细胞焦亡与骨质疏松症关系研究
2023-09-12麻永胜蒋宜伟宋重东周玉英刘沛郝东昶罗春晖魏博俞中超
麻永胜 蒋宜伟 宋重东 周玉英 刘沛郝东昶 罗春晖 魏博俞中超
1.甘肃中医药大学,甘肃 兰州 730000
2.甘肃中医药大学附属医院,甘肃 兰州 730000
细胞焦亡属于新型的细胞程序性死亡机制,其形态学特征和分子机制不同于细胞的自噬、凋亡、坏死等[1]。
细胞焦亡激活过程中释放炎症因子及炎症小体,引发局部或全身的炎症反应[2]。适度可控的细胞焦亡有助于机体及时清除病原体。然而,细胞过度焦亡会促发强烈的炎性反应,加剧疾病病理进程[3]。骨质疏松症是临床中一类可发生在全身的骨代谢疾病,多见于老年人和绝经后女性,其特征为骨密度降低,骨量明显减少,临床表现多为腰背部及全身疼痛、脊柱畸形以及容易发生骨折等。相关研究发现[4],全球大约有2亿多名骨质疏松症患者,其中绝大多数的老年人群骨折病例是由骨质疏松引起的,已成为老年人死亡的主要原因之一。近年来研究发现,成骨细胞发生焦亡参与了骨组织的免疫防御反应,过度细胞焦亡的发生将会导致更为严重的骨质损伤。细胞焦亡参与介导OP的发生发展,提示其与骨质疏松症有密切的关系,现就细胞焦亡与骨质疏松症的相关性展开说明。
1 细胞焦亡概述
细胞焦亡是抵御病原体及危险信号的重要防线,它影响机体正常的免疫反应,进而增加疾病发生发展的风险。细胞焦亡现象于1992年被发现,Zychlinsky等[5]在感染福氏志贺菌的巨噬细胞中注意到一种裂解式死亡,发现和细胞凋亡的形态学特征不同。后来Cookson等[6]提出“焦亡”来描述促炎性细胞的程序性坏死,其表现为细胞不断膨胀,然后破裂,从而发挥机体免疫反应,有效的对抗感染。如图1[9]所示,在病原体分子模式(PMAPS)和损伤分子模式(DAMPS)中,细胞焦亡有赖于半胱天冬氨酸蛋白酶(caspase)的激活转导,通过入侵的病原体和内源性刺激,细胞应激反应激活炎性小体而引起细胞焦亡[7]。具体机制是炎性半胱天冬酶(caspase)被激活后转导,效应蛋白消皮素D(gasdermin D,GSDMD)的N端裂解,并激活 proIL-1β和proIL-18(白细胞介素1β和白细胞介素18的前体),通过焦亡信号途径其下游白细胞介素-1β、白细胞介素-18随之释放,并诱导GSDMD的活化成熟,促使孔隙形成和细胞内物质及炎症因子释放[8],引发细胞焦亡。
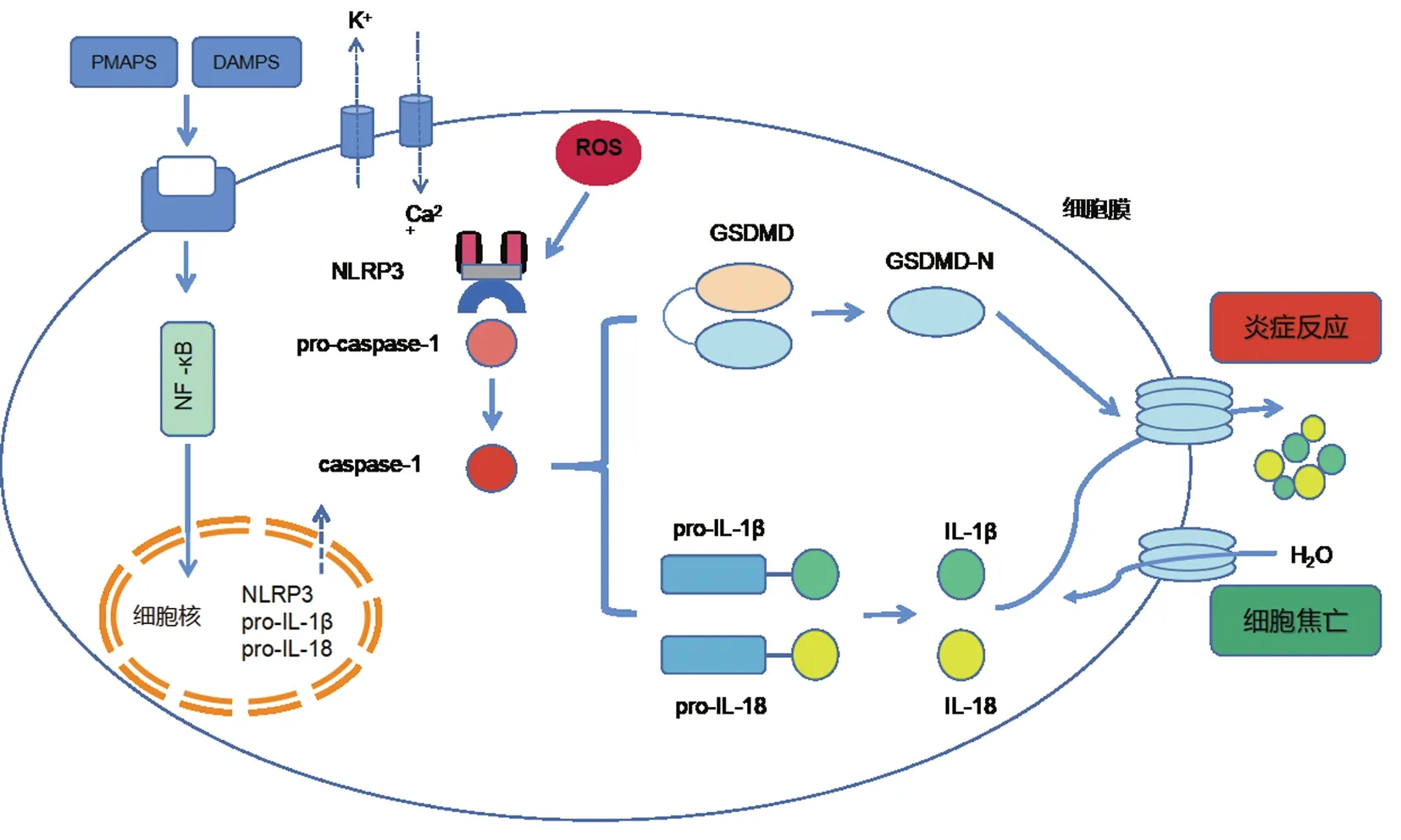
图1 细胞焦亡信号传达图
2 细胞焦亡的信号途径
细胞焦亡的信号途径主要为经典途径和非经典途径。病原微生物感染后,经典途径对病原体和损伤相关分子模式作出反应,而非经典途径对革兰氏阴性菌的细胞内脂多糖 (LPS)作出反应。炎性体作为细胞内模式识别受体,激活caspase[10]。具体来说,经典炎症小体被激活,为多分子蛋白复合物和平台募集caspase-1,导致其自身蛋白水解激活,随后产生成熟的白细胞介素IL-1b和IL-18,导致细胞溶解形式的细胞死亡。同样,非经典炎症小体半胱天冬酶(人类 caspase-4/5和小鼠caspase-11)既可作为传感器又可作为效应器,识别细胞内LPS等刺激物并诱导细胞焦亡[11]。
2.1 经典细胞焦亡途径
首先,在经典途径中,当胞质蛋白检测到危险信号及病原刺激因素后,通过衔接蛋白募集前caspase-1,经活化成为caspase-1,介导细胞焦亡[12]。内源性的分子模式和外源性的病原体模式都能结合识别受体激活细胞焦亡[13]。核苷酸受体蛋白3(NLRP3)作为一种模式识别受体,与炎症密切相关[14]。其中通过病原模式激活核因子κB和丝裂原活化蛋白激酶通路,促进NLRP3、前体白细胞介素IL-1β和IL-18的表达;另一种模式是激活NLRP3后通过衔接蛋白募集pro-caspase-1形成炎症小体,生 成 活 化 的caspase-1,将前体白细胞介素活化成熟,然后裂解GSDMD产生N端结构域,GSDMD-N作用于胞膜形成小孔,细胞渗透性改变,致使其肿胀而细胞膜破裂发生焦亡。
2.2 非经典细胞焦亡途径
细胞焦亡非经典途径是由caspase-4/5/11激活介导的,caspase-4/5/11能被LPS直接激活[15]。LPS存在于革兰阴性菌外膜,进入胞浆后与pro-caspase-4/5/11结合并将其活化,活化后的caspase-4/5/11进而作用于GSDMD的N端,GSDMD-N介导细胞膜裂解,孔隙形成。然后随着细胞膜孔的数量不断增多,一些离子和水分子进入细胞,使得细胞不断膨胀,一旦超过细胞的承受能力,最终引起其肿胀破裂[17]。此外,这一焦亡途径激活NLRP3,导致IL-1β和IL-18外流产生细胞因子并扩大炎症信号反应。与此同时,随细胞焦亡破裂释放的这些炎症因子可以招募免疫细胞,其中IL-18可以刺激T细胞在炎症部位增殖。caspase-4/5/11也可以激活泛连接通道蛋白1(pannexin-1),该蛋白通过排出钾离子激活NLRP3炎症小体,使促炎物质活化并释放[16]。通过GSDMD介导和pannexin-1激活途径将协同发挥细胞焦亡作用。
3 细胞焦亡与骨质疏松症的相关性
细胞焦亡最关键的作用是诱导强烈的炎症反应,帮助宿主防御病原体感染。焦亡途径是重要的药物靶点,其在多种疾病中发挥关键作用,包括但不限于肿瘤、败血症、心脏疾病、糖尿病、肝病、阿尔茨海默病、人类免疫缺陷病毒(HIV)和痛风。除上述疾病外,骨质疏松症与细胞焦亡并没有明确直接的联系,但骨细胞现象都与炎症密切相关。炎症因子IL-1β和IL-18在caspase激活细胞焦亡下参与骨质疏松症的发病机制,NLRP3在炎症诱导下通过细胞焦亡抑制成骨细胞,有些中药影响细胞焦亡相关蛋白的表达水平。因此,细胞焦亡与骨质疏松症的发生发展关系紧密,细胞焦亡会导致骨细胞损伤的加剧,从而出现骨质逐渐疏松并加剧骨折的风险。此外,作为焦亡途径中的重要参与者,NLRP3、GSDMD、IL-1β、IL-18、和caspase 1/4/5/11都是积极追求的药物靶点。
3.1 IL-1β和IL-18参与细胞焦亡介导骨质疏松症的发病机制
IL-1β和IL-18的前体在细胞焦亡期间被caspase-1加工,并与细胞碎片一起分泌到细胞外环境中,从而加重炎症状况。IL-1β和IL-18可直接或间接促进造血干细胞和前破骨细胞分化为成熟破骨细胞,导致破骨细胞过度形成。此外,NLRP3的过度激活可以通过处理IL-1β和IL-18前体来影响破骨细胞的形成,IL-1β和IL-18在破骨细胞生成中产生多种作用,并且在老年骨质疏松症中细胞焦亡对破骨细胞生成的刺激可能更为明显。破骨细胞的形成超过成骨细胞,从而导致过多的骨吸收和骨重塑的不平衡,从而导致骨质疏松症[18]。当机体受到细菌感染或衰老信号时,或者由于激素缺乏,会激活多种炎症通路,炎症因子分泌增加,NLRP3炎症小体放大该信号。还有实验研究用IL-17处理的成骨细胞出现大量毛孔、肿胀和破裂,其中NLRP3炎性体通路参与了成骨细胞的焦亡[19]。Lina Yang等[20]发现高糖通过caspase-1/GSDMD/IL-1β通路激活细胞焦亡,细胞焦亡对牙槽骨成骨细胞增殖分化的抑制作用。成骨细胞发生焦亡,进一步激活破骨细胞,导致骨质流失加重。此外,炎症小体的激活参与了其他全身/局部疾病与骨质疏松症中骨质流失的相互作用。一些研究还发现,IL-1β在许多骨骼疾病中发挥重要作用,例如雌激素短缺下IL-1β的高表达水平通过抑制成骨分化加剧骨质疏松症[21]。综上所述,炎症小体激活诱导的细胞焦亡在破骨细胞和成骨细胞的作用,可能在骨质疏松症中起关键作用。
3.2 NLRP3通过细胞焦亡对成骨细胞的影响机制
NLRP3通过细胞焦亡在炎症诱导的骨质疏松中起核心作用,通过影响成骨细胞直接抑制成骨。McCall SH等[22]证明成骨细胞表达NLRP3,其介导细菌诱导的细胞焦亡并参与炎症期间的骨质流失。许多研究发现骨质疏松症引起的细菌感染,NLRP3的相关表达是必不可少的[23-25]。研究发现NLRP3炎性体在骨组组中表达不断上升可以激活caspase-1细胞焦亡通路,导致成骨细胞死亡[26],这时炎症因子IL-1和IL-18的表达水平也上调。有学者发现雌激素缺乏的骨质疏松症中NLRP3水平也会逐渐升高,对成骨细胞活力影响明显[27],通过灭活NLRP3炎性小体可以促进成骨细胞形成,骨质疏松症也得到缓解[28]。在绝经后骨质疏松症大鼠中也观察到依赖于NLRP3表达的成骨细胞死亡增加,导致小梁厚度、小梁数量、小梁分离和BMD降低[29]。因此,上述发现是NLRP3影响成骨细胞焦亡的证据。体内外实验表明,NLRP3蛋白的过表达与骨吸收加重和成骨缺陷密切相关。还有,NLRP3炎症小体的异常激活不仅会产生炎症,还会通过上调Caspase-1和GSDMD的表达而导致成骨细胞的焦亡和功能障碍[30]。综上,NLRP3炎症小体总体上不仅加速骨吸收,而且抑制骨形成,助长了骨质疏松症的风险,NLRP3激活的细胞焦亡在骨质疏松症的发病机制中发挥着至关重要的作用。
3.3 细胞焦亡治疗骨质疏松症靶向思路
目前临床上用于治疗骨质疏松症的药物主要是经典的抗破骨细胞药物如双膦酸盐或抗成骨的特立帕肽。此外,根据研究Romosozumab治疗可增加骨形成并改善骨密度(BMD),瑞舒单抗和罗莫索单抗在骨质疏松症的治疗中显示出良好的前景,但需要进一步的研究来更好地评估其副作用[31]。该研究强调了细胞焦亡过程中如基本成分GSDMD、NLRP3以及caspase作为药物靶点的潜力。然而,详细的激活和作用机制尚不完全清楚。因此,笔者总结了一些与细胞焦亡有关的重要因素及其与骨质疏松症相关的串扰途径。研究发现,髓系分化因子88 (MyD88)在大鼠中的最大表达可能通过上调MCP-1和RANKL的表达使得破骨细胞增多,但下调MyD88则消除了IL-1的影响,降低了破骨细胞的发生[32]。在链脲菌素诱导的糖尿病大鼠体内,破骨细胞的数量和骨吸收能力显著升高,胫骨近端炎性体相关蛋白MAPK-、NF- ii B-和NLRP3的表达也显著升高,然而,胰岛素和LXA4治疗逆转了这种升高[33]。此外,经Pyrrolidine Dithiocarbamate (PDTC)治疗[34]后,NF-ⅱB表达明显降低,炎症反应减轻,骨吸收过程(破骨细胞形成、脂肪形成和细胞凋亡)明显减少,而在诱导骨坏死后,骨吸收过程明显增强。PDTC还显著增加了被干扰的Wnt/ catenin通路,并提高了骨形成过程。因此,提出NF-ⅱb靶向治疗为一种新的治疗策略。综上,上述分化因子、细胞信号、细胞通路都是潜在的治疗靶点。
一些促炎细胞因子,如 IL-7、IL-12、IL-23和IL-17以及INF-g,已被证明具有双重促破骨细胞和抗破骨细胞特性。似乎它们的净效应在体内取决于研究其骨骼特定病理生理状况,而在体外取决于破骨细胞的发育阶段。其中,IFN-g可通过直接靶向破骨细胞前体抑制破骨细胞生成,但通过刺激T细胞活化和破骨细胞生成因子(TNF-g、RANKL)的产生使破骨细胞形成并且促进骨吸收。在缺乏雌激素和炎症存在的情况下,这两种相反特性的净效应偏向于骨吸收[35]。并且IL-12的双重作用可以被视为IFN-g的相反特性,因为它是IFN-g的有效诱导剂[36]。IL-17处理诱导成骨细胞焦亡,而NLRP3抑制剂阻止其炎性体的激活,从而恢复了成骨细胞的形态和功能。IL-17通过诱导“NF-kappaB (RANK)配体受体激活剂”(RANKL)的表达,刺激破骨骨吸收。此外,IL-17A能够通过减少破骨细胞组织蛋白酶K和相关因子的产生,抑制骨吸收过程中基质蛋白的水解[37]。在OP发生发展过程中,Ac-YVAD-CMK作为caspase-1抑制剂,可抑制caspase-1的激活,从而减轻糖尿病小鼠模型的骨坏死[38]。另外,caspase-1和NLRP3抑制剂显著降低了金黄色葡萄球菌诱导的成骨细胞焦亡,恢复骨形成特性,并在体外减弱骨髓巨噬细胞中的破骨细胞活化并减少体内骨丢失[39]。在葡萄球菌诱导的小鼠骨髓炎模型中,注意到细胞焦亡相关蛋白的表达较高。因此,焦亡相关蛋白的抑制剂在体内和体外都减轻了金黄色葡萄球菌诱导的焦亡。更重要的是,抑制细胞焦亡恢复了骨形成特性,减弱了体外破骨细胞的异常活化,逆转了体内骨损伤。综上,细胞焦亡中存在的其他成分对破骨细胞和成骨细胞分化和生成具有重要作用,寻找和研究相关蛋白抑制剂对于骨质疏松症的靶向治疗具有重要意义。
特别注意的是,近年来中医药抑制细胞焦亡防治疾病的研究不断涌现。研究发现,芍药苷单体衍生物、石斛生物碱、虎杖醇提取物等多种中药有效成分以及如独活寄生汤等中药复方能够降低caspase-1等焦亡相关蛋白的表达水平;人参、鸡血藤,红花、杜仲、女贞子、当归能够抑制 NLRP3与炎症小体的表达抑制细胞焦亡。淫羊藿苷结合MAPK信号通路[40],促进了骨髓间充质干细胞和成骨细胞增殖分化,抑制破骨细胞分化。综上,根据上述治疗OP的中药方剂在细胞焦亡方面的研究,对其进一步的深入研究或许可以为防治骨质疏松提供新的靶点。目前中医药抑制细胞焦亡的研究还处于初步探索阶段,因此,对于具有细胞焦亡靶向作用的中药有待深入研究。
4 总结与展望
综上所述,细胞焦亡与骨质疏松症之间有着密切相关性,细胞焦亡对骨细胞在抵御病原体时发挥的免疫防御和分化形成骨组织的作用极其重要。越来越多的研究证明细胞焦亡在骨质疏松症进展中起着重要作用,研究细胞焦亡介导骨细胞分子机制及其与骨质疏松症的关系,对于探寻新的治疗靶点具有重要意义。目前的研究多通过阻断相关传达信号蛋白及分子(如GSDMD、NLPR3、及IL-17等)来抑制焦亡对骨质疏松症的影响。事实上中医药参与抗细胞焦亡药物治疗以及针对焦亡的靶点在骨质疏松的治疗上有着广阔的发展前景,但研究还处在探索阶段,相关报道文献不足。由于目前相关的文献研究和实验研究不足,关于细胞焦亡介导骨质疏松症的发病机制以及细胞焦亡与骨质疏松症的相关性研究梳理尚不够全面,因此期待后续能够有更多的深入研究来进一步阐明两者的关系,为细胞焦亡机制对骨质疏松症的防治提供依据。
