芳樟醇、桉叶油素取食胁迫下香樟齿喙象(鞘翅目:象甲科)幼虫的生长发育及抗性基因的转录表达*
2023-08-09王璟廷李寿银徐文轩郝德君
王璟廷 李寿银 左 壮 徐文轩 郝德君
(南京林业大学南方现代林业协同创新中心 南京林业大学林学院 南京 210037)
植物与昆虫协同进化过程中,植物通过合成复杂多样的次级代谢产物以抵御植食性昆虫的取食(陈澄宇等, 2015; Yactayo-Changet al., 2020)。萜类化合物是一种挥发性次生物质,广泛存在于各种植物体内(Mithoferet al., 2012)。作为一种碳基化合物,该类化合物的结构组成以5 个碳的异戊二烯为基本单元,分为半萜、单萜、倍半萜、多萜以及系列萜类衍生物(Mithoferet al., 2012)。已有研究表明,萜类化合物对植食性昆虫具有拒食、产卵趋避、抑制生长发育、毒杀以及引诱天敌等直接或间接防御作用(Seyboldet al., 2006; Loivamakiet al., 2008)。香樟是我国南方重要的常绿树种,广泛用于园林绿化和涵养水源,树体富含大量芳樟醇、桉叶油素等单萜类含氧衍生物(张峰等, 2017;Liet al., 2022),基于这些萜类化合物形成一套特异性的化学防御系统,抑制昆虫的存活、生长发育以及繁殖。例如,芳樟醇和桉叶油素对长红锥蝽(Rhodnius prolixus)具有较强的熏蒸毒性和趋避作用(Sfaraet al., 2009);桉叶油素对苹果黄蚜(Aphis pomi)具有毒杀作用(Wei et al., 2019);合成芳樟醇的转基因烟草(Nicotiana tabacum)可以抑制棉铃虫(Helicoverpa armigera)的产卵(Mccallumet al., 2011)。香樟形成了基于萜类化合物的化学防御系统,但是某些昆虫已经进化出对香樟萜类防御物质的适应机制,实现以香樟作为寄主植物。然而,目前并未建立起某种专性为害香樟的昆虫与其寄主香樟间互作关系的深入研究。
昆虫为克服寄主植物次生物质的防御,也进化出相应的适应机制。其中,昆虫的解毒代谢机制已经开展广泛研究(陈澄宇等, 2015; Heidel-Fischeret al.,2015; Despréset al.,2007)。植物次生化学物质在昆虫体内的代谢可以分为3 个阶段(Sabinaet al., 2015):第1 阶段,在细胞色素P450 酶系(cytochrome P450,CYP450)和羧酸酯酶(carboxylesterases,COE)等解毒酶作用下,催化羟基、羧基等亲核官能团加入到外源化合物中,使其极性增加,易溶于水;第2 阶段,在UDP-葡萄糖醛酸转移酶(UDP- glycosyltransferases,UGTs)和谷胱甘肽S-转移酶(Glutathione S-transferase,GST)等转移酶作用下,将第1 阶段的产物与糖分子和谷胱甘肽等内源性分子轭合,使之易溶于水;第3 阶段,在ATP 转运蛋白(ATP binding cassette transporter,ABC)作用下,将轭合物运出细胞膜,最终排出体外。通常,参与外源物质代谢的解毒酶具有可诱导性。大量研究显示,不同外源物质胁迫能够诱导昆虫相关解毒酶基因的上调表达。例如,苹果蠹蛾(Cydia pomonella)幼虫取食含有香豆素和槲皮素的人工饲料后,中肠内CYP332A19和CYP337B19显著上调表达(陈高满等, 2020);COE2、CYP6-like 5和BtGST2的上调表达参与烟粉虱(Bemisia tabaci)对咖啡因、尼古丁、槲皮素等次生物质的代谢适应(Halonet al., 2015);华山松大小蠹(Dendroctonus armandi)在华山松(Pinus armandii)萜类熏蒸胁迫下CYP6基因家族表现出明显的诱导响应模式(Daiet al., 2015)。
表皮蛋白(cuticular protein, CP)是昆虫体壁的主要成分,在昆虫表皮形成和发育中具有重要作用。昆虫通过大量合成表皮蛋白,与几丁质形成复合体,沉积于原表皮层,使得表皮厚度增加,减少外源化合物的渗透,缓解了昆虫体内解毒代谢压力,降低了外源化学物质进入靶标位点的概率,形成表皮抗性(cuticular resistance)(梁欣等, 2014; 孙雅雯等, 2015;Balabanidouet al., 2018)。昆虫表皮抗性作为杀虫剂抗药性机制之一,已在卫生害虫研究中被证实。例如,溴氰菊酯抗性品系的淡色库蚊(Culex pipiens)共有14个表皮蛋白基因上调(Fanget al., 2015; Panet al.,2009),类似的结果在冈比亚按蚊(Anopheles gambiae)中也有报道。除虫菊酯和DDT 抗性品系按蚊的CPLCG3、CPR124、CPR127、CPR129基因表达量远高于敏感品系(Awololaet al., 2009)。然而,有关昆虫表皮蛋白在抵御植物次生物质的功能研究尚未见报道。但有研究表明,昆虫在寄主转换过程中大量表皮蛋白基因出现差异表达,推测表皮蛋白参与昆虫对不同寄主植物化学物质的适应(Birnbaumet al., 2020)。
1 材料与方法
1.1 试验材料
1.1.1 供试植物 香樟的1~3 年生新鲜枝条采自南京林业大学校园,带回实验室后去除叶片,保留枝干部分,清洗干净后晾干。利用高速万能粉碎机FW100(天津市泰斯特仪器有限公司)粉碎后过80 目筛,将粉末置于4 ℃下储存,备用。
1.1.2 供试昆虫 香樟齿喙象幼虫采自上海市松江区泖港镇香樟人工纯林(30°56′6.15″ N ,121°12′32.76″E),带回实验室后参照Li 等(2021)的方法进行人工饲养,饲养条件:温度28 ℃±1 ℃,相对湿度70%±5%,光周期为16∶8(L∶D)。选择健康的羽化成虫置于养虫盒内配对,收集卵粒,待卵孵化后,选择生理状态一致的初孵幼虫用于后续试验。
1.1.3 试剂和试剂盒 芳樟醇(CAS: 78-70-6,纯度95%)和桉叶油素(CAS: 470-82-6,纯度99%),购自南京斑马实验器材有限公司。HiScript 1st Strand cDNA Synthesis Kit 反转录试剂盒和SYBR Premix Ex Taq II试剂盒,购自南京诺唯赞生物科技股份有限公司。
1.2 毒力测定
采用人工饲料混药法(Pintoet al., 2003)进行毒力测定。分别配制不同质量分数的芳樟醇(0、2、4、6、10、12、30、50、70、100、150 以及200 mg·g-1)和桉叶油素(0、1、2、3、5、7、20、40、50、70、90 以及100 mg·g-1)的人工饲料。利用不同质量分数处理的饲料喂食香樟齿喙象初孵幼虫72 h,记录死亡个数(用毛刷拨动虫体,无明显生命迹象的幼虫视为死亡个体)。每个质量分数处理组设置不少于60 头幼虫。
1.3 芳樟醇和桉叶油素长期胁迫下香樟齿喙象幼虫生长发育指标测定
根据预试验结果,设定2 种质量分数处理(芳樟醇:10.0 mg·g-1和19.7 mg·g-1;桉叶油素:5.0 mg·g-1和15.2 mg·g-1),随机选择200 头初孵幼虫进行取食胁迫饲养,直至化蛹。以未添加2 种萜类的饲料喂养的幼虫作为对照。定期记录不同处理幼虫的蜕皮次数、体重以及死亡情况。根据幼虫各龄期的存活率(LV)、发育历期(DT)和体重(BW)指标,计算相对表现指数(relative performance index, RPI)(Sotoet al., 2014)。公式如下:
在侦查决策中,决策方案随着案件条件不同而各异。在某一起案件侦查中发挥重要作用的决策方案不一定能够应用于其他同类案件。针对未发案件所作的应急决策方案虽然可以参考、借鉴以往侦破的成功方案,但是针对已发案件的决策方案则不能简单的照搬、刻板的模仿,侦查决策方案的风险因素更加复杂。因而如何评价侦查决策方案,尽可能地将侦查决策中的风险因素降低,以更好地实现侦查目的就成为需要研究的内容。笔者认为在评价侦查决策时,应主要衡量侦查成本、侦查效益两个因素。
1.4 基于qRT-PCR 的基因表达量测定
1.4.1 RNA 提取和第一链cDNA 合成 参照1.3 中处理方法,分别选取LC15浓度下芳樟醇和桉叶油素饲养至4 龄的香樟齿喙象幼虫,以及对照组幼虫各15 头。每个处理组设置3 个生物学重复,每个重复5 头幼虫。液氮速冻后用于总RNA 提取。利用TRIzol 提取法,提取各个样品总RNA,并检测RNA 质量和浓度。RNA 完整性通过1%琼脂糖凝胶电泳进行检验。使用HiScript 1st Strand cDNA Synthesis Kit 反转录试剂盒合成第一链cDNA。
1.4.2 实时荧光定量PCR 根据前期香樟齿喙象转录组测序数据(Chenet al., 2020),参照黑腹果蝇(Drosophila melanogaster)、 赤拟谷盗(Tribolium castaneum)解毒酶基因(Matthewset al.,2015; Richardset al., 2008)进行本地blast 比对, 分别挑选5 条CYP450、COE、UGT、GST、ABC 以及表皮蛋白基因的核苷酸序列,利用Primer Premier 5.0 设计实时荧光定量PCR 特异性引物(附表1)并通过blastx 比对到NCBI nr 库分析基因同源性(附表2)。使用SYBR Premix Ex Taq II 试剂盒在Applied Biosystem 7500 实时荧光定量PCR 系统进行PCR 反应。反应体系:5 µL Hieff qPCR SYBR Green Master Mix (2×),上下游引物(10µmol·L-1)各0.4 µL,2 µL cDNA 模板,7.2 µL ddH2O。每个反应体系设置3 个技术重复。PCR 反应条件:95 ℃ 5 min,95 ℃ 10 s,60 ℃ 40 s,循环数为40。
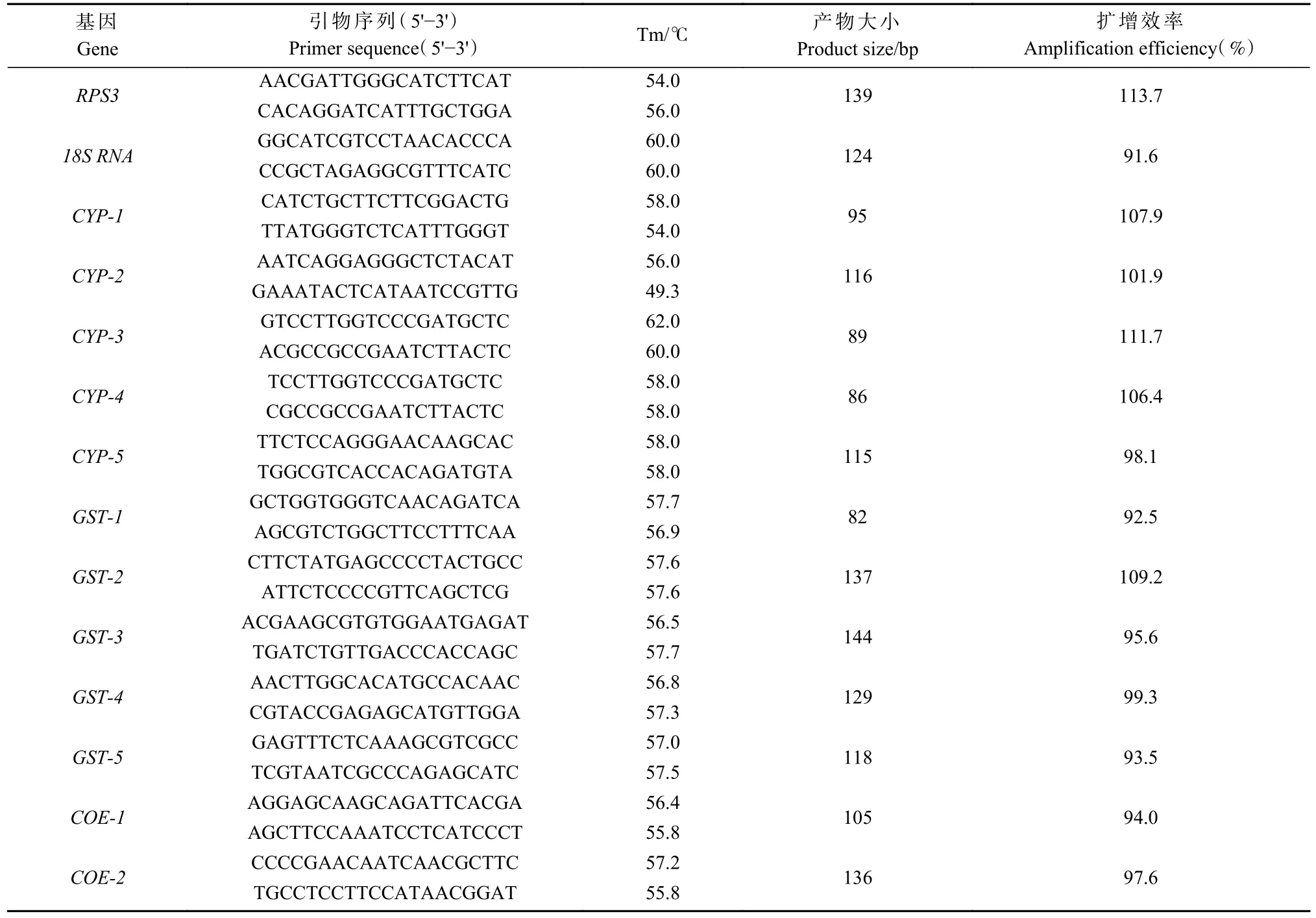
附表1 所选香樟齿喙象有关解毒酶基因和表皮蛋白基因转录组基因序列引物Appendix 1 The selected primers for detoxification gene and cuticle protein gene in Pagiophloeus tsushimanus
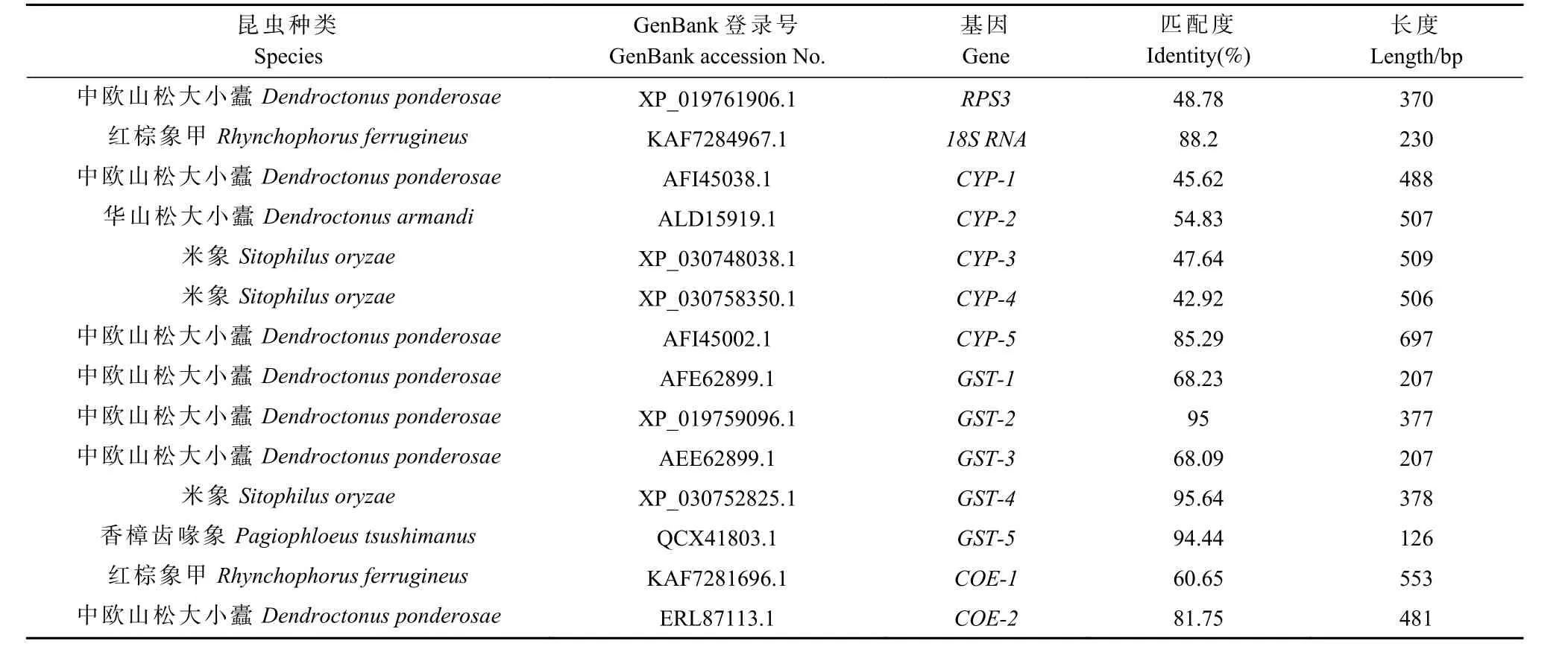
附表2 香樟齿喙象有关解毒酶基因和表皮蛋白基因转录组基因序列同源性分析Appendix 2 Homology analysis of gene sequence related to detoxification enzyme gene and epidermal protein gene transcriptome of Pagiophloeus tsushimanus
1.5 数据处理与分析
利用二次逻辑回归函数对毒力测定结果进行拟合,计算半数致死浓度(LC50)和亚致死浓度(LC15),回归函数表达式如下:
式中,y为香樟齿喙象幼虫死亡率(%),x为药剂质量分数(mg·g-1),a1、a2、p、x0为参数。通过卡方列检验(P<0.05)确定最大非致死浓度(LC00)。使用Kruskal-Wallis ANOVA 检验分析不同阶段的发育历期、幼虫和蛹的重量(P<0.05),进行成对检验以评估处理之间的差异显著性。使用Kaplan- Meier 生存分析法绘制不同次生物质、不同质量分数处理下香樟齿喙象幼虫的存活曲线,通过对数秩检验确定各个处理间是否存在显著差异性。以RPS3 和18sRNA 作为双内参基因(Chenet al., 2020),采用2-ΔΔCt法计算基因相对表达量。利用单因素方差分析(ANOVA)以及t检验,分析处理组与对照组之间基因表达量的差异显著性。所有数据处理及绘图工作均在软件Origin 2021b 中完成。
2 结果与分析
2.1 芳樟醇和桉叶油素的毒力分析
采用人工饲料混药法对香樟齿喙象初孵幼虫的毒力结果见图1。可知,香樟齿喙象幼虫在取食添加芳樟醇和桉叶油素的人工饲料3 天后,2 种萜类的致死中浓度LC50分别为47.2 、41.7 mg·g-1,说明香樟齿喙象对于芳樟醇比桉叶油素显现出更高的耐受性。通过卡方列检验,芳樟醇和桉叶油素的最大非致死浓度分别为10.0 、5.0 mg·g-1,亚致死浓度LC15浓度分别为19.7、15.2 mg·g-1。
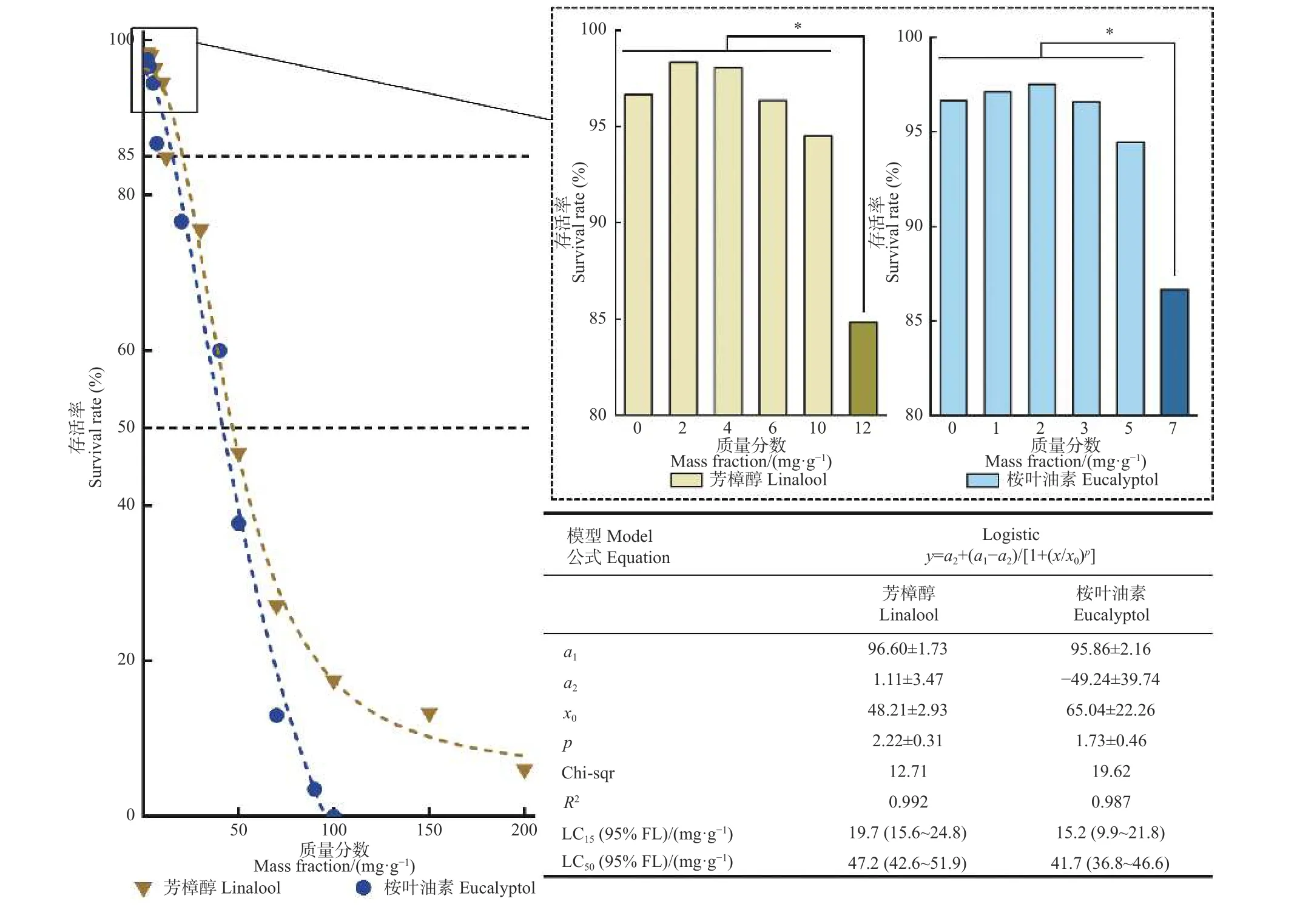
图1 芳樟醇和桉叶油素对香樟齿喙象初孵幼虫的室内毒力测定结果Fig. 1 Toxicity of linalool and eucalyptol to newly hatched larvae of P. tsushimanus
2.2 2 种质量分数芳樟醇和桉叶油素对香樟齿喙象幼虫存活率和发育历期影响
由图2 可以看出,取食含有不同质量分数的芳樟醇和桉叶油素人工饲料的香樟齿喙象幼虫生长龄期存在差异。结果表明,2 种萜类的胁迫均引起香樟齿喙象幼虫的发育历期延长。在芳樟醇和桉叶油素LC00剂量下,香樟齿喙象一龄幼虫龄期无显著影响,从2 龄开始直至化蛹,发育历期延长。在芳樟醇和桉叶油素LC15剂量下,香樟齿喙象各龄龄期均有所延长。在5 龄和预蛹期,2 种萜类物质对香樟齿喙象幼虫影响最大,龄期增加10~20 天。在桉叶油素和芳樟醇胁迫下,香樟齿喙象幼虫的发育历期从长到短的依次为芳樟醇19.7 mg·g-1(120.41 天)>桉叶油素15.2 mg·g-1(114.79 天)>芳樟醇10.0 mg·g-1(109.78 天)>桉叶油素5.0 mg·g-1(99.24 天)>对照组CK(88.66 天)。
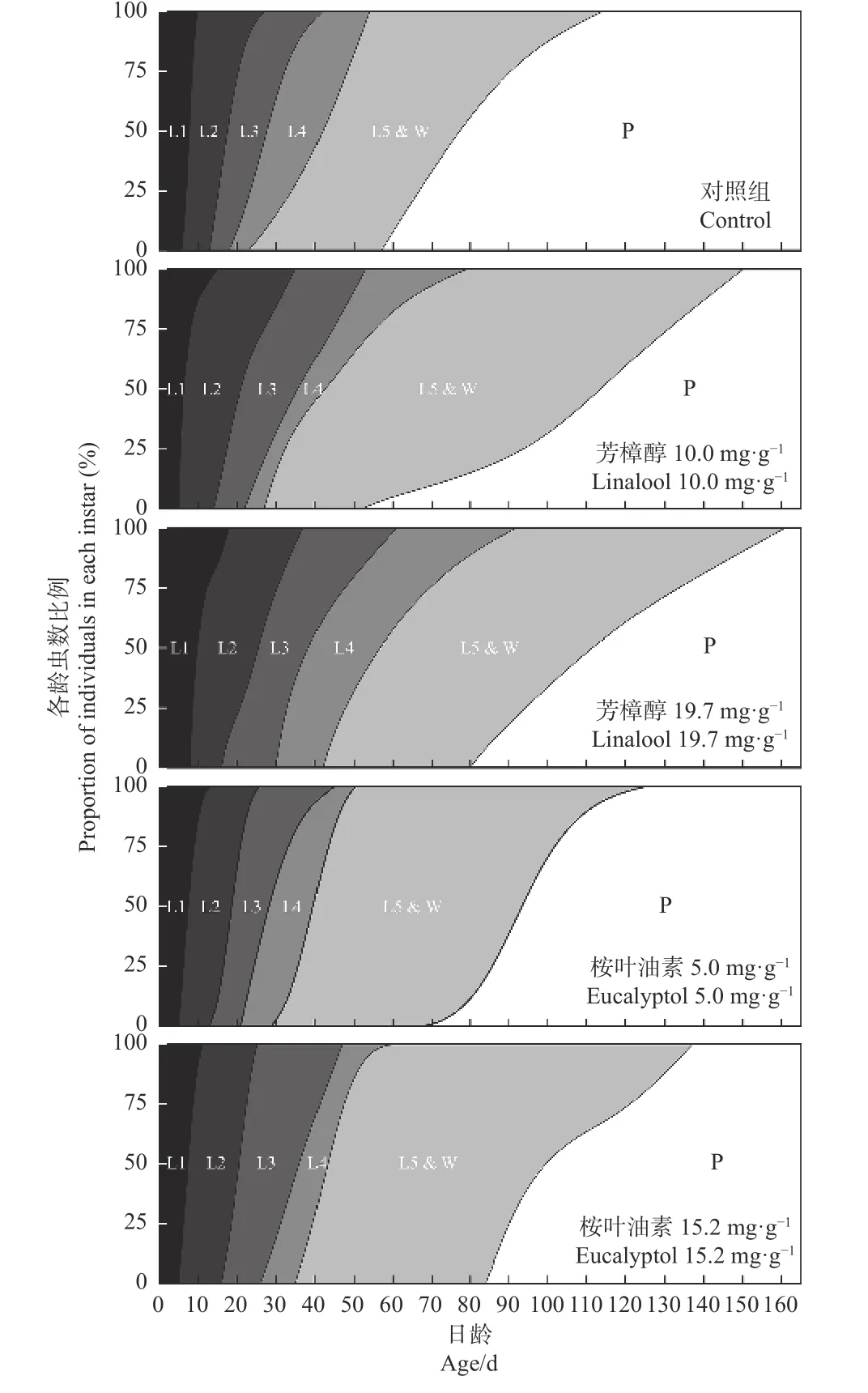
图2 2 种质量分数芳樟醇和桉叶油素对香樟齿喙象幼虫发育历期的影响Fig. 2 The effects of two mass fraction of linalool and eucalyptol on the development duration of P. tsushimanus larvae
取食含有不同质量分数芳樟醇和桉叶油素的人工饲料对香樟齿喙象不同龄期幼虫体重的影响结果见表1。可以看出,在芳樟醇和桉叶油素胁迫下,香樟齿喙象幼虫各个发育阶段的取食和消化能力均受到影响,处理组幼虫的体重显著低于对照组(P<0.05)。取食19.7、10.0 mg·g-1处理的芳樟醇和15.2 mg·g-1处理的桉叶油素饲料的蛹重明显小于对照组(P<0.05),而桉叶油素5.0 mg·g-1胁迫下,处理组和对照组的蛹重无明显差异(P>0.05)。
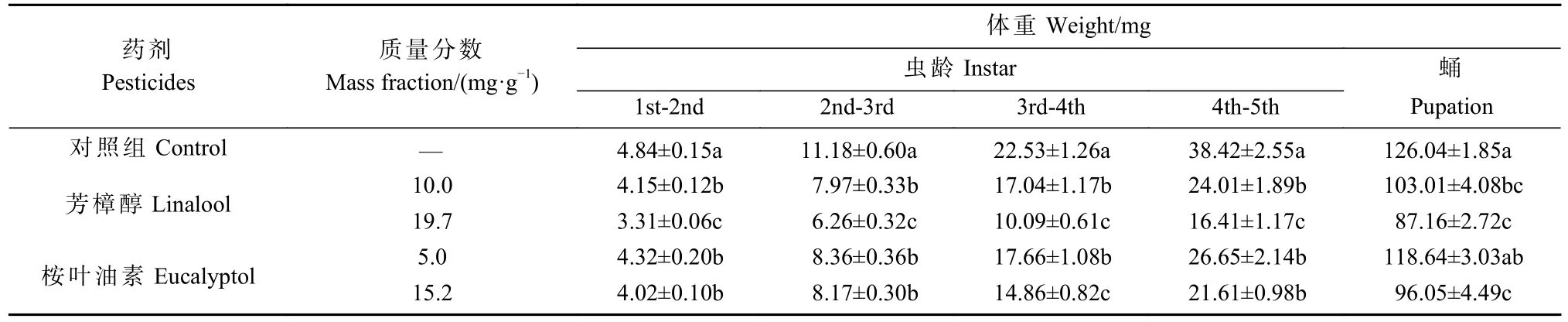
表1 2 种质量分数芳樟醇和桉叶油素胁迫下对香樟齿喙象幼虫体重和蛹重的影响①Tab. 1 The effects of two mass fraction of linalool and eucalyptol on the larval and pupa weight of P. tsushimanus
利用Kaplan-Meier 生存分析法绘制香樟齿喙象幼虫在不同质量分数的芳樟醇和桉叶油素的存活曲线,并进行卡方检验。结果如图3 所示,可知含有不同质量分数的芳樟醇和桉叶油素的半人工饲料饲喂幼虫的存活曲线存在明显差异(χ2=300.679,P<0.001)。通过比较死亡时间,发现香樟齿喙象幼虫死亡集中于初孵和蜕皮时,即0~3、6~12、18~21、30~33 天几个时间节点,由于试虫在这些节点刚完成蜕皮进入下一龄期,相对较脆弱,故死亡率较高。观察发现,幼虫表现为头壳发白,体壁僵硬,丧失进食能力。
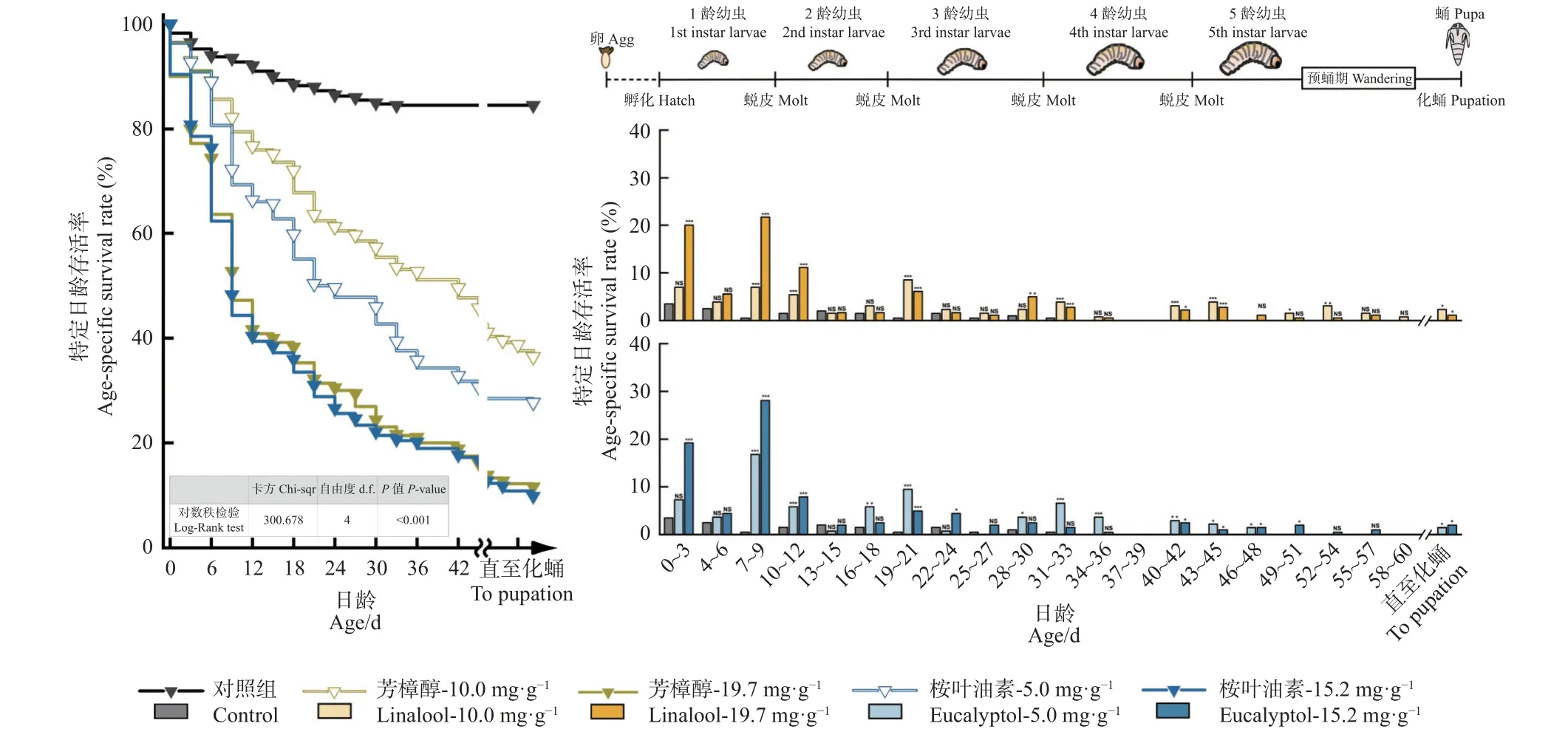
图3 2 种质量分数芳樟醇和桉叶油素胁迫下对香樟齿喙象存活率的影响Fig. 3 The effects of two mass fraction of linalool and eucalyptol on the survival rate of P. tsushimanus
结合体重、龄期、存活率等指标,将其组合成相关表现指数(RPI),作为评价香樟齿喙象对2 种次生化学物质的活性指标。由表2 可知,取食不同质量分数芳樟醇和桉叶油素饲料的香樟齿喙象幼虫的相关表现指数均小于对照组,当质量分数提高,差距则增大。以上结果表明,不同质量分数芳樟醇和桉叶油素均可抑制香樟齿喙象幼虫的生长,且抑制作用随着质量分数的增加而增强。
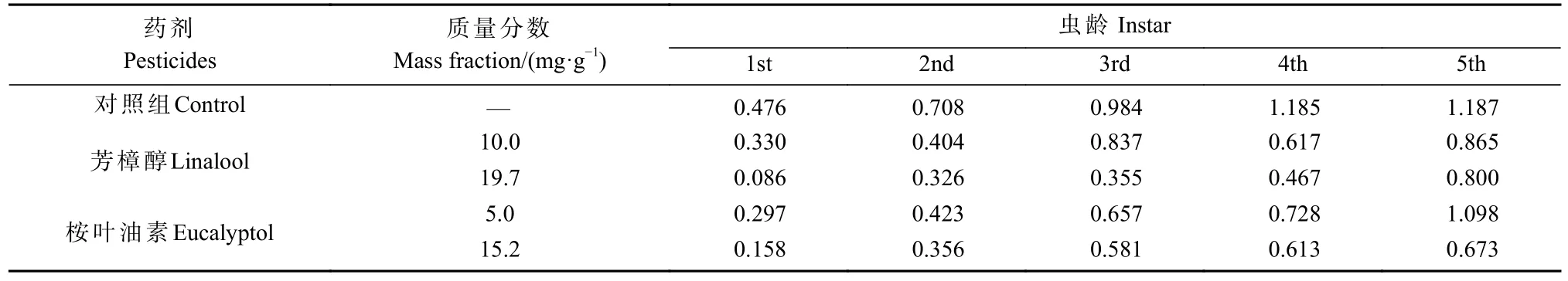
表2 2 种质量分数芳樟醇和桉叶油素引起的香樟齿喙象幼虫相关表现指数Tab. 2 The relative performance index of linalool and eucalyptol under the stress of two mass fraction in P. tsushimanus
2.3 芳樟醇和桉叶油素对香樟齿喙象幼虫解毒酶基因和表皮蛋白酶基因表达的影响
在芳樟醇和桉叶油素两种萜类的胁迫下,香樟齿喙象4 龄幼虫解毒酶基因和表皮蛋白基因表达特性见图4。在2 种质量分数芳樟醇的胁迫下,细胞色素P450 基因家族中CYP1、CYP3、CYP4 基因表达和表皮蛋白基因家族中CP1、CP3、CP4、CP5 均显著上调(P<0.05),GST1、GST2、GST5 的基因则明显下调表达(P<0.05),其他解毒酶基因表达和对照组无明显差别(P>0.05)。在2 种质量分数桉叶油素胁迫下,UGT1基因显著上调表达(P<0.05),COE1 基因则明显下调表达(P<0.05),其他解毒酶基因和表皮蛋白基因与对照组相比无显著差异(P>0.05)。
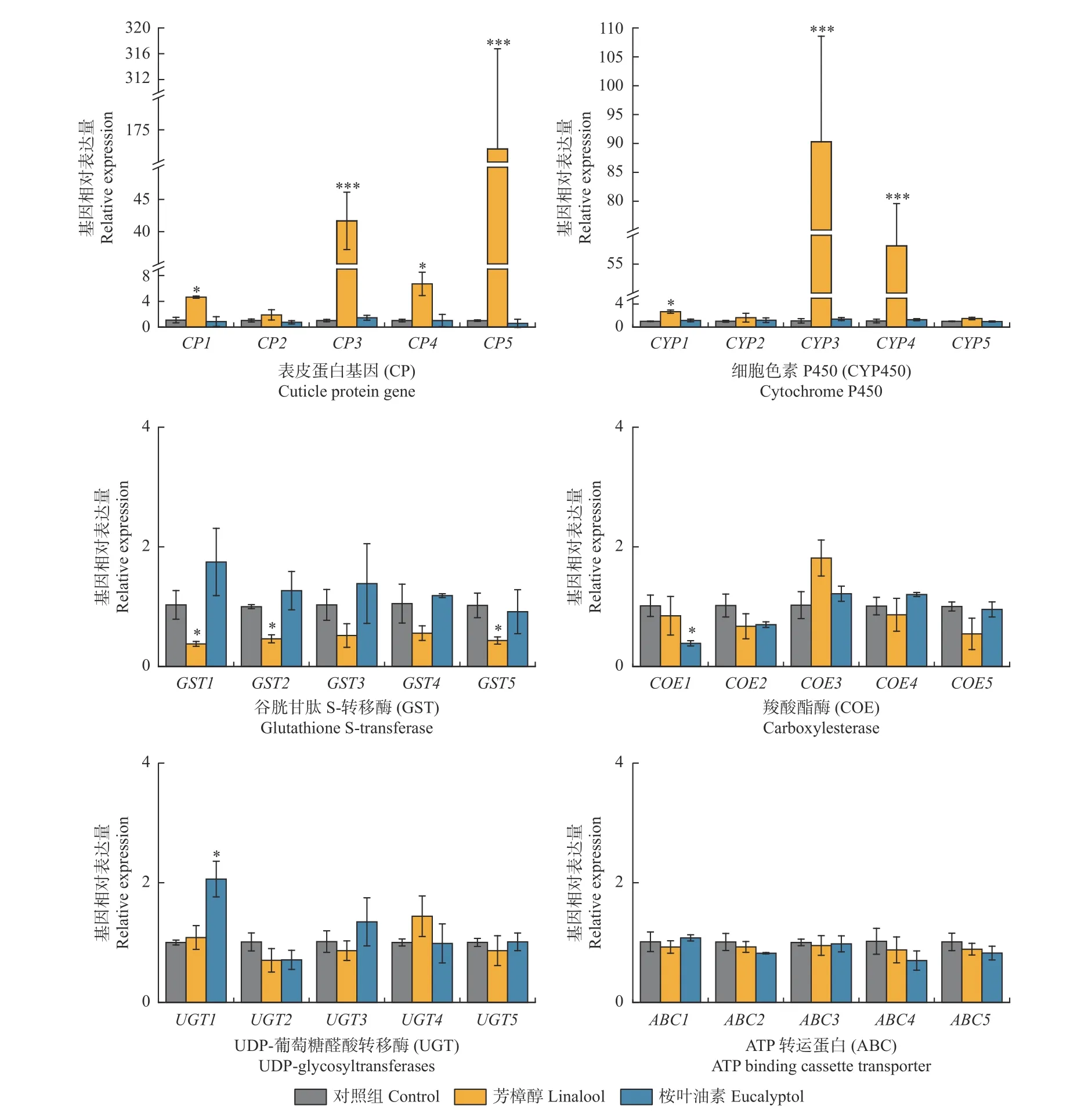
图4 芳樟醇、桉叶油素胁迫下香樟齿喙象4 龄幼虫解毒酶和表皮蛋白基因表达情况Fig. 4 Relative expression levels of detoxification genes and cuticule proteins in P. tsushimanus fourth larvae with linalool and eucalyptol
3 讨论
植物为应对植食性昆虫侵害进化出多重的防御机制(谢辉等, 2012)。其中,复杂多样的次生代谢物质构成了植物的化学防御系统(Nishida, 2014; 刘兴平等, 2003)。已有研究表明,萜类次生物质广泛存在于植物体内,对昆虫具有毒杀、趋避等生物活性(Huanget al., 2017)。本研究选取芳樟醇和桉叶油素2 种香樟树体内主要萜类,测定其对专性为害香樟的香樟齿喙象的生物活性。结果表明,在芳樟醇和桉叶油素短时胁迫下,初孵幼虫表现出不同程度上的耐受性,即半致死浓度(LC50)分别为19.7 和15.2 mg·g-1。然而,长期取食含有芳樟醇和桉叶油素的人工饲料,香樟齿喙象幼虫的生长发育受到不同程度的抑制作用,并且这种抑制作用随2 种萜类浓度增加而增强,可能是由于植物次生物质对昆虫的影响仅在一定浓度下起作用(陈澄宇等. 2015; Mauryaet al., 2020)。
尽管萜类化合物一定程度上为香樟提供了防御作用,但仍有超过20%的香樟齿喙象通过延长发育历期,降低蛹重等方式在低剂量胁迫下完成了其幼虫阶段的生长发育,进入蛹期。值得注意的是,萜类胁迫下香樟齿喙象幼虫集中死亡发生于蜕皮期前后,表现为无法完成蜕皮过程或鞣化不完全,可能是由于旧表皮褪去,而新表皮刚刚形成,该阶段的下表皮对外源萜类化合物的阻隔能力较弱,导致短时间内大量萜类进入虫体而引起死亡。这一现象在类似研究中也被报道,如甜菜夜蛾(Spodoptera exigua)在有机磷杀虫剂处理后,蜕皮后气管边出现裂缝、角质层畸形等现象,导致其在蜕皮期大量死亡(Adamskiet al., 2009)。淡色库蚊幼虫在新烟碱类杀虫剂处理后,蜕皮过程中其头壳无法完全脱去,引起死亡(Songet al., 2016)。基于这一现象,未来可以香樟齿喙象表皮合成相关基因作为潜在的分子靶标,为开发新型防治技术提供理论依据。
为进一步明确香樟齿喙象幼虫体内可能参与响应芳樟醇和桉叶油素胁迫的关键功能基因,分别测定了几种解毒酶基因和表皮蛋白基因的转录表达水平。结果发现,芳樟醇胁迫能够同时诱导香樟齿喙象幼虫体内细胞色素P450 酶和表皮蛋白的上调表达。昆虫细胞色素P450 酶可通过羟基化、环氧化等方式代谢萜类物质,例如,中欧山松大小蠹(Dendroctonus ponderosae)进化出完善的细胞色素P450 酶解毒代谢体系以代谢寄主植物的次生代谢物质,其中CYP6DJ1基因将松油烯和柠檬烯氧化为乙醇和环氧化物,CYP6DE1 基因将α-派烯、β-派烯氧化为醇类物质(Blomquistet al., 2021; Chiuet al., 2019a; Chiuet al.,2019b)。本研究中,芳樟醇诱导香樟齿喙象幼虫的CYP1、CYP3、CYP4 3 条基因上调表达,暗示CYP450参与芳樟醇的代谢分解,后续将深入研究芳樟醇在香樟齿喙象体内的代谢途径,以验证CYP450 酶的代谢功能。表皮蛋白作为昆虫体内重要的结构蛋白,大量富集于上表皮和外表皮,构成昆虫抵御外源化学物质的物理屏障(Balabanidouet al., 2018; Moussian, 2010)。研究发现,昆虫表皮蛋白基因数量超过该昆虫基因组中蛋白质编码基因总数的1%,在昆虫生长发育、繁殖、环境适应中发挥重要作用(梁欣等, 2014; Charles,2010)。在冈比亚按蚊、淡色库蚊等昆虫中发现,化学农药胁迫下会诱导表皮蛋白基因的表达,导致表皮层增厚,增强对化学农药的阻隔,提高自身抗药性(Yahouedoet al., 2017; Fanget al., 2015; Lillyet al.,2016)。但是,表皮蛋白基因介导昆虫抵御植物次生物质的作用报道较少。已有研究发现,植食性昆虫在转主胁迫后由表皮蛋白基因构成的表皮结构成分具有显著差异(Birnbaumet al., 2020),表明植食性昆虫在转移到新寄主后可能利用表皮蛋白应对其中的植物次生物质。本研究发现,芳樟醇诱导香樟齿喙象幼虫CP1、CP3、CP4、CP5 4 条CP 基因上调表达,表明香樟齿喙象幼虫可能通过合成大量表皮蛋白,引起表皮增厚,进而减缓芳樟醇的渗透作用。因此,尚需探究芳樟醇胁迫后幼虫表皮厚度的变化以及表皮蛋白的富集定位,明确香樟齿喙象对芳樟醇的解毒代谢机制。
此外,本研究发现桉叶油素取食胁迫并未诱导解毒酶基因和表皮蛋白基因的明显表达,仅有1 条UDP-葡萄糖醛酸转移酶基因的表达量显著上调。可能是由于不同外源化合物种类、处理质量分数均会对相关基因的表达响应产生不同影响。例如,咖啡因和苯巴比妥能够诱导黑腹果蝇细胞色素P450 酶和谷胱甘肽-S-转移酶基因的上调表达,但二嗪农、氯吡脲等杀虫剂则无诱导作用(Willoughbyet al., 2006)。类似的结果在甜菜夜蛾中也有报道,高效氯氟氯氰菊酯、氯虫腈、甲氟腙和茚虫威可以诱导甜菜夜蛾体内细胞色素P450 酶和UDP-葡萄糖醛酸转移酶基因的上调表达,但阿维菌素则表现出明显的抑制作用(Huet al.,2019)。毒力测定结果表明香樟齿喙象幼虫对桉叶油素更敏感,说明桉叶油素抑制了香樟齿喙象体内相关解毒酶系的活性,有助于提高寄主植物对香樟齿喙象的防御能力。
4 结论
香樟齿喙象幼虫对芳樟醇和桉叶油素具有一定的耐受能力,但是在长期胁迫下其生长发育则受到不利影响。其中,芳樟醇胁迫同时诱导幼虫体内细胞色素P450 酶和表皮蛋白基因的上调表达,而桉叶油素胁迫仅诱导幼虫体内一条UGT 解毒酶基因的上调表达。香樟齿喙象幼虫能够调控自身解毒代谢机制和表皮阻隔作用,以适应香樟的化学防御。
