高温储藏条件对花生油脂和蛋白质品质劣变的影响
2023-08-12李尤好马高兴胡秋辉
李尤好,刘 潇,沈 飞,刘 强,裴 斐,马高兴,胡秋辉
(南京财经大学食品科学与工程学院,江苏省现代粮食流通与安全协同创新中心,江苏高校粮油质量安全控制及深加工重点实验室,江苏 南京 210023)
花生作为全球最重要的油料作物之一,在我国的种植业中占有重要地位,据统计,近年来我国花生播种面积达470万 hm2以上,总产量近1 800万 t,保障了我国植物食用油的安全供给[1]。与高产量随之而来的花生储藏问题一直是人们研究的重点[2]。花生在储藏过程中极易受到环境温湿度、光线、空气组分等外部因素以及自身水分含量、水分活度等内部因素的影响[3-5]而导致氧化、出油、变味等品质劣变,造成花生品质发生变化的主要因素就是花生中含有的油脂和蛋白质[6]。已有的研究表明,较低的环境温度有助于花生品质的保持[7-8]。
花生中油脂平均相对含量高达50%,而其中又有80%为不饱和脂肪酸,不饱和脂肪酸极易被花生仁中的氧化酶类催化发生氧化反应[9-10]。过氧化值能够反映初级氧化产物的含量,酸价则能够反映游离脂肪酸的含量,在油脂的氧化过程中,酸价始终上升,而过氧化值在达到一定峰值后会出现下降的情况[11]。光照、高温、水分和氧化酶类加速了油脂的代谢过程,生成甘油三酯和游离脂肪酸,游离脂肪酸进一步被氧化成氢过氧化物,并裂解成小分子的醛、酮、酸类物质,从而造成油脂氧化程度上升、风味劣变和品质下降[12]。
花生蛋白质平均相对含量可达30%,花生在储藏过程中,其蛋白质的氧化除了会导致蛋白质功能特性下降、风味改变,更会引起花生营养损失[13-14]。而花生品质劣变的现象在炎热的夏天尤为明显[15],研究表明,温度高于20 ℃时,温度越高,花生内源酶活性越高,从而加剧花生仁中油脂酸败、蛋白质氧化[16]。此外,蛋白质在与脂质氧化产物接触后,会以化学键的形式与其结合形成脂蛋白复合体,从而导致蛋白质氧化程度加剧[17];同时,也有部分研究表明,蛋白质可以通过金属螯合和自由基清除作用抑制脂质的氧化[18],因此,脂质的氧化同时也是影响蛋白质氧化的因素之一。
本实验通过对花生仁在15、25、35 ℃条件下进行模拟储藏,测定30 周储藏时间内氧化酶、抗氧化酶活力以及油脂和蛋白质相关氧化指标,探究储藏温度对花生仁储藏过程中油脂和蛋白质氧化程度的影响。
1 材料与方法
1.1 材料与试剂
‘四粒红’花生,原产地山东德州,采购自南大和园农贸市场。
正己烷、异丙醇、可溶性淀粉、冰乙酸、盐酸、Tris-Gly缓冲液、聚乙烯吡咯烷酮(polyvinyl pyrrolidone,P V P) 上海阿拉丁生化科技股份有限公司;乙醚、三氯甲烷 南京化学试剂股份有限公司;百里香酚酞、二硫苏糖醇(dithiothreitol,DTT)溶液(1 mol/L)、碘化钾、5,5’-二硫代双(2-硝基苯甲酸)(5,5’-dithiobis-(2-nitrobenzoic acid),DTNB)上海麦克林化学试剂有限公司;氢氧化钠、硫代硫酸钠、无水硫酸钠 西陇科学股份有限公司;三氯乙酸、95%乙醇、尿素 广东光华科技股份有限公司;以上试剂均为分析纯。
1.2 仪器与设备
DF-101S集热式恒温磁力搅拌器 上海力辰邦西仪器科技有限公司;HWS型恒温恒湿箱 宁波东南仪器有限公司;LABCONCO冷冻干燥机 北京照生行仪器设备有限公司;GL-21M离心机 上海市离心机械研究所有限公司;Hei-VAP ML Adv/Pre(EU)旋转蒸发仪 海道尔夫仪器设备(上海)有限公司。
1.3 方法
1.3.1 花生的处理
分别称取带壳花生3 份,每份(7.5±0.1)kg,装入45 cm×32 cm×21.5 cm收纳盒中,分别置于25、35、45 ℃和50%相对湿度的恒温恒湿箱,每隔3 周取样进行各指标测定。
1.3.2 花生酶类活力的测定
脂肪酶活力参照GB/T 5523—2008《粮油检验 粮食、油料的脂肪酶活动度的测定》进行。
脂肪氧化酶活力参照Zhang Shen等[19]的方法测定。取0.1 g花生粉末,加入20 mL含质量分数1% PVP的磷酸盐缓冲液(50 mmol/L、pH 7.0),于4 ℃下10 000 r/min离心15 min,取上清液。取2.875 mL 0.1 mol/L乙酸-乙酸钠缓冲溶液(pH 5.5),加入25 μL 10 mmol/L亚油酸钠溶液,35 ℃保温10 min后,加入0.1 mL上述上清液,混匀。以蒸馏水为空白对照,测定234 nm波长处吸光度,每30 s记录一次,以每分钟吸光度变化0.01为1 个脂肪氧化酶活力单位U,结果以U/g表示。
过氧化物酶的活性使用过氧化物酶测定试剂盒进行检测。
1.3.3 花生油脂、蛋白质制备
花生油脂制备参照GB 5009.229—2016《食品安全国家标准 食品中酸价的测定》方法进行。
将花生脱脂后样品风干制成脱脂花生粉,参照杨曦等[17]的方法并有所改动,脱脂花生粉按料液比1∶10加入水,磁力搅拌2 h,用2 mol/L NaOH溶液调节pH值至8.5,经室温提取1 h后,于4 ℃、6 000 r/min离心两次,每次15 min,所得上清液用2 mol/L HCl溶液调节pH值至4.5后经上述条件离心,所得沉淀调至中性,冻干即得到花生蛋白。
1.3.4 花生油脂氧化指标测定
酸价参照GB 5009.229—2016《食品安全国家标准 食品中酸价的测定》进行测定;过氧化值参照GB 5009.227—2016《食品安全国家标准 食品中过氧化值的测定》进行测定;丙二醛(malondialdehyde,MDA)含量使用MDA测定试剂盒测定。
1.3.5 花生蛋白质氧化指标测定
羰基含量使用蛋白质羰基含量测定试剂盒测定。
游离巯基和二硫键含量参照Beveridge[20]、Zhao Xiaoyan[21]和彭武[22]等的方法并有所改动,取花生蛋白0.1 g加入30 mL去离子水,磁力搅拌2 h后定容至50 mL。
取1 mL蛋白溶液溶解于5 mL Tris-Gly-尿素(尿素8 mol/L)缓冲液中,涡旋振荡后加入50 μL DTNB溶液(4 mg/mL)中,在25 ℃条件下反应1 h,测定412 nm波长处吸光度。按式(1)计算游离巯基含量。
式中:CSH为游离巯基含量/(μmol/g);A412nm为412 nm波长处吸光度;ρ为样品质量浓度/(mg/mL);D为稀释因子。
取1 mL上述蛋白溶液于3 mL含有10 mmol/L DTT的8 mol/L尿素溶液中,涡旋振荡后于25 ℃条件下反应1 h,添加6 mL体积分数为12%的三氯乙酸溶液后于25 ℃条件下再反应1 h,将样品于4 ℃、10 000 r/min条件下离心10 min。将沉淀加入到9 mL 8 mol/L尿素溶液和90 μL DTNB(4 mg/mL)溶液中混匀,测定412 nm波长处吸光度。按式(2)、(3)分别计算总巯基含量和二硫键含量。
式中:ρ为样品质量浓度/(mg/mL);CSS为二硫键含量/(μmol/g);CSHT为总巯基含量/(μmol/g);CSH为游离巯基含量/(μmol/g)。
1.4 数据处理与分析
采用SPSS Statistics 25软件处理数据,结果以平均值±标准差表示,采用Origin 2018软件作图。
2 结果与分析
2.1 花生储藏过程中酶活力变化
2.1.1 脂肪酶和脂肪氧化酶活力变化
花生中的氧化酶和抗氧化酶类活力在不同温度下呈现不同的变化趋势,由此直接或间接地影响花生中脂肪和蛋白质的氧化。不同温度储藏条件下花生脂肪酶和脂肪氧化酶活性变化如图1、2所示。对于脂肪酶,15 ℃组在第3周时活力达到峰值((41.60±1.23)U/g),此后该组脂肪酶活力始终保持较低水平,这是因为低温能有效抑制脂肪酶的活性[23]。25 ℃组脂肪酶活力在第3周和第9周分别达到两个峰值((40.39±1.12)U/g和(40.82±0.70)U/g),随后下降,第12周起从(35.03±1.01)U/g开始稳步上升,并在实验结束时达到(47.12±1.34)U/g,超过了实验初期的两个峰值,且上升的趋势并未放缓,与35 ℃组脂肪酶活力差距逐渐减小。35 ℃组脂肪酶活力在第3周达到第一个峰值((44.93±1.03)U/g),并自第6周起活力保持上升趋势,在实验结束时35 ℃组脂肪酶活力达到(59.00±1.70)U/g,显著高于实验初期该组脂肪酶活力达到的峰值,这是因为较高的温度可以提高脂肪酶的活性,并且随实验时间推移,脂肪酶的活性始终保持上升趋势。对于脂肪氧化酶,在15 ℃储藏条件下,样品的脂肪氧化酶活力在第18周达到峰值((1 036.14±34.49)U/g),此后样品的脂肪氧化酶活力始终保持较低水平。35 ℃组样品的脂肪氧化酶活力第18周达到峰值((1 287.17±98.45)U/g),在第18周后脂肪氧化酶活力呈下降趋势,直至实验结束时脂肪氧化酶活力下降至(1 009.62±42.39)U/g,其原因是长时间储藏于高温条件下会钝化酶,从而导致活性大幅下降[24],对于25 ℃组来说,样品脂肪氧化酶活力自开始储藏后总体呈上升趋势,并在第24周达到峰值((1 265.19±32.50)U/g),最终在实验结束时降至(1 009.62±42.39)U/g。综合分析3 种储藏温度条件下样品脂肪氧化酶活力的变化过程可知,较高的储藏温度会提高脂肪氧化酶的活性,但高温也会破坏酶的活性中心,导致储藏后期酶活性的下降,低温同样能抑制脂肪氧化酶的活性。脂肪酶调控脂肪的氧化速率,催化脂肪水解生成大量的游离脂肪酸,使油脂酸败、酸价升高、油脂风味变差。脂肪氧化酶会催化多不饱和脂肪酸氧化生成氢过氧化物,氢过氧化物会促进蛋白质等组分的氧化,同时自身会分解成其他挥发性风味物质,影响花生的风味[13],二者的活性变化会对花生储藏过程中的油脂和蛋白质氧化进程产生影响。
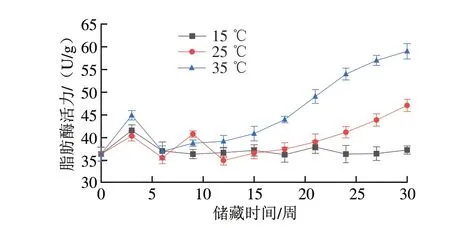
图1 不同温度条件下储藏花生脂肪酶活力变化Fig.1 Changes in LPS activity of peanut stored at different temperatures

图2 不同温度条件下储藏花生脂肪氧化酶活力变化Fig.2 Changes in LOX activity of peanut stored at different temperatures
2.1.2 过氧化物酶活力变化
过氧化物酶可将脂质的部分二次氧化产物转化为无害的物质,由此清除积累的过氧化物,抑制花生品质的劣变[25]。由图3可知,15、25、35 ℃组过氧化物酶活力均在第3周达到峰值,分别为102.3、104.5 U/g和107.1 U/g,25 ℃组和35 ℃组过氧化物酶活力在达到峰值后之后呈下降趋势,15 ℃组过氧化物酶活力在第12周后不再发生明显变化。其原因是花生籽粒作为活的生命有机体,在面对非生物胁迫时,活性氧等指标水平显著上升,在此情况下籽粒自身会启动防御响应来抵御活性氧对细胞造成的损伤,包括酶类和非酶类的活性氧清除系统,其中酶类的防御响应主要包括超氧化物歧化酶、过氧化物酶、过氧化氢酶等抗氧化酶类活性的升高,非酶类的防御响应主要包括植物体内的多种维生素、渗透调节物质含量提升以及如鞘脂代谢增强[26-28]等,因此在温度胁迫下,储藏初期花生的过氧化物酶活力上升。部分研究表明,在长期储藏过程中,植物的抗氧化酶类活性总体呈下降趋势,且温度越高,活性下降越明显,低温能够有效抑制过氧化物酶活性的降低。在本实验中,第3周过氧化物酶活力上升是由于受到温度胁迫时植物体启动了防御响应,而在长时间储藏过程中,25 ℃组和35 ℃组由于自由基过量积累和细胞衰老等原因[29],抗氧化酶活性无法长期保持较高的水平,因此在储藏中后期活性明显降低。
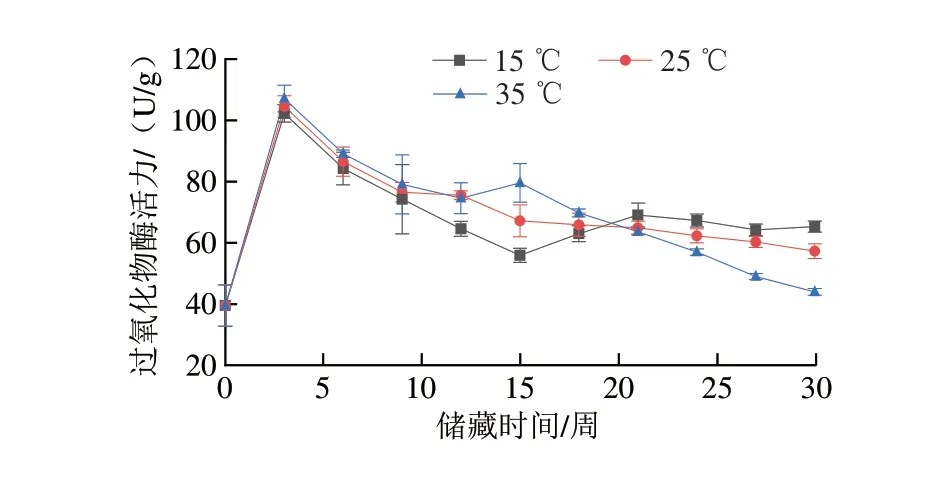
图3 不同温度条件下储藏花生过氧化物酶活力变化Fig.3 Change in POD activity of peanut stored at different temperatures
2.2 花生储藏过程中油脂氧化指标变化
氧化劣变的程度通常可用过氧化值、酸价以及MDA含量等指标量化。过氧化值能够反映初级氧化的程度,酸价则被用来评估小分子的醛、酮、醇类等二次氧化产物水平,而MDA是脂质过氧化的主要产物,其含量可以反映脂质过氧化的程度。在50%恒定相对湿度,15、25 ℃和35 ℃储藏温度下,储藏30 周过氧化值、酸价以及MDA含量的变化分别如图4~6所示。在整个储藏过程中,不同温度条件下的样品各指标均随储藏时间延长而升高,且上升幅度与温度有密切联系,高温环境下储藏的样品各指标上升幅度显著高于低温环境下储藏的样品。储藏30 周后,储藏于35 ℃条件下的花生仁酸价从储藏初期(0.48±0.01)mg/g上升至(3.15±0.10)mg/g,增加了5.6 倍,储藏于15、25 ℃条件下的花生仁酸价同期分别上升至(0.99±0.09)mg/g和(1.78±0.02)mg/g,增加了1.06 倍和2.71 倍。储藏于35 ℃条件下的花生仁过氧化值从储藏初期0.14 mmol/kg上升至7.75 mmol/kg,增加了54.4 倍,储藏于15、25 ℃条件下的花生仁过氧化值同期分别增加了18.4 倍和28.6 倍。同时,储藏于35 ℃条件下的花生仁MDA含量从从储藏初期的23.03 nmol/g上升至1 039.63 nmol/g,增加了44.1 倍,储藏于15、25 ℃条件下的花生仁同期分别增加了25.0 倍和35.8 倍。花生仁中油脂经脂肪酶的催化能分解成甘油和游离脂肪酸,脂肪酸进一步分解成低级的醛、酮类化合物,对水稻、核桃等粮油作物的研究均表明高温引起的氧化酶类活性升高是导致花生酸价上升、风味劣变和品质降低的原因[30-32]。综上,高温对脂肪氧化的影响一方面是加速脂肪碳链受热断裂,另一方面是加速酶类对脂类的水解,产生更多游离脂肪酸[33],而低温储藏条件除了抑制氧化酶类的活性,还减少了抗氧化物质如VE的消耗[8],起到了减缓花生油脂氧化的作用。
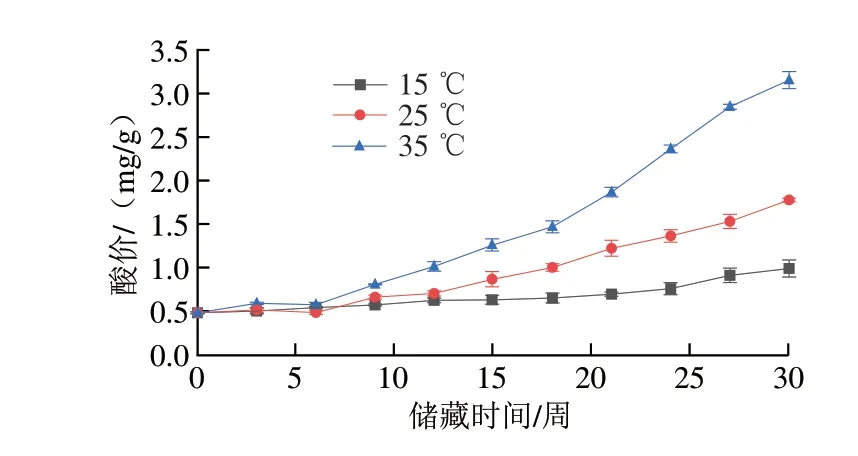
图4 不同温度条件下储藏花生酸价变化Fig.4 Changes in AV of peanut stored at different temperatures
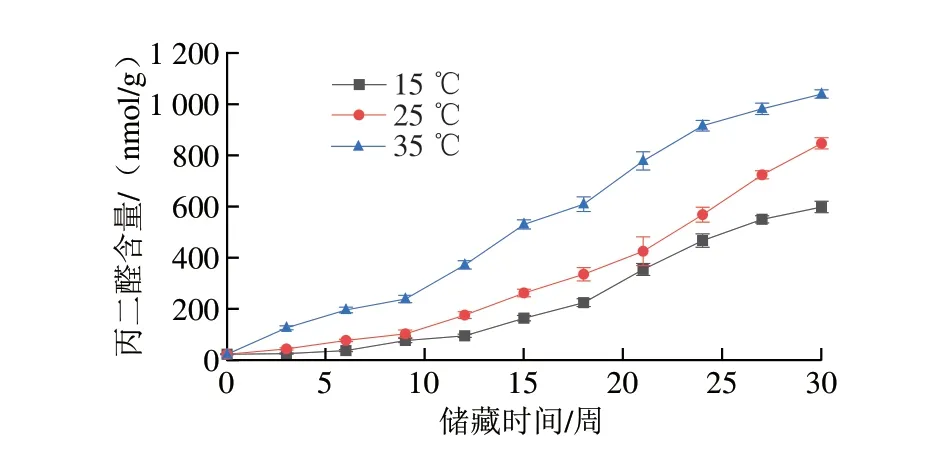
图6 不同温度条件下储藏花生MDA含量变化Fig.6 Changes in MDA content of peanut stored at different temperatures
2.3 花生储藏过程中蛋白质氧化指标变化
2.3.1 羰基含量变化
蛋白质氧化反应可分为直接反应和间接反应。蛋白质可以被活性氧直接诱导发生主肽链或侧链基团氧化,或者被脂质过氧化反应产生的活性中间产物诱导而发生氧化[34]。由图7可知,不同样品在30 周的储藏周期内羰基含量均随储藏时间延长而升高。第30周,储藏于35 ℃条件下的花生仁羰基含量从储藏初期的(3.19±0.24)μmol/g上升至(124.86±3.07)μmol/g,增加了38.1 倍,储藏于15、25 ℃条件下的花生仁羰基含量同期分别上升至(57.78±2.06)μmol/g和(118.61±6.41)μmol/g,增加了17.1 倍和36.2 倍。蛋白质氨基酸侧链的直接氧化、肽骨架的断裂和还原糖反应或者和非蛋白羰基化合物相结合都可以产生羰基[35]。高温条件下氧化酶的高活性催化了油脂的氧化,也使反应继续向生产羰基化合物的方向进行,同时,由于储藏后期35 ℃组脂肪氧化酶活性下降,而25 ℃组脂肪氧化酶活性持续上升,二者的羰基含量依然保持上升趋势并逐渐接近。
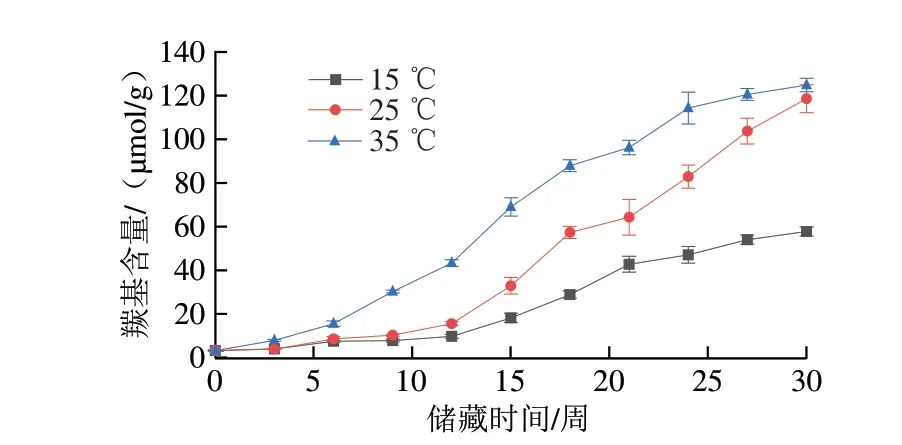
图7 不同温度条件下储藏花生羰基含量变化Fig.7 Changes in carbonyl content of peanut stored at different temperatures
2.3.2 巯基与二硫键含量变化
二硫键和巯基是稳定蛋白质分子构象的重要化学键,蛋白质游离巯基和二硫键含量的变化与蛋白质中的活性基团特别是半胱氨酸残基密切相关,氧化可以改变半胱氨酸的氧化还原状态,进而改变蛋白质分子中巯基和二硫键的含量和分布[22],在蛋白质氧化中,巯基会被转变为二硫键[36]。因此,游离巯基与二硫键含量可以作为评价蛋白质氧化程度的重要指标。由图8、9可知,不同样品在储藏过程中游离巯基含量明显下降,同时二硫键含量明显上升。储藏30 周后,各组花生仁游离巯基含量较储藏初期下降了72.5%~83.1%。储藏于35 ℃条件下的花生仁二硫键含量从储藏初期的37.98 μmol/g上升至65.14 μmol/g,增加了0.72 倍,储藏于15、25 ℃条件下的花生仁游离巯基含量同期分别增加了0.63 倍和0.41 倍。出现上述现象的原因是在储藏过程中蛋白的结构被破坏,暴露出更多易氧化的半胱氨酸及其残基,同时游离巯基被氧化为二硫键,导致游离巯基含量的减少和二硫键含量的上升。但随着样品中各氧化酶类活性的升高,大量的游离巯基被转化为二硫键,这一现象在35 ℃组中尤为明显。
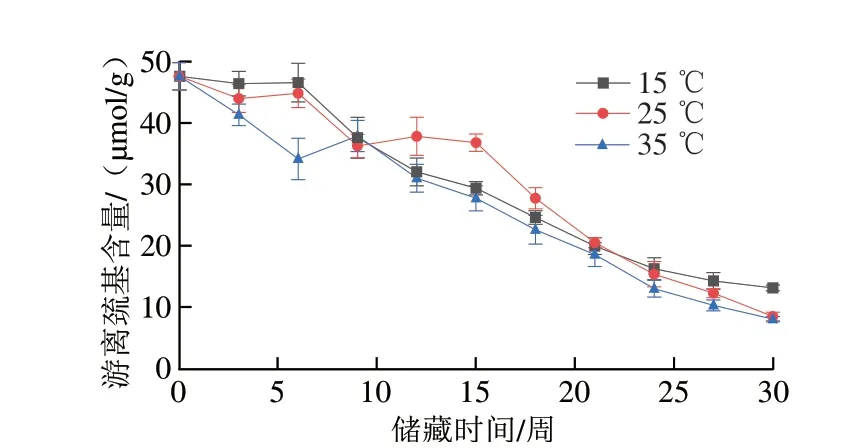
图8 不同温度条件下储藏花生游离巯基含量变化Fig.8 Change in free sulfhydryl content of peanut stored at different temperatures
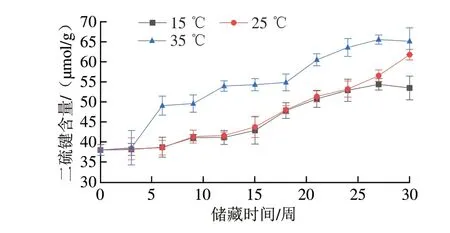
图9 不同温度条件下储藏花生二硫键含量变化Fig.9 Changes in disulfide bond content of peanut stored at different temperatures
除了温度、酶类对花生蛋白质氧化产生影响以外,油脂的氧化同样对蛋白质氧化产生影响,油脂和蛋白质的共氧化是近几年新兴的研究方向,由于脂类和蛋白质之间存在着复杂的相互作用[37],因此二者的共氧化机制还在研究中。目前对共氧化的研究方法主要有3 种,以研究蛋白质氧化为例:第一种方法是去除样品中的油脂成分,探究在无油脂存在的情况下,蛋白质的氧化情况[38-39];第二种方法是抑制样品中油脂的氧化,比如通过添加抗氧化剂的方式,探究在油脂氧化被抑制的情况下蛋白质的氧化情况[40];第三种方法是将蛋白质置于模拟的脂类氧化条件下探究蛋白质的氧化规律[17]。在脂质过氧化反应中产生的脂质氢过氧化物通过一定的条件可转化生成活性羰基化合物[41],蛋白质侧链基团可与活性羰基化合物反应产生共价交联物[22],其中最为常见的是活性醛氧化蛋白质,上文中提到的MDA就是典型的活性醛类化合物。这一过程在一些植物性原料中已被证实,如尤翔宇发现脂质过氧化对米糠蛋白的结构和功能特性等存在影响[36],但该方向的研究大部分集中于肉类蛋白,对于植物蛋白与油脂氧化的相互作用研究还待进一步深入。这些因素对蛋白质氧化的促进作用导致了蛋白质风味改变、营养缺失和功能特性的改变。
3 结论
本研究结果表明,脂肪酶和脂肪氧化酶的活性以及活性变化速率受温度影响较大,高温可以提升脂肪酶和脂肪氧化酶的活性,而低温可使氧化酶类活性始终保持较低水平。各组过氧化物酶的活性无法在长时间储藏下保持高水平的活性且高温对酶活性中心破坏最严重。储藏过程中各组样品的油脂和蛋白质氧化指标水平均呈上升趋势,且高温条件下样品的二者氧化程度最为严重,其中油脂的一次、二次氧化产物含量均增加,表现为过氧化值和酸价的上升,影响蛋白质氧化的活性醛类化合物如MDA含量也同时上升,蛋白质由于结构破坏和氨基酸侧链氧化等多种原因也发生氧化。综上,较高的储藏温度下可以通过影响酶类的活性提高花生油脂和蛋白质氧化速率,导致花生油脂和蛋白质氧化程度加深,由此影响花生的品质。
