Comparison of cell wall changes of two different types of apple cultivars during fruit development and ripening
2023-09-16LIXiangluSUQiufangJIARongjianWANGZidunFUJianghongGUOJianhuaYANGHuijuanZHAOZhengyang
LI Xiang-lu,SU Qiu-fang,JIA Rong-jian,WANG Zi-dun,FU Jiang-hong,GUO Jian-hua,YANG Huijuan, ,ZHAO Zheng-yang,
1 College of Horticulture,Northwest A&F University,Yangling 712100,P.R.China
2 Apple Engineering and Technology Research Center of Shaanxi Province,Yangling 712100,P.R.China
Abstract
Fruit development and ripening is a complex procedure (Malus×domestica Borkh.) and can be caused by various factors such as cell structure,cell wall components,and cell wall hydrolytic enzymes.In our study,we focused on the variations in fruit firmness,cell wall morphology and components,the activity of cell wall hydrolytic enzymes and the expression patterns of associated genes during fruit development in two different types of apple cultivars,the hard-crisp cultivar and the loose-crisp cultivar.In this paper,the aim was to find out the causes of the texture variations between the different type cultivars.Cell wall materials (CWMs),hemicellulose and cellulose content were strongly associated with variations in fruit firmness during the fruit development.The content of water soluble pectin (WSP) and chelator soluble pectin (CSP) gradually increased,while the content of ionic soluble pectin (ISP) showed inconsistent trends in the four cultivars.The activities of polygalacturonase (PG),β-galactosidase (β-gal),cellulase (CEL),and pectate lyase (PL) gradually increased in four cultivars.And the activities of PG,β-gal,and CEL were higher in ‘Fuji’ and ‘Honeycrisp’ fruit with the fruit development,while the activity of PL of ‘Fuji’ and ‘Honeycrisp’ was lower than that of ‘ENVY’ and ‘Modi’.Both four cultivars of fruit cells progressively became bigger as the fruit expanded,with looser cell arrangements and larger cell gaps.According to the qRT-PCR,the relative expression levels of MdACO and Mdβ-gal were notably enhanced.Our study showed that there were large differences in the content of ISP and hemicellulose,the activity of PL and the relative expression of Mdβ-gal between two different types of apple cultivars,and these differences might be responsible for the variations in the texture of the four cultivars.
Keywords: apple (Malus×domestica Borkh.),fruit softening,cell wall,firmness
1.Introduction
The formation of apple (Malus×domesticaBorkh.) fruit texture is a complex process determined by various physiological and biochemical events and is under strict genetic control,and the appearance,flavor and aroma of fruit are also affected during the period of fruit development and ripening (Seymouretal.2002; Giovannonietal.2004; Dheillyetal.2016).And it is a key factor influencing consumer preference for apples and affects the market share of apple cultivars (Hampsonetal.2000; Bowenetal.2019).The ripening of fruit is usually accompanied by softening and the anatomy,physicochemical properties and cell turgor are strongly connected to the decrease of fruit firmness.The softening process involves the breakdown of the primary cell wall and middle lamella polysaccharide polymers,which include a series of pectins and hemicelluloses,among others.In addition,the morphological size,spatial organization,and arrangement of the cells significantly influence the texture and firmness of the fruit (Brummell and Harpster 2001).
Fleshy fruit softening is mainly due to the changes in cell wall structure and breakdown of different cell wall components (Brummell and Harpster 2001; Muñoz-Bertomeuetal.2003).The pectin,cellulose and hemicellulose are randomly populated by hydrogen bonds,covalent bonds and hydrophobic forces and constitute the polysaccharide network structure of the cell wall (pectin-cellulose-hemicellulose) (Jarvis 2011).As for apple,the cell wall of fruit is made of pectin,hemicellulose,cellulose and various structural proteins,like other fleshy fruits (Bennett and Labavitch 2008).According to Giongoetal.(2013),the middle lamella and primary cell wall disintegration,and also a decrease in the content of total pectin are the main causes of the decline in fruit firmness in the process of ripening.Yangetal.(2017) and Dongetal.(2018) measured the water soluble pectin (WSP) in pears and tomatoes and found that the content of WSP in fruits was negatively correlated with fruit firmness suggesting that the content of WSP in fruits could contribute to fruit softening by reducing cell adhesion.In the course of ripening softening studies on bananas and others,softening was found to be connected to hemicellulose modification and degradation,with hemicellulose breakdown serving as the primary trigger for the process of softening of fruit (Chengetal.2009).
The decrease of fruit firmness is achieved through a series of enzymes associated with cell wall degradation,while the reduction of a single enzyme activity usually has almost no influence on fruit softening (Sheehyetal.1988; Smithetal.1990; Goulao and Oliveira 2008).The process involves the coordinated action of cell wall modifying enzymes and proteins including pectin methyl esterase (PME),polygalacturonase (PG),β-galactosidase (β-gal),expansion protein (EXP),pectate lyase (PL) and xyloglucan endoglycosylase/hydrolase (XTH) (Bennettetal.2008; Goulao and Oliveira 2008; Hanetal.2016).Although the depolymerization of pectin is minimal in many species during the early stages of fruit softening,but the correlations between the expression ofPGand softening rate,as well as the analysis of QTL suggest thatPGis involved in fruit softening and texture alterations (Costaetal.2010; Ghianietal.2011).Functional validation of four genes associated with ripening in 14 apple cultivars showed a strong correlation between the expression ofMdPG1and the ripening and softening process (Wakasaetal.2006).WhenPG1was overexpressed in apple plants,the intercellular adhesion of leaves was reduced and when the expression ofPG1was suppressed resulted in firmer fruit (Atkinsonetal.2002).In strawberry (Fragaria×ananassa),the firmness of fruit increased when the expression of ripening associatedFaPGwere repressed indicating thatPGis a crucial role involved in the process of the decrease of fruit firmness (Quesadaetal.2009).Morever,PL,XTH,and β-gal all play significant roles as the fruit ripens (Ngetal.2015; Keetal.2018; Witasarietal.2019).
Ripening of apple fruit is accompanied by ethylene production,respiratory burst and flesh softening (Toivonen and Brummell 2008).It has been studied that the ripening process of fruit is greatly regulated by endogenous plant hormones,including ethylene (ETH),abscisic acid (ABA),methyl jasmonate (MeJA),gibberellin (GA),brassinolide (BR),and salicylic acid (SA) (Kouetal.2021).Ethylene is the primary factor that induces and regulates almost the nearly ripening process in climacteric fruit (Moya-Leónetal.2019).Apple cultivars withACS1andACO1have been shown to be associated with apple fruit firmness (Oraguzieetal.2004; Zhu and Barritt 2008).
In our study,we selected two hard-crisp cultivars ‘ENVY’ and ‘Modi’,and two loose-crisp cultivars ‘Fuji’ and ‘Honeycrisp’,both of which are of excellent quality and have a high market value.We hypothesized that their variability might be directly linked to their cell wall components,structure and developmental period.In our study,we investigated how changes in fruit development,the activity of hydrolytic enzymes and structural traits related to fruit firmness.In order to understand the changes and molecular mechanisms of their fruit growth,we also analyzed the expression profiles of associated enzymes and genes involved in ethylene biosynthesis during their development.
2.Materials and methods
2.1.Plant materials
The test site was located at the experimental station of Northwest A&F University,Baishui County,Shaanxi Province,China.The materials were ‘ENVY’,‘Modi’,‘Fuji’ and ‘Honeycrisp’ cultivars of apple with the same age and growth,grafted on M26 rootstock and planted in 1.5 m×4 m plot.The orchards were subjected to the same environmental factors and management techniques.A total of 20 fruits of uniform size,free of pests and diseases,and free of mechanical damage were picked from the middle layer of the canopy at 60,80,100,120,140,and 160 days after full bloom (DAFB).All apple samples were transported back to the laboratory immediately after picking,and six apple samples were selected from each period for firmness determination and three fruit cut samples were fixed in 70% FAA solution and 4% glutaraldehyde for cell microstructure observation.The remaining samples were frozen using liquid nitrogen and stored at –80°C for subsequent experiments with three biological replicates.
2.2.Measurement of content of sugar components
Add 20 mL of ddH2O to 2 g of liquid nitrogen ground sample,and extracted at 75°C for 30 min.After cooling to ambient temperature,2 mL of supernatant was centrifuged at 12 000 r min–1for 5 min,and then through 0.22 μm filter membrane for determining high-performance liquid chromatography (HPLC).
2.3.Measurement of fruit firmness
The firmness of apple fruit was measured using a FTA GS-15 fruit firmness tester (Germany).The fruits of each cultivar were randomly selected at different developmental periods and were randomly selected at three different locations at the equatorial position for firmness measurement.
2.4.Determination of cell wall degradation related enzymes activity
The enzyme solutions were prepared and the activity of enzymes was measured by the methods of Brummelletal.(2006) and Geetal.(2019),with slight modifications.The activity of PG and CEL was analyzed by DNS colorimetric method and measured at 540 nm and glucose as the standard curve.The activity of PG was performed as the amount of galacturonic acid produced by catalyzing the hydrolysis of polygalacturonic acid per hour per gram of fresh weight fruit,and the activity CEL of was expressed as the amount of reducing sugar formed by catalyzing the hydrolysis of carboxymethyl cellulose per hour per gram of fresh weight fruit.The activity of β-gal was determined by nitrogalactoside hydrolysis and measured at 540 nm.The activity of β-gal was expressed as the amount ofp-nitrophenol material released per hour per gram of fresh fruit catalyzed by the hydrolysis ofp-nitrophenyl-β-glucopyranoside (PNP).The activity of PL was determined by the method of Payasietal.(2004) and measured at 550 nm and galacturonic acid was used as a standard curve for PL activity per unit fresh weight of fruit per unit time.
2.5.Determination of cell wall components content
The cell wall components of apple pulp were separated and their contents were determined by using the different solubility of cell wall fractions according to the method of Melton and Smith (2001).Water soluble pectin (WSP),chelator soluble pectin (CSP),ionic soluble pectin (ISP) and hemicellulose were extracted sequentially,and then the remaining precipitation was cellulose.The carbazoleethanol method was used to test the content of pectin,the anthrone colorimetric method was used to measure the content of hemicellulose,and the weight method was used to test the the content of cellulose.
2.6.Analysis of microstructure of the cell wall
The method of Zhangetal.(2020) was referred to with slight modifications.The pulp was cut into small rectangular pieces of 5 mm×2 mm×1 mm,and the pulp was completely immersed in 70% FAA and fixed at 4°C for 12 h.Then,the samples were repeatedly rinsed with PBS 5 times for 15–20 min each,followed by dehydration with 30,50,70,80,and 90% 1.5 mL ethanol for 15–20 min each,and finally the samples were rinsed again with 100%.The samples were soaked in pure white glue for 48 h.The samples were then packed in transparent capsules containing white glue,and finally the capsules were placed in an oven at 55°C for two days.After drying,the samples were removed from the capsules with a razor blade,and the pulp samples were sliced with a semi-thin slicer (Leica,Germany) at a thickness of 2 μm.The samples were fixed on slides,stained with toluidine blue,washed with pure water to remove excess stain,and allowed to dry before image acquisition under an upright fluorescence microscope (BX51,Olympus,Japan).
Referring to the method of Suetal.(2022) with slight modifications.Longitudinal sample tissues were cut into small rectangular pieces of 5 mm×3 mm×2 mm,fixed overnight at 4°C in 4% glutaraldehyde,rinsed four times by 0.1 mol L–1PBS (pH 6.8) for 10 min each; dehydrated in ethanol of different concentration gradients for 10–20 min each,and 100% ethanol twice for 20–30 min each time; followed by stepwise de-alcoholization in gradient solutions of different concentration tert-butanol for 15 min each,and finally in pure tert-butanol for 30–40 min,freeze-dried for 3 h and sprayed with gold coating before being examined in an S-3400N SEM (FEI Quanta 200,Thermo Fisher Scientific,USA) at 5 kV to observe the fractured surface of apple flesh tissue and acquire images.
2.7.Expression patterns analysis of cell wall de- gradation enzymes related genes
Transcriptomic data were provided by Liuetal.(2021) and were available in the NCBI database repository; the accession number is: PRJNA728501.Samples of four cultivars fruits were taken at different developmental and ripening stages,and total RNA was extracted from apple samples using Quick RNA isolation Kit (Tiangen,Beijing,China).PrimeScript™ RT reagent Kit (TaKaRa) was used for reverse transcription to obtain cDNA,which was stored at –20°C.The primers of qRT-PCR were designed by apple cell wall-associated candidate gene sequences using Primer Premier Software 5.0.The SYBR Green Master Mix (Vazyme,Nanjing,China) and ABI7500 System were used to conduct the qRT-PCR procedures.TheActingene was used as an internal control and the 2-ΔΔCtmethod was used to calculate.The sequences of qRT-PCR primers were provided in Appendix A.
2.8.Statistical analysis
The SPSS 19.0 was used for statistical analysis with one-way ANOVA with Duncan’s test.P<0.05 indicated a significant difference.
3.Results
3.1.Analysis of dynamic changes during fruit develop- ment of two different type cultivars
Apple fruits have a long growth and development cycle.To specifically analyze the changes in firmness of different cultivars with the process of development and ripening of fruit,the measurement of firmness and growth dynamics were performed every 20 days according to the cultivar development cycle.As shown in Fig.1-A,the skin color of four cultivars changed from green to red during fruit development.The fruit development cycle was longer for ‘ENVY’,which took 160 days,while other three cultivars took only 140 days to complete its development.And ‘Honeycrisp’ had larger fruit size compared to ‘ENVY’,‘Modi’ and ‘Fuji’.The fruit texture was affected by fruit firmness.The fruit firmness of ‘ENVY’ and ‘Modi’ was higher than that of ‘Fuji’ and ‘Honeycrisp’ at each stage of fruit development,with the highest firmness of 22.86 kg cm–2of ‘ENVY’ at 60 DAFB.With the development of fruit,the lowest firmness was observed around 160 DAFB of ‘ENVY’ and the remaining cultivars was observed around 140 DAFB (Fig.1-B).Moreover,we also analysed the content of sugar components during the development stages of the four cultivars (Fig.2).From 80 days before harvest to the ripening stage,the contents of glucose,fructose and sucrose of the four cultivars showed an upward trend during the development stage,and the content peaked at the ripening stage (Fig.2-A–C).In general,the glucose and fructose content of ‘Fuji’ and ‘Honeycrisp’ is higher than that of ‘ENVY’ and ‘Modi’,while the sucrose content is higher in the hard-crisp cultivars than that of the loose-crisp cultivars during development and maturation.The content of sorbitol in the four cultivars showed a trend of decreasing first and then increasing (Fig.2-D).During the early stage of development,the sucrose content was lower than that of the glucose content of the four cultivars,but as the fruit matured,the sucrose content was higher than the glucose content during the same period.In addition,the total soluble sugar content of the four apple cultivars during the development period showed an overall upward trend (Fig.2-E).
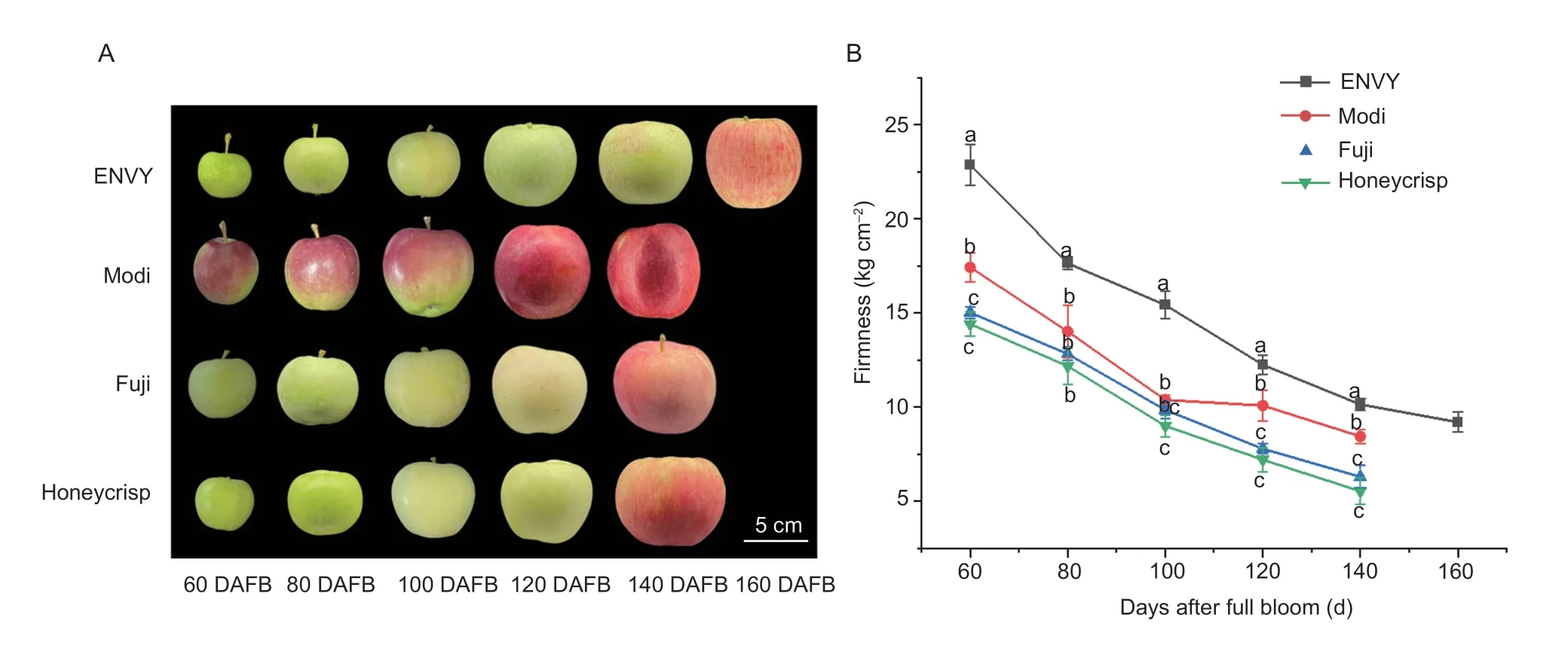
Fig.1 Changes in the appearance and firmness of apple fruits of different cultivars at the stage of fruit development.A,changes in the appearance of ‘ENVY’,‘Modi’,‘Fuji’ and ‘Honeycrisp’ fruits at different stages.B,changes of firmness of four cultivars during development and ripening.DAFB,days after full bloom.

Fig.2 Variations in the content of sugar components of differ cultivars at the stage of fruit development.A,glucose.fruotose.C,sucrose.D,sorbitol.E,total soluble sugar.indicates the stage of maturity.Error bars indicate SD (n=Different lowercase letters in columns indicates significa differences between sampling cultivars for each date Duncan’s test (P<0.05).ent B,M 3).nt by
3.2.Analysis of cell wall degradation enzyme activi- ties during fruit development of four cultivars
In the process of fruit development and ripening,the firmness of two different type cultivars showed consistent trends,but there were differences in firmness between periods.Therefore,we measured the activities of enzymes related to cell wall degradation during development and ripening in four cultivars.The enzyme activities of all four cell wall degradation enzymes showed an increasing trend during the ripening process of two cultivars,especially a significant increase in enzyme activities during the first 40 days before harvest (Fig.3).During the development and ripening,the activity of PG,β-gal and CEL was all high in loose-crisp cultivar compared to hard-crisp cultivar,except for 80 and 60 days before harvest (Fig.3-A–C).In contrast,the activity of PL was higher in ‘ENVY’ and ‘Modi’ fruit than that of ‘Fuji’ and ‘Honeycrisp’,and the difference between the two type cultivars was significant near harvest period (Fig.3-D).At 80 and 60 days before harvest,the activity of β-gal was significantly higher in ‘ENVY’ than in ‘Honeycrisp’,while as it approaches harvest,the activity of β-gal of ‘Honeycrisp’ increases rapidly and was higher than that of ‘ENVY’.The activity of CEL was significantly higher in ‘Fuji’ and ‘Honeycrisp’ at 40 and 20 days before harvest,and the activities of other three enzymes were also significant in the other periods in four cultivars (Fig.3-C).During the ripening period,the activity of PG was significantly different in the four cultivars,especially the activity of PG of ‘Honeycrisp’ was notably higher than that of other three cultivars (Fig.3-A).Interestingly,the activity of PL in different type cultivars showed an opposite trends to PG,β-gal and CEL activities,and the different textures of two type cultivars could be attributed to the activity of PL.And at 40 days before harvest,the PL enzyme activity of four cultivars increased rapidly and the activity peaked at the maturity stage (Fig.3-D).
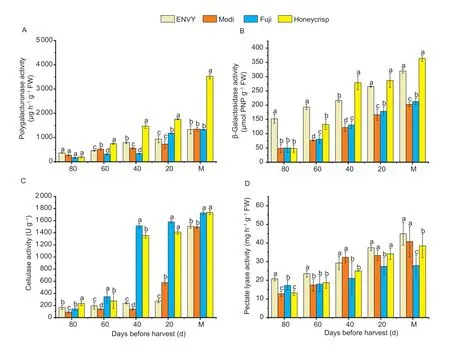
Fig.3 Changes in relevant cell wall degradation enzymes during fruit development and ripening of four cultivars.A,polygalacturonase (PG).B,β-galactosidase (β-gal).C,cellulase (CEL).D,pectate lyase (PL).M indicates the stage of maturity.Error bars indicate SD (n=3).Different lowercase letters in columns indicates significant differences between sampling cultivars for each date by Duncan’s test (P<0.05).
3.3.Changes in cell wall components during fruit development of four cultivars
The decrease of fruit firmness was closely related to the changes of cell wall components and cell turgor.The determination of cell wall degradation enzyme activity revealed that the trend of the activity of PL was significantly different in the two type cultivars.Therefore,in order to further analyze the variations of four cultivars during fruit development and ripening,we determined the contents of water soluble pectin (WSP),chelator soluble pectin (CSP),ionic soluble pectin (ISP) and hemicellulose.From 80 days before harvest to the period of ripening,the WSP content of two different type cultivars showed an overall increasing trend with higher in ‘Fuji’ and ‘Honeycrisp’ than that of in ‘ENVY’ and ‘Modi’.And there is a little difference in WSP content between the four cultivars during the developmental stage but dramatic changes in WSP conetent during the ripening stage and higher in ‘Fuji’ than that of other three cultivars (Fig.4-A).The overall CSP content of ‘Fuji’ and ‘Honeycrisp’ was notably higher than that of the hard-crisp cultivar during ripening,except for the first 20 days before harvest (Fig.4-B).The changes in ISP was more complex than other three cell wall components.With the porcess of fruit development and ripening,the ISP content of ‘Honeycrisp’ showed a trend of decreasing,then increasing,and then decreasing,while the ISP content of ‘ENVY’,‘Modi’ and ‘Fuji’ showed an overall trend of increasing,then decreasing,and the content of ‘Honeycrisp’ was generally higher than that of ‘ENVY’ (Fig.4-C).The contents of the four cultivars differed greatly at the early stage of development and there was basically no significant difference near the maturity stage.The content of cellulose and hemicellulose of four cultivars exhibited comparable decreasing trends in the process of all developmental stages,but the hemicellulose content of ‘Honeycrisp’ and ‘Fuji’ was higher than that of ‘ENVY’ at all stages except for ‘Modi’,while the cellulose content was higher in hardcrisp cultivars than in loose-crisp cultivars and changed significantly in the early developmental stages (Fig.4-D and E).The cell wall material (CWM) was measured from the developmental stage to ripening stage of four cultivars and the CWM content of the four cultivars showed a decreasing trend throughout the developmental stage.The CWM content was higher in ‘ENVY’ and ‘Modi’ than in ‘Honeycrisp’ and ‘Fuji’ with significant differences in the CWM from the early developmental stage to 20 days before harvest.The rate of decline was faster from the fruitlet stage (80 days before harvest) to the expansion stage (60–20 days before harvest),but slower from the late development stage to the ripening stage (20 days before harvest) (Fig.4-F).
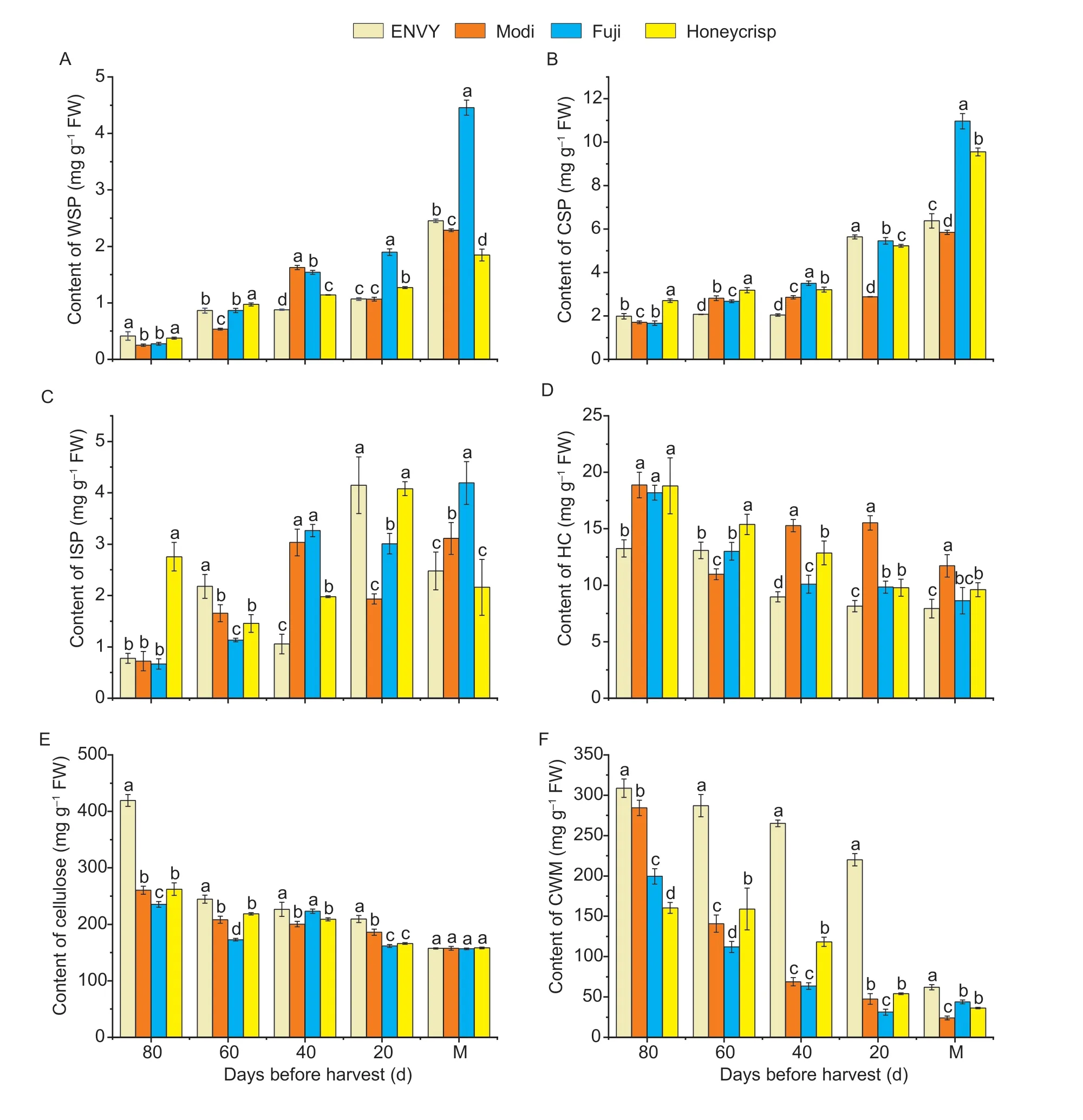
Fig.4 Changes in relevant cell wall components during fruit development and ripening of four cultivars.A,water soluble pectins (WSP).B,chelator soluble pectins (CSP).C,ionic soluble pectins (ISP).D,hemicellulose (HC).E,cellulose.F,cell wall material (CWM).M indicates the stage of maturity.Error bars indicate SD (n=3).Different lowercase letters in columns indicates significant differences between sampling cultivars for each date by Duncan’s test (P<0.05).
3.4.Analysis of the differences in cell microstructure between two different type cultivars during fruit development
During the development of ‘ENVY’,‘Modi’,‘Fuji’ and ‘Honeycrisp’ fruits,the fruitlet,expanding and mature stages were selected according to their respective developmental stages and observed in each stage.From 60 DAFB to the maturity stage,the fruit cells of four cultivars changed from round to long-oval shape,cell arrangement became loose,cell gaps increased,cell number decreased,and cell edges curled but each had differences (Fig.5).At the fruitlet stage (60 DAFB),the fruit cells of both four cultivars were tightly arranged and small in cell size.In the same field of view,the number of cells was higher in the ‘ENVY’ and ‘Modi’ fruit than in the ‘Fuji’ and ‘Honeycrisp’.At the expansion stage,the cells of both cultivars began to increase in size,with the cells of the ‘ENVY’ and ‘Modi’ fruit remaining tightly arranged while the cells of the ‘Fuji’ and ‘Honeycrisp’ fruit began to become irregular in shape and had increased cell spaces.At the mature stage,‘Fuji’ and ‘Honeycrisp’ fruit cells almost disintegrated and the cell edges curled severely.In contrast to loose-crisp cultivar,‘ENVY’ and ‘Modi’ fruit cells were still compactly arranged,but the cell edges also became irregularly curled.The difference in cell structure (size and arrangement) may be one of the reasons for the different textures of the different type fruits.
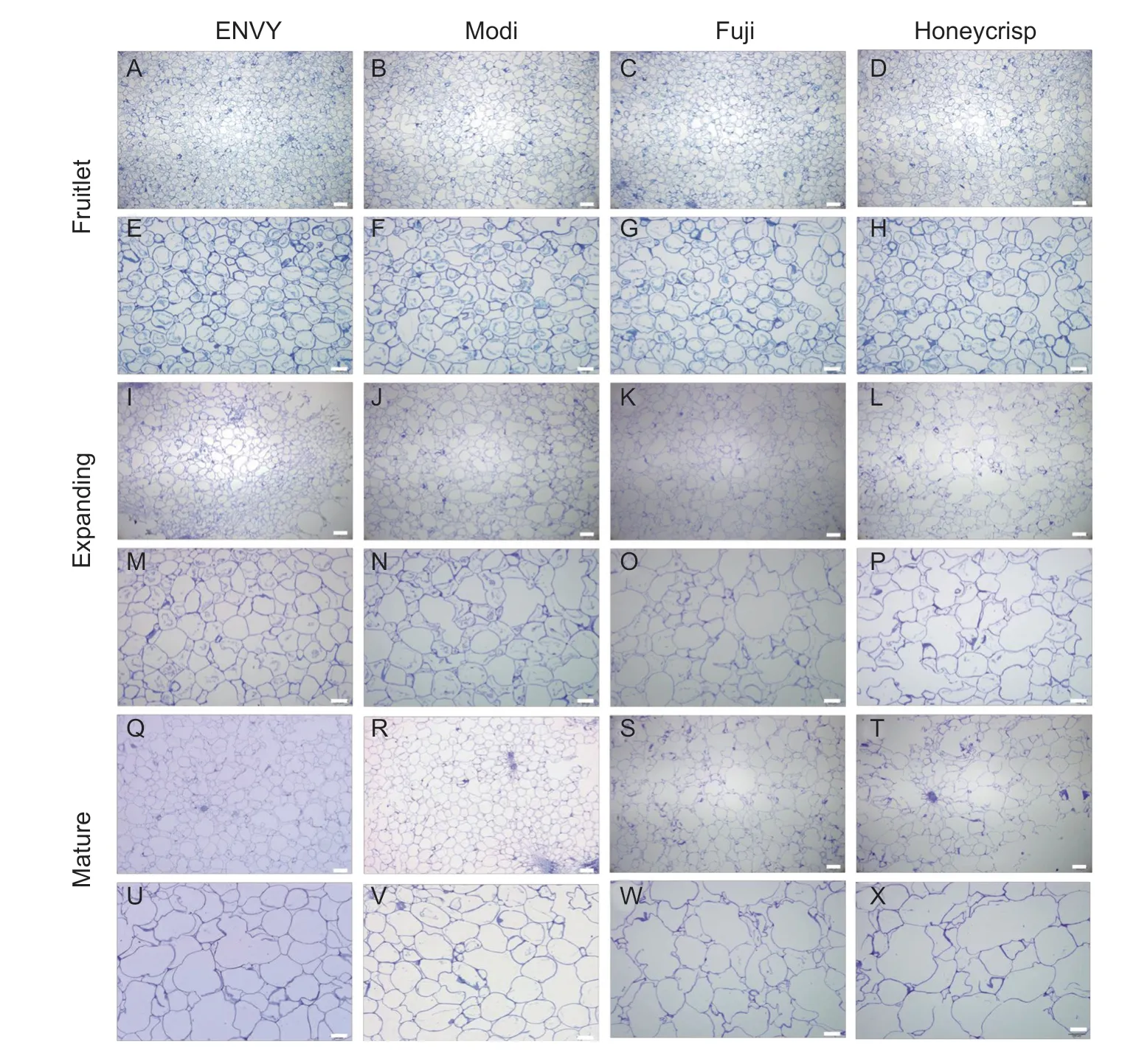
Fig.5 Changes of cell arrangement and size during fruit development and ripening of four cultivars.A–D,I–L,Q–T,obtained at 40× magnification (bar,200 μm).E–H,M–P,U–X,obtained at 100× magnification (bar,100 μm).
By scanning electron microscopy (SEM),we observed the microstructure of fruit flesh tissues of four cultivar fruits at three different developmental stages.As the fruit matured,the thin-walled tissue cells of four cultivars changed from small to large,the cell arrangement became loose and uneven,and the fruit cell walls became thinner with larger cell gaps (Fig.6).At the fruitlet stage (60 DAFB),both four cultivars showed consistent,entire,and distinct honeycomb structures.And the cells of four cultivars were arranged neatly and closely,and intracellular breaks could be observed with small gaps between adjacent cells.In addition,the fractured surfaces of fruit cells of four cultivars were smooth and flat at different developmental stages.As a result,the thickness of the cell wall as well as the size and arrangement of apple fruit cells may have an impact on how the texture changes as the fruit develops.
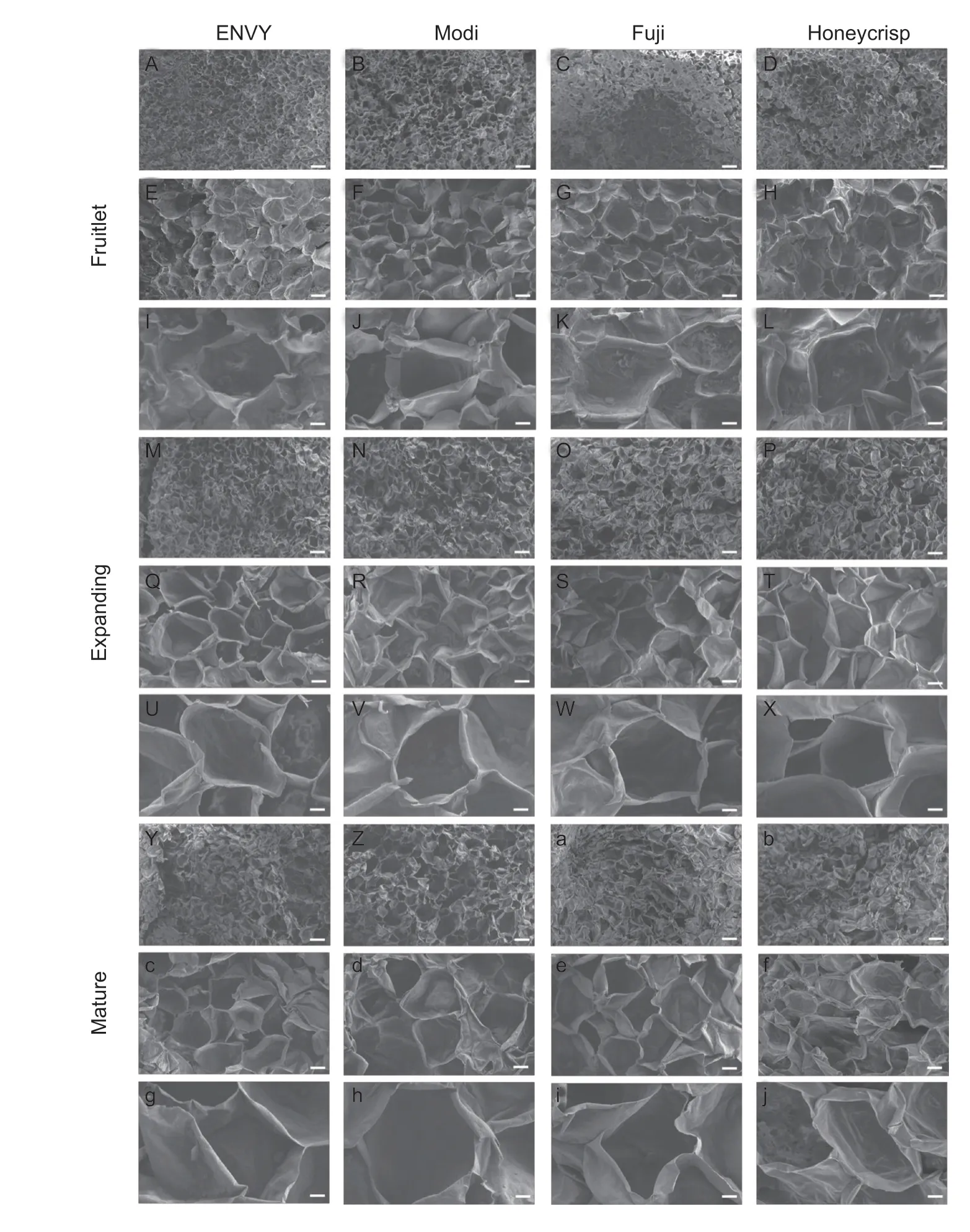
Fig.6 Scanning electron microscopy showing tissue fracture patterns during ripening progress of different cultivars of apple fruit.A–D,M–P,Y–b,obtained at 50× magnification (bar,1 000 μm).E–H,Q–T,c–f,obtained at 200× magnification (bar,200 μm).I–L,U–X,g–j,obtained at 500× magnification (bar,100 μm).
3.5.Expression analysis of cell wall degradation- related enzymes genes during fruit development of four cultivars
Using previous transcriptomic data,cell wall hydrolytic enzymes and ethylene-related genes with significantly altered expression during fruit ripening were screened.The relative expression of eight genes was examined by qRT-PCR during development and ripening in four cultivars (Fig.7).The relative expression of the twoMdPLswas overall higher in the ‘Fuji’ fruit than in the other three cultivars,and the expression trend of the twoMdPLswas reversed.The expression trends ofMdXTHandMdPL(MD16G1070600) were similar,with a decreasing trend from 80 DAFB.The relative expression ofMdPGwas low in the early developmental stages in four cultivars and started to increase slowly in the late developmental stages.The relative expression ofMdEXPgradually increased from 60 DAFB and peaked at maturity in ‘ENVY’ fruits,while the relative expression ofMdEXPin ‘Honeycrisp’ and ‘Fuji’ fruits showed the same trend as that of ‘ENVY’ but decreased at maturity.In addition,the relative expression ofMdACO1showed an increasing trend in three cultivars in addition to ‘Modi’.And the expression ofMdACO1was higher in ‘ENVY’ and ‘Honeycrisp’ than that of ‘Modi’ and ‘Fuji’ at the maturity stage,while the expression ofMdACS1showed a decreasing trend.
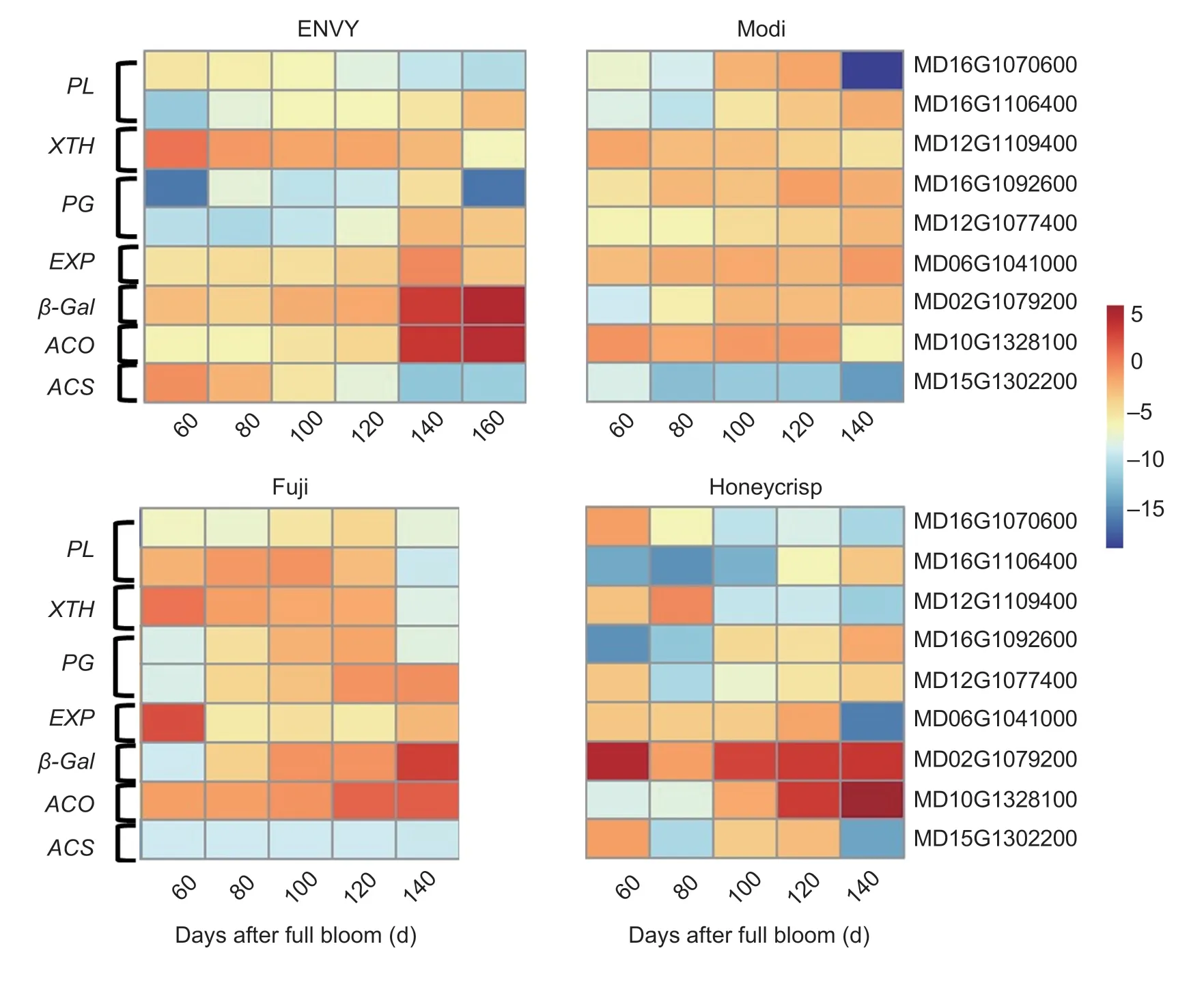
Fig.7 Expression patterns of cell wall degradation enzymes genes of four cultivars.
4.Discussion
4.1.Comparison of changes in firmness and cell wall components during fruit development and ripening in four cultivars
The rate of ripening and softening of apple fruits determines its merchantability and shelf life.The inappropriate levels of firmness or softness of fruits result in diminished shelf life or consumer rejection.Therefore,several species of fruits have experienced selection to improve their palatability and decades of research have explored the causes of fruit softening (Gross and Olsen 2010; Wangetal.2018).In addition,there are numerous studies on the relationship between the cell wall components and fruit firmness,such as strawberry,pear and tomato (Yangetal.2017; Dongetal.2018; Zhangetal.2020).
In the present study,we analyzed the mechanisms regulating the ripening and softening of cell wall components and structures during development in two different types of apple cultivars.In our experiments,we selected two hard-crisp cultivars (‘ENVY’ and ‘Modi’) and two loose-crisp cultivars (‘Fuji’ and ‘Honeycrisp’).The developmental trends of four cultivars were basically the same,but there were also some significant differences.With the development of fruits,the sugar content of the four cultivars showed an increasing trend and the difference in maturity stage was significant.
In terms of fruit firmness,the fruit firmness of ‘ENVY’ and ‘Modi’ was higher than that of ‘Fuji’ and ‘Honeycrisp’,and the fruit firmness gradually decreased with the development of fruit.Previous studies have shown that the content of cellulose and pectin,which make up the cell wall,and the form in which they exist play a key role (Trainottietal.2001).The contents of pectin,cellulose and hemicellulose changes with fruit development and ripening,including the transformation of protopectin into water soluble pectin,degradation of the middle lamella,destruction of the cell wall structure,gradual loosening of the cellulose and hemicellulose skeletal structure,reduction of intercellular adhesion,increase in cell gaps,reduction of fruit firmness,and fruit softening (Brummell and Harpster 2001; Brummell 2006; Lietal.2022).And the most crucial aspect of fruit softening is the convertion of insoluble protopectin (Suetal.2022).In the present study,the contents of WSP and CSP of four cultivars showed an increasing trend in the early stage of fruit development,while the content of ISP first showed an increasing trend,followed by a decreasing trend,and finally an increasing trend again,which is consistent with the previous findings in Zhangetal.(2020).
Moreover,the WSP and CSP contents of the four cultivars differed significantly at the ripening stage.This is probably due to the degradation of protopectin in the early development stage,and then pectin was converted into pectic acid.The variation of ISP content was more different,with the decline in the content of ISP of ‘Honeycrisp’ occurring earlier,around 80 days before harvest and that the content of ISP of ‘ENVY’ occurring 60 days before harvest.And the ISP content of ‘Fuji’ increased notably in the late development.In line with the trend of cell wall material content during fruit development,the contents of cellulose,hemicellulose and CWMs similarly constantly declined with fruit development and ripening.Therefore,the variations causing fruit firmness of the four cultivars in this study strongly connected with the degradation of ISP and hemicellulose.
4.2.The effect of cell wall hydrolytic enzymes during fruit development and ripening in different cultivars
Softening is closely related to the modification of cell wall polysaccharide structure (Fischer and Bennett 1991).Many fruits undergo the programmed process of softening during the maturity stage,which calls for the loosening of the cell walls and the reduction of cell adhesion which the process is made possible by the dissolving of the pectinrich middle lamella (Jarvisetal.2003; Brummell 2006).One of the main components of the cell wall is pectin,which is linked together in the form of multiple acidic hetero-polysaccharides to form a pectin network (Willatsetal.2001).
During the process of ripening,the pectin is degraded by the combined action of a series of pectin-degrading enzymes,the most studied being the PG,followed by studies reporting that PL,β-gal and PME can degrade the pectin,thereby affecting the cell wall integrity (Weietal.2010; Wangetal.2018).The activity of PG enzyme has been reported to be the main cause of fruit ripening and softening in apple (Lietal.2013).The activity of PG enzyme of four cultivars gradually increased with fruit development and ripening process and differed significantly at the ripening stage,which is consistent with previous findings (Lietal.2013).The PL enzyme acts similarly to PG,catalyzing de-esterified pectin cleavage,and the PL enzyme can promote fruit softening in strawberry and tomato (Santiago-Doménechetal.2008; Uluisiketal.2016; Zhangetal.2022).
Our study revealed that the activity of PL enzyme increased gradually,with the highest activity at the maturity stage,and was higher in ‘ENVY’ and ‘Modi’ fruits than in ‘Honeycrisp’ and ‘Fuji’ fruits throughout development and ripening,in contrast to the performance of the other three enzyme activities.As β-gal exacerbates protopectin cleavage,which results in structural alterations of the fruit cell wall and a reduction of fruit firmness,the activity of β-gal increased dramatically with fruit growth,especially during later stages of fruit development.The cellulose can be degraded by CEL,the activity of CEL increased significantly during the development of four cultivar fruits and was higher in loose-crisp cultivars than in hard-crisp cultivars.In this study,the activities of PG,β-gal and CEL were higher in ‘Honeycrisp’ and ‘Fuji’ fruit than in ‘ENVY’ and ‘Modi’,which may be related to the texture more crisp of ‘Honeycrisp’ and ‘Fuji’ fruits.
4.3.Microstructural differences between four cultivars during fruit development and ripening
Previous studies have shown that fruit firmness depends mainly on the cells of size,shape,compactness and the integrity of their cell walls (Huber 1983; Redgwelletal.1997).The microstructural analysis of the fruit revealed that the cells of the four cultivar fruits were significantly smaller and more compact in the early stages of development,and the flesh cells of four cultivars expanded rapidly in the middle and late stages of fruit development and the cells of the ‘ENVY’ and ‘Modi’ fruits were also significantly more over-shaped and neatly arranged than those of ‘Honeycrisp’ and ‘Fuji’.The variations of fruit cell wall structure in apple were strongly associated with the changes in fruit firmness.With the development and maturity of fruit,the cell wall structure changed from dense to loose,and the cell arrangement changed from compact to discrete,and the fruit firmness also decreased.The results of SEM revealed that the thin-walled tissue cells of four cultivars became smaller and larger as the fruit matured,and the cell arrangement became loose and untidy,and the fruit cell walls became thinner and the cell gaps became larger (Fig.6).In addition,the four cultivars belonged to brittle fruit therefore the fracture surfaces of fruit cells of four cultivars at different developmental stages were smooth and flat with no significant differences.
4.4.Expression patterns of ethylene synthesis and cell wall degradation-related enzymes during fruit development and ripening
By qRT-PCR,we could clarify the expression patterns of the cell wall degradation-related enzymes in different cultivars,which is beneficial to investigate the mechanisms affecting fruit firmness and understand the fruit ripening process in depth.Our research revealed that the relative expression ofMdPL(MD16G1106400) gradually increased with fruit development and ripening,while the relative expression ofMdXTHshowed a decreasing trend and the relative expression ofMdPGincreased,indicating that thePGandPLmay be significantly associated with the degradation of pectin and the reduction of firmness.Several studies have demonstrated that thePGandPLare essential for the quick softening of apple,strawberry,and banana (Marín-Rodríguezetal.2003; Quesadaetal.2009; Costaetal.2010).And at the early stage of development,the expression ofPGwas not very high,but the expression ofPGwas enhanced in late development and maturity stages.It has also been shown thatPGacts to the late stage of softening (Costaetal.2010; Ghianietal.2011).The relative expression ofMdXTHwas higher in the early stages of fruit development,suggesting thatMdXTHtakes part in fruit softening during the early stages of fruit development.The relative expression ofMdβ-galof four cultivars was highly upregulated in their late developmental stages,but there were differences in the degree and period of upregulation which could result in differing ripening-stage firmness characteristics.In addition,the transcript levels ofMdACO1,marker genes for fruit ripening,were significantly upregulated upon reaching the ripening stage and differed in the four cultivars.
5.Conclusion
This study provides a broad physiological,biochemical,structural,and molecular basis for fruit softening in four cultivars of apples.Changes in fruit cell structure and arrangement,elevated activity of cell wall hydrolytic enzymes and enhanced expression levels of cell wall hydrolase-related genes occurred with fruit development and maturity.These results suggest that the fruit firmness decreases under the combined effect of these aspects.The two different type cultivars differed in ISP and hemicellulose content,the activity of PL and the relative expression ofMdβ-gal,and these events may be responsible for the differences in fruit texture between the hard-crisp cultivars and the loose-crisp cultivars.
Acknowledgements
This work was supported by the China Agriculture Research System of MOF and MARA (CARS-27).
Declaration of competing interest
The authors declare that they have no conflict of interest.
Appendixassociated with this paper is available on https://doi.org/10.1016/j.jia.2023.07.019
杂志排行

Journal of Integrative Agriculture的其它文章
- Combining nitrogen effects and metabolomics to reveal the response mechanisms to nitrogen stress and the potential for nitrogen reduction in maize
- Natural variations and geographical distributions of seed carotenoids and chlorophylls in 1 167 Chinese soybean accessions
- Carbon sequestration rate,nitrogen use efficiency and rice yield responses to long-term substitution of chemical fertilizer by organic manure in a rice–rice cropping system
- ldentification and epitope mapping of anti-p72 single-chain antibody against African swine fever virus based on phage display antibody library
- Dissecting the genetic basis of grain color and pre-harvest sprouting resistance in common wheat by association analysis
- The PcERF5 promotes anthocyanin biosynthesis in red-fleshed pear (Pyrus communis) through both activating and interacting with PcMYB transcription factors