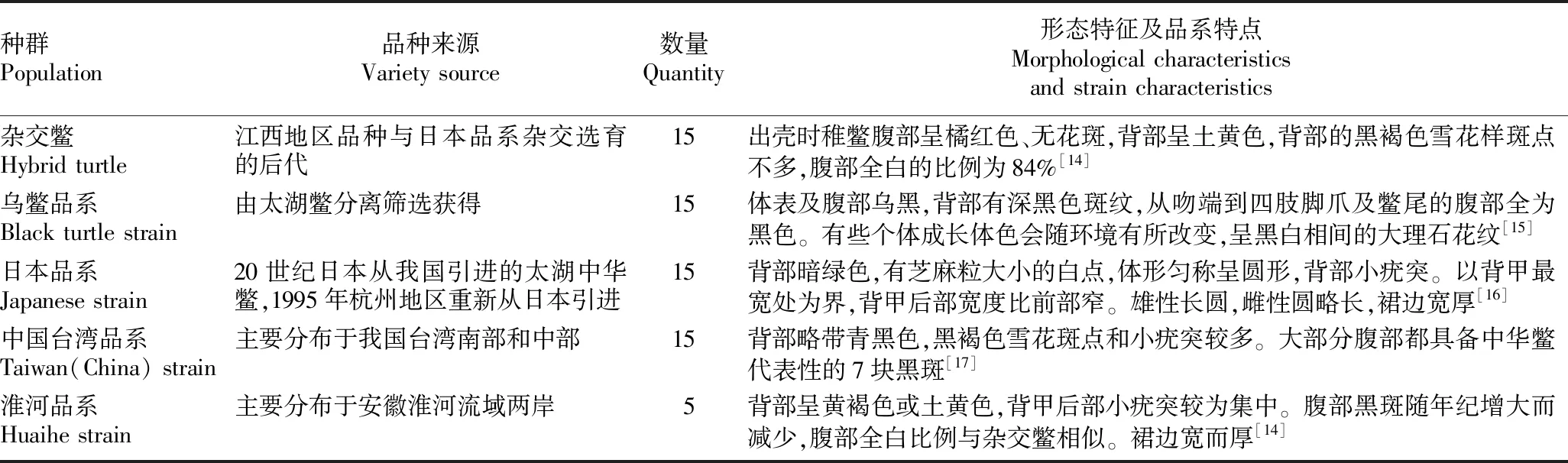
表1 中华鳖5个供试居群的基础信息
1.2 DNA的提取及检测采用Ezup柱式动物基因组DNA抽提试剂盒(生工生物工程(上海)股份有限公司)提取肌肉组织基因组DNA。1%琼脂糖凝胶电泳检测DNA纯度,超微量分光光度计检测核酸浓度,所提取DNA的OD260/OD280为1.70~1.94,完整度良好。
1.3 PCR扩增与电泳检测所用引物序列来源于加拿大哥伦比亚大学公布的第九套ISSR引物序列,由生工生物工程(上海)股份有限公司合成。筛选出10对多态性高、扩增条带清晰的ISSR引物用于扩增(表2)。反应体系为20 μL:cDNA 2.0 μL,2×RapidTaqMaster Mix 10.0 μL,引物(10 μmol/L)0.8 μL,ddH2O 7.2 μL,扩增反应程序为:95 ℃预变性5 min;95 ℃变性15 s,52 ℃退火30 s,72 ℃延伸30 s,进行35个循环;72 ℃彻底延伸5 min,4 ℃保存。扩增后的产物通过1.5%琼脂糖凝胶150 V电泳1 h,置于紫外凝胶成像仪中拍照保存后进行条带统计与分析。
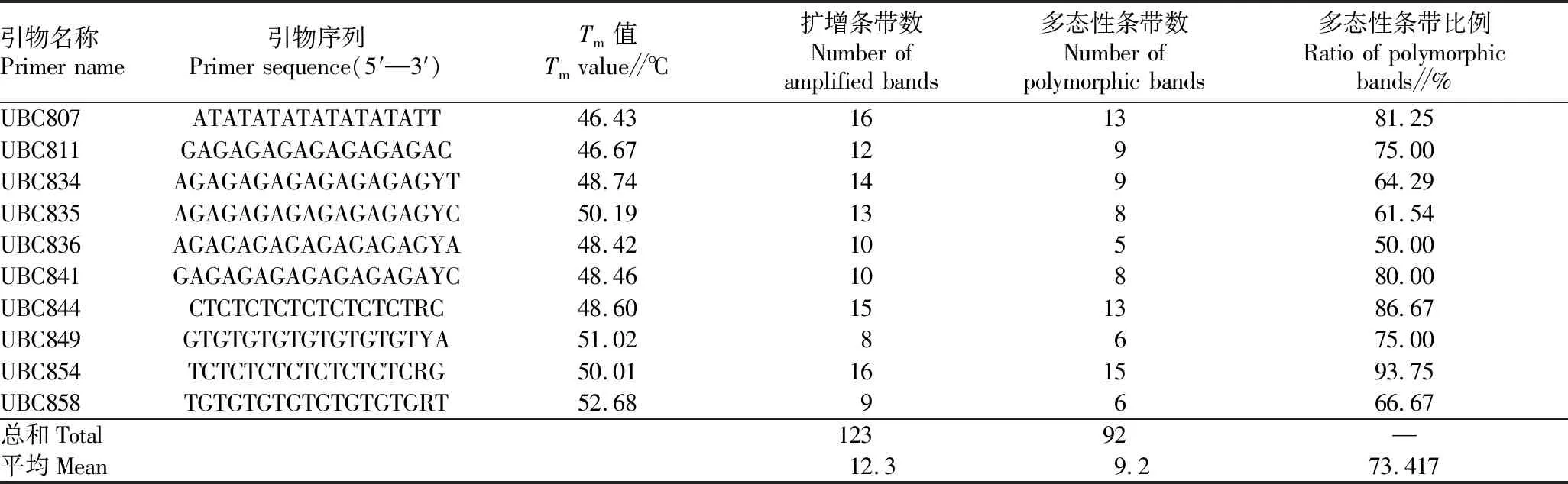
表2 ISSR引物序列及其扩增产物的多态性
1.4 数据统计与分析由于ISSR为显性标记,同一引物下PCR产物电泳迁移率一致的条带代表1个等位基因位点。根据同一位点条带的有无记录电泳结果,有带的记为1,无带的记为0,将所得数据输入Excel表格构建0/1矩阵。统计总条带数、多态性位点数、多态性条带数,计算多态性条带(PPB)比例。用Popgen 32软件计算等位基因数(Na)、有效等位基因数(Ne)、多态性位点(PPL)比例、Nei’s基因多样性指数(H)、居群总遗传多样性指数(Ht)、Shannon多样性指数(I)、居群总遗传多样性指数(Ht)、居群内遗传多样性指数(Hs)、基因分化系数(Gst)和基因流(Nm)。用GenAlEx 6.5软件计算遗传一致度、遗传距离,并进行分子方差(AMOVA)分析。用NTSYSpc 2.10e软件以及MEGA软件构建非加权组平均法(UPGMA)品种间聚类图。
2 结果与分析
2.1 引物多态性用筛选出的10条引物对65个中华鳖样本进行PCR,扩增结果见表2。共获得123条清晰可见的条带,其中92条条带具有多态性,平均多态性条带(PPB)比例为73.417%,每条引物平均扩增出的条带数为12.3条,多态性条带数为5~15条。引物多态性条带比例为50.00%~93.75%。其中引物UBC854多态性条带比例高达93.75%。图1为用引物UBC807扩增的电泳条带。
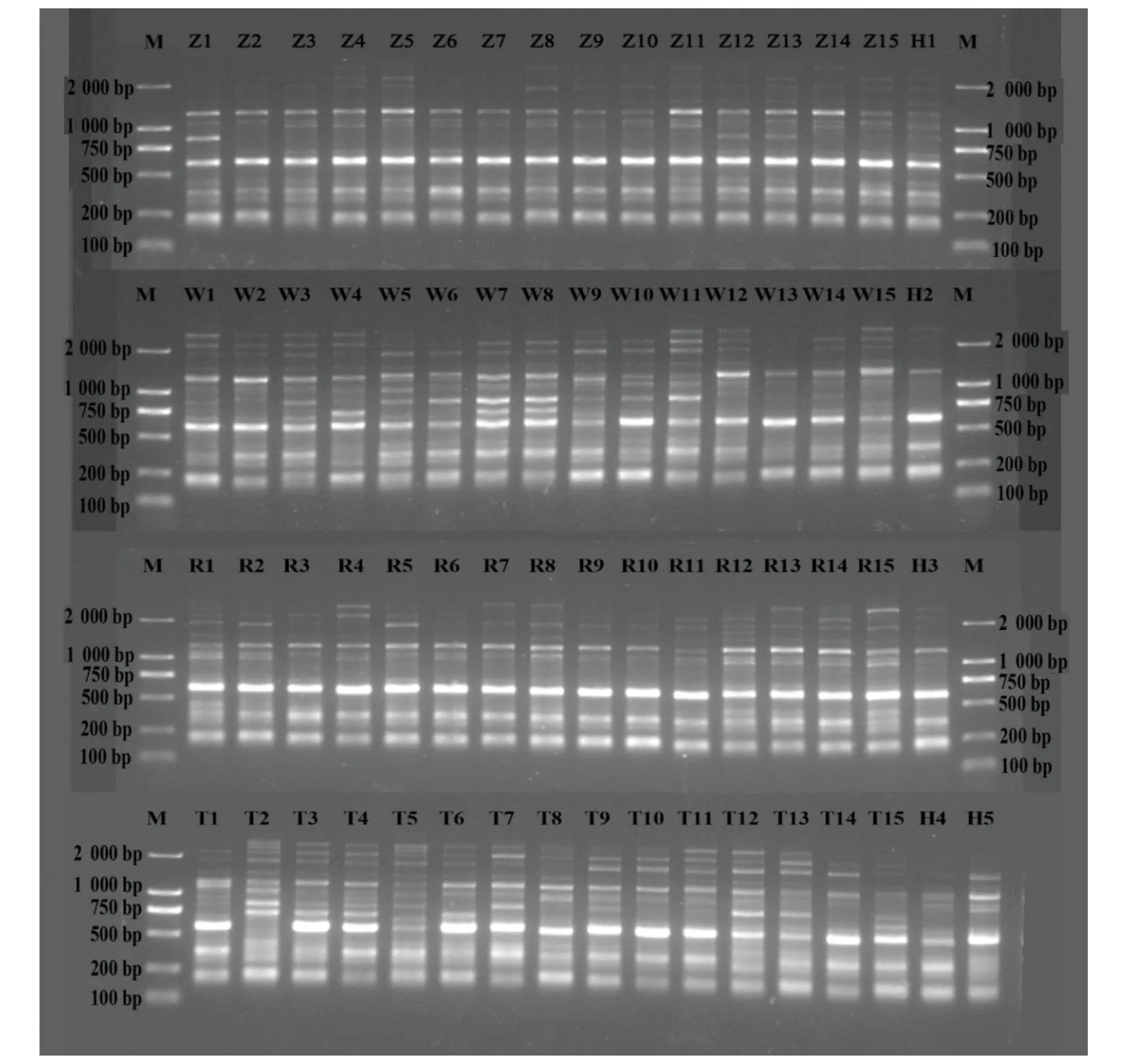
注:Z.杂交鳖;W.乌鳖品系;R.日本品系;T.中国台湾品系;H.淮河品系;M.Marker,DL 2 000。
2.2 遗传多样性5个居群中华鳖的遗传多样性参数见表3。中华鳖5个居群的等位基因数(Na)为1.748 0,有效等位基因数(Ne)为1.417 7,Nei’s基因多样性指数(H)为0.245 5,Shannon多样性指数(I)为0.370 4,多态性位点(PPL)比例为74.80%。在中华鳖5个居群中,乌鳖群体的遗传多样性水平要高于其他中华鳖群体,Na为1.536 6,Ne为1.374 3,H为0.207 5,I为0.302 5,PPB为66,PPL为53.66%。淮河鳖居群的变异程度最小(Na=1.365 9,Ne=1.247 5,H=0.139 2,I=0.204 9,PPB=45,PPL=36.59%)。
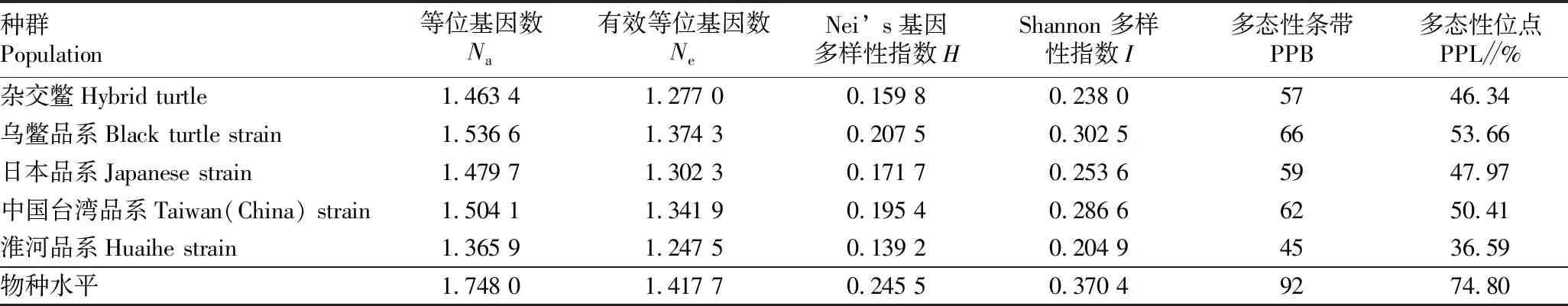
表3 中华鳖5个居群的遗传多样性参数
2.3 居群遗传分化根据Popgen 32的计算结果,居群总遗传多样性指数(Ht)=0.240 0±0.037 1,居群内遗传多样性指数(Hs)=0.174 7±0.021 6,基因分化系数(Gst)=0.271 9,基因流(Nm)=1.339 0。说明有27.19%的遗传变异出现在居群间,有72.81%的遗传变异出现在居群内。分子方差分析(AMOVA)结果与上述Popgen计算结果一致,结果见表4。这说明中华鳖的遗传多样性主要分布在居群内,各居群间表现出一定的遗传分化水平。
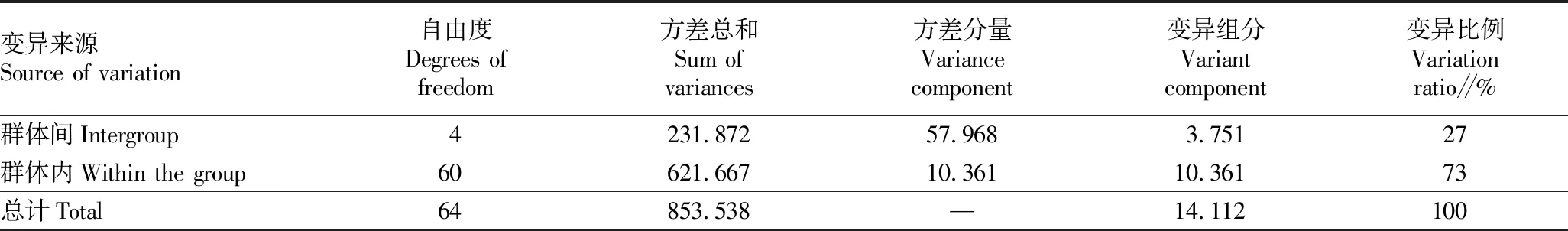
表4 中华鳖5个居群的AMOVA分析
2.4 亲缘关系及聚类结果由表5可知,5个居群的遗传一致度为0.880 3~0.937 1,均值为0.910 1,杂交鳖与日本品系遗传一致度最高;5个品系遗传距离均值为0.943 7,杂交鳖与日本品系之间的遗传距离最小(0.064 9),中国台湾品系和乌鳖品系的遗传距离最大(0.127 5)。从图2可以看出,5个地理居群的中华鳖遗传系数为0.89~0.94,在相似系数为0.89时,分为2个类群,在相似系数为0.93时,可分为3个类群,杂交鳖、日本品系和淮河品系聚为一类群,其中,杂交鳖与日本品系的相似系数最高,可达到达0.94。乌鳖单独为一类群,可见乌鳖品系与其他品系的差异较大。
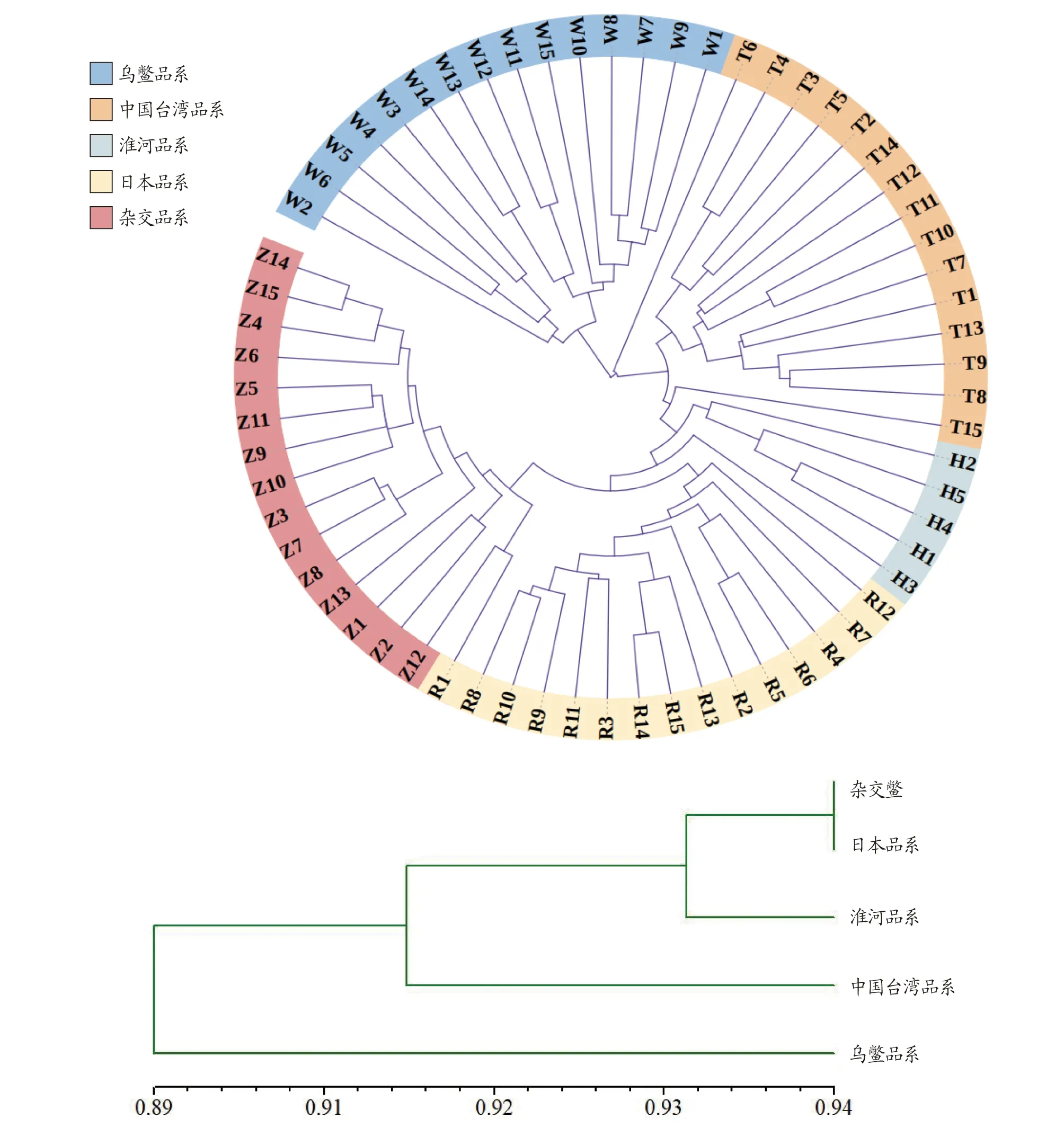
图2 中华鳖5个居群的聚类分析图
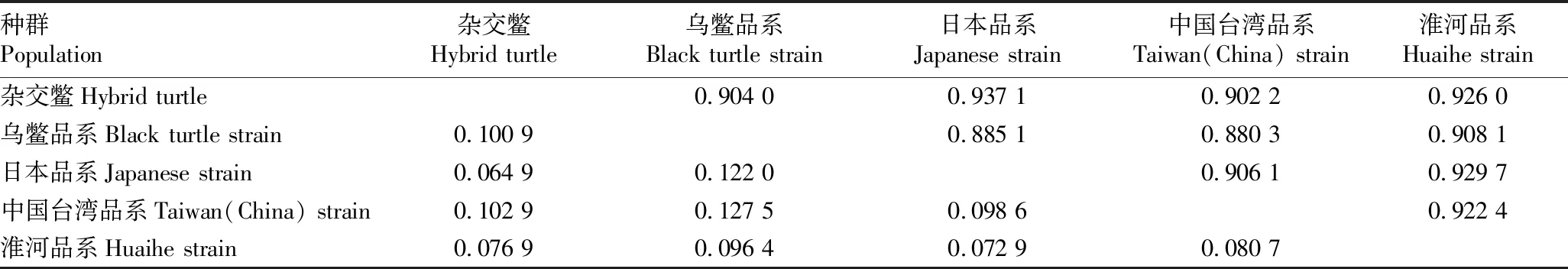
表5 中华鳖5个居群间的遗传一致度(对角线上面)和遗传距离(对角线下面)
3 讨论
中华鳖群体的准确区分一直是育种工作的重点与难点,前人通过对不同品系中华鳖的表型特征进行区分,建立的判别函数可以作为品系区别的重要依据。陆文浩等[18]通过多元统计分析建立了判别方程,其中洪泽湖和太湖群体判别准确率可达100%,长江口群体的判别准确率达93.3%。聚类分析表明,长江口群体与太湖群体亲缘关系很近,洪泽湖群体与它们的亲缘关系相对较远。当品系表型区别较大,地理差距较远时,通过成分分析统计通常能得到较高的判别率。但当亲缘关系越近,或者品系过多、群体形态相似度很高时,传统的形态学分析方法通常难以获得较高的判别准确率。梁宏伟等[19]建立了中华鳖3个品系的判别函数,淮河品系、黄河品系和日本品系的判别准确率分别为91.7%、75.0%和73.8%。李思发等[14]对中华鳖7个养殖群体的判别分类的准确率以黄河鳖群最高(73%),淮河鳖群和太湖鳖群较差(分别为45%、44%);聚类分析显示,在亲缘关系上太湖鳖与崇明鳖较近,黄河鳖和淮河鳖较近,洞庭湖鳖和杂交鳖较近。这表明仅通过分析体形和体表差异进行的比较研究难以保证高的判别准确率。
相比于传统的表型特征成分分析,分子标记不依赖于动物的表型特征,而是根据遗传信息进行区分,即使面对多个表型极其相似的品种也可以很好地将其区分开来。对中华鳖线粒体DNA中SNP位点进行基因分型和区分,共检测到7个SNPs,可用于鉴定太湖品系、日本品系、中国台湾品系和黄河品系[20]。Zhang等[21]对中华鳖的线粒体基因进行了扩增和酶切,建立了太湖、中国台湾、日本和黄河4个品系中国鳖的PCR-RFLP鉴定方法。在该试验中,杂交鳖为日本鳖和江西鳖的后代,外形最为相似,而乌鳖全身乌黑,表型与其他品系相差甚远;聚类分析结果也表明,日本品系与杂交鳖亲缘关系最近,乌鳖与其他鳖亲缘关系最远。同时个体系统发育树的分析结果与聚类分析保持一致,且5个品系的65个个体的中华鳖都得到了准确区分,因此,使用ISSR的分析参数及试验方法对于区分中华鳖品系有高度准确率。
除此之外,分子标记还可获得更多的物种遗传参数,可以反映遗传特征、种群分化和物种间的进化关系。遗传多样性是生物多样性的基本来源以及自然选择进化的原材料,常被用来量化种群内遗传变异性大小[13]。Xie等[22]利用RAD-seq开发中华鳖SNP标记,共鉴定了105个高质量的SNP标记。群体遗传统计显示,观察杂合度(Ho)和期望杂合度值(He)分别为0.114 3~0.676 5和0.107 8~0.447 7。该试验中5个群体He为0.139 2~0.207 4。He越高,反映群体的遗传一致性就越低,其遗传多样性就越丰富。该试验中5个居群中,乌鳖遗传多样性参数最高,且由聚类结果可知,乌鳖品系与其他4个品系的亲缘关系也最远。这表明乌鳖不仅形成了区别与其他品系的中华鳖稳定群体,还具备变异获得更多优良性状的可能。淮河鳖居群的变异程度最小,淮河鳖作为我国淮河流域的一支中华鳖地理种群,其纯合体较为稀少,遗传多样性处于较低水平的原因可能是养殖场累代人工选育、近亲繁殖的结果。另外,杂交中华鳖的遗传特性和经济性状有着更加显著的生长活力,与亲本相比,杂交鳖的特异等位基因数和多态片段数增加[23],表明杂交增加了中华鳖的遗传多样性。
该研究中基因分化系数(Gst)=0.271 9,表明群体间存在高等的遗传分化,分子方差分析(AMOVA)结果表明,有27%遗传变异出现在居群,73%变异发生在居群内部,这说明该研究中华鳖群体间差异不及群体内差异,同一群体内的差异过大,遗传多样性高,性状难以固定。Bu等[24]对3个地理种群(芜湖、安徽大别山、山东聊城)的遗传变异分析也表明,中华鳖的变异主要发生在居群内(97.4%),而不是种群间(2.6%)。基因流(Nm)是指不同种群之间基因交流的过程中,一个等位基因从一个种群进入到另一个种群,可发生在同种或不同种的生物种群之间,是维持种群内遗传同质性和消除种群间遗传分化的重要途径。Feller[25]研究认为,Nm>1时,主要发挥均质化作用,即从其他种群流入的基因会阻止种群间的局部分化,该试验中Nm为1.339 0,这可能也是中华鳖遗传分化程度较低的原因之一。由于养鳖业的快速发展,养殖户对各品系的杂交造成了中华鳖基因的混杂,频繁的基因流和均质化作用使得种群间的遗传分化程度低,该结论与AMOVA分析一致。
综上所述,该试验可以根据所筛选的ISSR引物以及遗传参数分析区分5个中华鳖种群,其中乌鳖遗传多样性最高,杂交鳖和日本鳖亲缘关系最近,群体总体的遗传参数较为丰富,但同时也迫切需要采取保护措施来维持不同种群的遗传稳定性。该研究结果为进一步研究中华鳖的保护生物学、系统发育和进化分析提供了基本的遗传资料,与形态鉴定相比,大大提高了对不同品系中华鳖的鉴别能力,为中华鳖的鉴定和指导中华鳖的选育提供了进一步技术支持。
