水肥胁迫对基于时间序列图像的苗木表型信息的影响*
2023-08-09杨琨琪张慧春边黎明
杨琨琪 张慧春 张 萌 边黎明
(1. 南京林业大学机械电子工程学院 南京 210037;2. 南京林业大学林学院 南方现代林业协同创新中心林木遗传与生物技术教育部重点实验室 南京 210037)
植物生长发育过程受基因和环境共同影响,逐渐形成对逆境的适应和抵抗能力,逆境环境下植物表型体现出复杂性,搭建植物表型信息采集平台对植物生长过程进行连续动态监测,可研究植物在时间跨度上的表型结构变化,有助于了解植物生长发育情况和不同环境变化下植物的响应状况(Johannsenet al.,2014;张晓东等,2019;王传宇等,2018)。可见光相机能够实现低成本、近距离、快速的图像捕获以测量植物表型参数(Underhillet al.,2020;李玉荣等,2020;李秋洁等,2021),结构简单且适应性强(杨有新等,2015;Crainet al.,2016),应用极为广泛。基于图像的植物表型信息监测,图像获取时间可跨越整个生长季,然而现有研究多关注某个特定时刻的植物图像采集,面向苗木表型信息提取的自动化时间序列图像采集和处理方法鲜见报道。
水分和养分是植物正常生长的重要因子,其在植物生长发育过程中起着重要作用,合理的水肥配置能够显著提高植物生产力,尤其在水分和养分受到限制时,科学、精准地确定水分和养分用量具有重要意义(陈隆升等,2021;倪铭,2021;杨会肖等,2021),研究随时间变化植物在干旱胁迫与梯度养分下的适应机制和能力,将为植物栽培与管理提供理论依据和技术支持。
簸箕柳(Salix suchowensis)为杨柳科(Salicaceae)柳属(Salix)植物,基因组大小356 Mb,具有易繁殖、速生丰产和轮伐期短等特点,是重要的生物质能源和造林用材树种(石吉勇等,2019)。本研究以簸箕柳为试验样本,构建多视角植物表型信息采集平台,通过分析一定时间跨度上的簸箕柳序列图像,获取其在水肥胁迫处理下的表型参数并进行分析,以期为苗木表型信息的自动提取、植物逆境响应机制的精准解析提供科学依据。
1 材料与方法
1.1 供试材料与试验设计
选用高度12±0.3 cm、直径4±0.2 mm、生长健壮且长势一致的48 株簸箕柳枝条进行扦插式种植,统一放入内径10 cm、深11.5 cm 的圆柱形花盆中,每盆1 株,扦插基质中泥炭、珍珠岩比例为3∶1。苗木置于人工气候箱(宁波江南制造厂,RXZ型)内培养,气候箱环境参数设置如下:湿度65%、温度28 ℃、光照强度278 µmol·m-2s-1、昼夜时间比16 h∶8 h。
设置氮肥和水分2 个因素,采用完全随机设计。氮肥因素设置T1(尿素每盆0.1 g)、T2(尿素每盆0.1 g,扦插种植第40 天再施加尿素每盆0.1 g)、T3(尿素每盆0.2 g)、T4(尿素每盆0.2 g,扦插种植第40 天再施加尿素每盆0.2 g)、T5(尿素每盆0.4 g,扦插种植第40 天再施加尿素每盆0.4 g)5 个水平,同时设置1 个CK 处理组(尿素每盆0 g)作为对照,每组4 株;水分因素设置正常水分(normal water supply,W)和干旱胁迫(drought stress,D)2 个水平,干旱处理前,保证基质含水量相同。对正常水分处理组,扦插后第8~20 天每株浇水量为每3 日20 g,第20~30 天每株浇水量为每2 日30 g,第30~40 天每株浇水量为每2 日60 g,第40~50 天每株浇水量为每2 日80 g;对旱胁迫处理组,扦插后第8~20 天每株浇水量为每3 日20 g,第20~30天每株浇水量为每2 日30 g,第30~50 天每株浇水量为每2 日30 g。从扦插种植第26 天开始图像自动采集,手动测量表型参数,以便对照检验,每2 日1 次,共采集、测量10 次,扦插种植第50 天结束试验。
1.2 植物表型信息采集
单一视角采集植物图像信息存在枝叶遮挡无法获得植物完整形态结构的情况,多视角采集则可弥补该缺陷(Luet al.,2020;刘阳等,2020)。本研究多视角植物表型信息采集平台(图1)主要包括待测植物簸箕柳、CCD(charge coupled device,电荷耦合器件)相机(型号GO-5000C-PGE,分辨率2 560 像素×2 048 像素、LM8HC-SW 定焦镜头)、相机升降模块、旋转模块、稳定光环境暗箱。升降模块利用丝杆控制相机上下移动,旋转模块利用蜗轮蜗杆结构精准控制装置圆周旋转,以植物固定不动、相机围绕旋转的方式稳定拍摄,对每株植物以18°为旋转角度间隔采集图像,1 个圆周采集20 张图像;稳定光环境暗箱主要包括上方四边缘排布的4 W 白光色LED 光源和用黑色植绒布全覆盖的1.2 m×1.2 m×1.5 m 型材框架,以提供稳定的成像光环境。

图1 多视角植物表型信息采集平台Fig. 1 Multi-perspective plant phenotype information collection platform
1.3 图像拼接
植株与相机距离保持固定不变,当植株随时间推移自身不断长高时,会超出相机拍摄范围;如果增加相机与植株距离,会导致采集平台所占空间不断变大。为解决上述问题,本研究利用LabVIEW 软件进行相机高度判断,实时采集图像并判断整株植物是否在相机拍摄范围内,当整株图像不在拍摄范围内时,采用调整相机高度方式分段采集植株图像,之后拼接2 张局部图像得到完整得植物表型信息(图2)。

图2 平台调整相机高度拼接完整图像示意Fig. 2 Platform adjustment camera height mosaic complete image schematic
1.3.1 直接拼接法 直接拼接法预先设置重复部分高度,将高角度图像去除重复部分与低角度图像直接拼接,图3a 为直接拼接法效果,由图中虚线框可见图像拼接处有一定位移偏差,这是因为平台轻微抖动以及相机去畸变算法的局限性,2 张图像存在一定细节不能吻合,位移偏差会造成植物表型参数提取不准确等问题。

图3 3 种拼接算法效果Fig. 3 Three kinds of splicing algorithm effect
1.3.2 暴力拼接法 暴力拼接法是对2 张图像采用ORB(oriented fast and rotated brief,面向角点的快速检测)算法,为便于处理,该算法将植物高角度图像与低角度图像从RGB(red green blue,红绿蓝)色彩空间转换至GRAY(raw gray bytes,原始灰色字节)色彩空间得到植物灰度图像。
色彩空间转换的计算方法如下:
式中:Gray 为灰度值;R、G、B为植物图像在RGB 色彩空间下三通道的分量值,无量纲单位。
转换后的灰度图像先采用FAST(features from accelerated segment test,快速特征点检测)算法进行特征点检测,再采用BRIEF(binary robust independent elementary features,二进制独立特征点处理)算法将这些关键点转换为特征向量。BRIEF 针对同一关键点重复该流程256 次,256 个像素亮度比较结果输入该关键点的二元特征向量中,以此类推,为图像中每个关键点创建一个向量。根据匹配向量对高角度图像进行裁剪和透视变换,对低角度图像进行拼接。
虽然暴力拼接法计算速度快,但只针对灰度图像进行特征点运算。图3b 为暴力拼接法效果,可以看出因簸箕柳叶片没有明显形状特征和易于检测定位关键点,拼接效果较差。
1.3.3 近似拼接法 为解决直接拼接法和暴力拼接法因位移偏差导致潜在表型提取困难,本研究提出一种近似拼接法。该算法在直接拼接法基础上,对2 张图像待拼接的边缘区域进行形态学处理和像素级对准,使高角度和低角度分别拍摄的上、下2 张图像更为准确地连结成一个整体,以便后期基于图像的苗木表型信息精确提取。
对高、低角度2 张图像进行二值化处理,并设置近似区域为20 像素高度区间,取低角度图像第0 行像素值,得到具有W个元素的数组ArrayM0,从数组ArrayM0非零值开始截取,至最后一个非零值结束,生成一个新的数组ArrayN0;同样方法处理高角度图像底部近似区域内像素,得到20 个元素数不尽相同的数组,将低角度图像生成的20 个数组与高角度图像生成的20 个数组进行对比,判断是否存在有数组相同。若存在一个高角度图像数组与低角度图像数组ArrayNn相同,则设置该数组为重合处,进行拼接并对应数组中第1 个非零值位置;若存在多个高角度图像数组与低角度图像数组N相同的情况,取最小M即满足条件下高角度图像最接近直接拼接的像素行;若不存在多个高角度图像数组与低角度图像数组N相同的情况,则采用直接拼接法对图像进行拼接。
图3c 为近似拼接法效果,可以看出该算法相较于暴力拼接法和直接拼接法具有更好适应性,能够较好消除拼接痕迹,实现图像无缝拼接,避免传统图像拼接法存在的问题,保证数据客观准确性。
2 表型参数提取
形态结构表型参数是表征植物生产力的重要指标(王旭等,2017;张慧春等,2020;翁杨等,2019)。对簸箕柳来说,直径细、树体矮的适于用作编织,直径粗、树体高的适于用作固沙等生态功能,因此通过准确、快速、无损方法测得植物形态结构表型参数(刘守阳等,2020),可有效监测植物长势和状态,进而根据栽培目标调控苗木水肥处理方法以及进行基因型选择等(Bevanet al.,2017)。
2.1 多视角图像下株高提取
采用像素计算法提取多视角苗木图像株高,为便于提取彩色图像中植株信息,需分割彩色图像的植株部分,首先将植物图像从RGB 色彩空间转到HSV(hue saturation value,色调、饱和度、明度)色彩空间,然后设定HSV 三通道下的阈值,将图像中植株部分分割出来。
RGB 色彩空间转换到HSV 色彩空间的计算方法如下:
式中:R、G、B为植物图像在RGB 色彩空间三通道中的值,无量纲单位。
对分割后的簸箕柳图像边缘进行标注并连通,统计图像中所有连通域,计算出最大连通域,去除噪点和土壤等干扰项,得到簸箕柳植株部分图像。绘制簸箕柳图像最小包围矩形,利用已知尺寸(2 cm×2 cm)的红色正方形标定板进行比例缩放,得到植株实际株高(H)。
簸箕柳株高计算方法如下:
式中:H为图像测得的株高实际值,cm;a为图像中红色正方形标定板的边长像素, pixel;h为图像处理得到的株高像素, pixel。
考虑到植物生长时会有一定倾斜角度,使测得株高偏小,本研究选取20 个图像角度下测得株高的平均值记为Have,株高最大值记为Hmax,正面拍摄单张图像提取的株高记为Hsingle,将三者进行对比分析。以统计量决定系数(R2)为评价指标评估3 种算法提取的株高准确率,以测试株高提取算法的性能。R2表示由算法模型解释的测量方差百分比,取值范围[0,1],R2越大,表示模型拟合效果越好。
R2计算方法如下:
式中:Hreal为手动测量真实株高,cm;Hpred为经图像处理后得到的株高,cm;Hmean为所有样本株高预测值的平均值,cm。
取85 组株高数据作为算法模型评估样本,利用上述统计值对不同算法提取的株高准确率进行评估。准确率比较如图4 所示,可以看出,单视角图像株高算法的R2为0.93,多视角图像株高平均算法和最大值算法的R2分别为0.95 和0.96。单张图像因植物枝干倾斜、枝叶遮挡等问题会影响测量准确性,而多视角图像可以很好弥补该缺点,其中最大值算法拟合效果更佳,表现更为稳定,具有更好的鲁棒性。
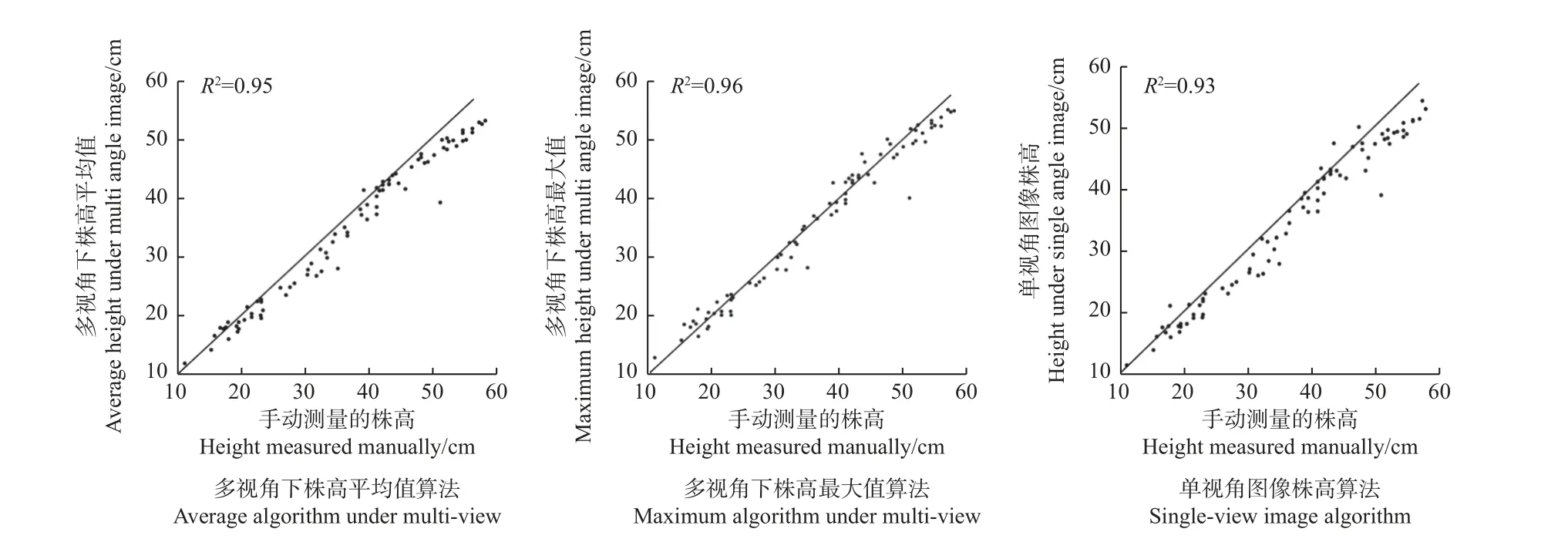
图4 不同株高提取算法准确率比较Fig. 4 Comparison of accuracy of different methods for extracting plant height
2.2 株高可视化
苗木生长周期较长,植物生长缓慢使其无法在短时间内对外界胁迫和环境变化表现出明显的表型改变,通过可视化图像实现“肉眼可见变化”之前看到变化,并进行量化分析,对于监测植物生长、评估植物长势具有重要作用。
本研究提出一种基于灰度图像的株高可视化算法,该算法设定植物根部Hmin处灰度值为0,植物最高点Hmax处灰度值为255(平台Hmax为60 cm),二值化后图像使用该算法的换算公式可以得到图像中各个点的灰度值Gray,转换后的灰度图像采用伪彩色算法进行处理。
可视化换算计算方法如下:
式中:H为多视角图像下提取的株高,cm;Hmax为平台采集的最高株高,cm;Gray 为换算得到的灰度值,无量纲单位。
伪彩色算法处理如下:
式中:r(i,j)、g(i,j)、b(i,j)分别为伪彩色图像红、绿、蓝3 种分量数值,无量纲单位;I(i,j)为图像灰度值,无量纲单位;TR、TB、TG分别为灰度级与R、G、B 三基色的线性映射关系。
2.3 二维图像下主茎直径提取
主茎是植物主枝干去除叶片外的茎干部分,主枝在分枝根部上方1 cm 左右的直径称为主茎直径。本研究以主茎直径表征簸箕柳苗期生长变化。由图5基径与主茎直径位置可以看出,主茎直径处有很多细小叶片遮挡,同时因簸箕柳枝条柔软,形态结构复杂,使得霍夫直线检测等方法不完全适用,且其颜色与茎干相近,也无法通过传统单一色彩因子参数阈值分割算法进行分割,其复杂的边缘信息使得边缘检测效果不尽如人意。

图5 单株簸箕柳主茎直径提取方法示意Fig. 5 Diagram of main branch diameter extraction method of Salix suchowensis
针对上述问题,本研究基于先前的植物茎叶色彩模型(Y=-8.51×lgG+11.68×R-26.48×G+18.30×B+2.81×G/R+3.85×G/B+40)进行主茎直径部分提取。利用缩放比例,截取自图像底部约3 cm 范围的图像,对图像进行开运算以去除噪点和部分叶脉,仅选取保留最大轮廓,实现底部枝条茎部分的提取。采用像素计数法计算图像中每行白色像素个数,因部分叶脉根部与茎部模型值结果相近,不能完全去除,考虑到该情况,将所有行白色像素个数按大小排列,取列表的中位数作为主茎直径像素值,去除因叶片遮挡使茎直径变小及因未去除叶脉造成茎直径变大形成的误差。
2.4 多视角图像下主茎直径提取
采用像素计数法计算所有图像主茎直径的平均值偏差,并设定真实主茎直径阈值范围,以剔除过大异常值,剩下的主茎直径取平均作为多视角图像下主茎直径提取结果。单视角和多视角提取方法的主茎直径准确率评估结果如图6 所示,可以看出单视角图像株高算法的R2为0.16,单张图像处理得到的像素值均为整数,呈多直线形分布,重合点多,且有时图像截取计算得到中位数处仍有细小叶脉,或插条部分,或未能很好去除叶片部分,无法准确提取到主茎直径,出现一对多的错误匹配情况。多视角提取算法则抛去截取失误图像,其R2为0.74,取得较好效果,是可行的主茎直径估测方法。

图6 单视角和多视角提取方法的主茎直径准确率评估结果Fig. 6 Evaluation results of main branch diameter accuracy of single and multi-view extraction methods
3 水肥处理下簸箕柳苗木表型变化
水分和肥料是影响植物生长的重要因素,植物在不同环境条件下的养分吸收能力可反映植物发育对环境的响应规律和资源分配对策(Kerkhoffet al.,2006)。本研究通过采集苗木时间序列图像获得表型参数变化,从而分析水肥对簸箕柳等速生苗木生长发育的影响。
3.1 水肥处理下苗木表型参数的变化
由图7 可知,簸箕柳苗木平均株高和主茎直径受梯度氮肥与水分处理影响。正常水分处理下,簸箕柳苗木平均株高和主茎直径分别为33.6 cm 和2.49 mm,干旱胁迫处理下,簸箕柳苗木平均株高和主茎直径分别为22.8 cm 和2.25 mm,正常水分处理的平均株高和主茎直径相较于干旱胁迫处理高出47.4%和10.7%。该结果表明,干旱会对苗木生长造成显著性影响,严重影响植物体中的物质运输和积累,使得植物平均株高和主茎直径小于正常水分下的平均株高和主茎直径(李聪聪等,2021;彭思利等,2021,谢佰承等,2021;孙明升等,2020)。
正常水分与干旱胁迫处理下梯度氮肥使簸箕柳苗木平均株高和主茎直径均呈先增长后下降的趋势。正常水分处理下,所有处理组簸箕柳苗木平均株高和主茎直径相较于CK 组均有所提高,T2 处理组增长最为显著,平均株高和主茎直径相较于CK 组显著提高15.0%和14.5%;干旱胁迫处理下,T2 处理组增长也最为显著,平均株高和主茎直径相较于CK 组显著提高20.1%和9.9%,而T4 和T5 处理组因水分缺失和氮元素过量的双重影响,平均株高相较于CK 组分别降低7.1%和30.7%,平均主茎直径相较于CK 组分别降低3.9%和14.4%。T5 处理组植物出现机械性损伤,细胞和器官死亡,植物停止生长发育,且主茎直径因缺水出现萎蔫。
选择施肥后第6、12、18 天为时间节点,探究时间序列下簸箕柳苗木株高和主茎直径生长速率(图8),其中第6 天T1 与T2 处理组相同,T3 与T4 处理组相同,第12 天T2、T4、T5 处理组完成再次施加氮肥。

图8 簸箕柳苗木施肥后时间序列生长速率Fig. 8 After fertilization time series growth rate of Salix suchowensis seedlings
第6 天时,正常水分处理下,簸箕柳苗木株高随氮肥含量增加而增加,T5 处理组株高生长速率最高,达2.08 cm·d-1,相较CK 处理组提高68.9%;主茎直径随氮肥含量增加先增加后降低,T2 处理组主茎直径生长速率最高,达0.075 mm·d-1,相较CK 处理组提高1.60 倍。这表明植物不同部位对氮肥含量需求并不相同,苗期时氮多用于茎叶等幼嫩器官发育,根茎对氮需求不明显,出现氮肥含量过高抑制主茎直径生长的现象。干旱胁迫处理下,簸箕柳苗木株高和主茎直径生长速率相较正常水分处理明显降低,随氮肥含量增加先增加再降低直至停止生长,T1 处理组株高和主茎直径生长速率最高,分别达2.26 cm·d-1和0.051 mm·d-1,T5 处理组因水分缺失和氮肥含量过高植物已停止生长,且主茎直径出现萎缩,生长速率甚至为-0.02 mm·d-1。
第12 天时,T2、T4、T5 处理组完成再次施加氮肥,正常水分处理下,簸箕柳苗木株高随氮肥含量增加而增加,T2 和T4 处理组因再次施加氮肥,相较T1 和T3 处理组分别提高7.9%和13.0%,T5 处理组株高生长速率仍最高,达2.37 cm·d-1,相较CK 处理组提高42.3%;主茎直径随氮肥含量增加先增加后降低,T3 处理组生长速率最高,达0.075 mm·d-1,相较CK 处理组提高1.27 倍;与第6 天一样,T5 处理组因水分缺失和氮肥含量过高植物已停止生长。干旱胁迫处理下,与第6 天生长规律相同,簸箕柳苗木株高和主茎直径生长速率相较正常水分处理显著降低,随氮肥含量增加先增加再降低直至停止生长,T2 处理组因再次施加氮肥,株高和主茎直径生长速率最高,分别达2.26 cm·d-1和0.075 mm·d-1。
第18 天时,正常水分处理下,簸箕柳苗木株高随氮肥含量增加先增加后降低,T3 处理组株高生长速率最高,达2.17 cm·d-1,T5 处理组株高生长速率明显下降,为1.77 cm·d-1,这说明植物对氮肥含量存在一定耐受性,超过耐受点后会抑制植物生长;主茎直径随氮肥含量增加先增加后降低,T3 处理组主茎直径生长速率最高,达0.065 mm·d-1;与第12 天一样,T5 处理组因水分缺失和氮肥含量过高植物已停止生长。干旱胁迫处理下,与第6、12 天生长规律相同,簸箕柳苗木株高和主茎直径生长速率相较正常水分处理显著降低,随氮肥含量增加先增长再降低直至停止生长,T2 处理组株高和主茎直径生长速率仍最高,分别达2.17 cm·d-1和0.065 mm·d-1。
总体来看,正常水分处理下,除T5 处理组外,其他组平均株高和主茎直径均高于CK 处理组,其中T2处理组生长速率最高,平均株高和主茎直径生长速率分别为1.92 cm·d-1和0.075 mm·d-1;干旱胁迫处理下,T1、T2 处理组平均株高高于CK 处理组,其中T2 处理组平均生长速率最高,平均株高和主茎直径生长速率分别为1.05 cm·d-1和0.035 mm·d-1。这表明,干旱胁迫下植物生长发育出现明显迟缓现象,随干旱程度加剧差异性更加明显,小部分植物出现机械性损伤甚至死亡。不同梯度氮肥处理下,植物对氮肥吸收具有一定限度,氮肥施加过多会抑制植物快速生长甚至烧伤植物,T2 处理组簸箕柳苗木能够长期稳定快速发育,可作为苗木科学施用氮肥的参考。
3.2 水肥处理下苗木表型变化的可视化
水肥胁迫下,植物表型差异有时并不明显,肉眼很难观察,从时间序列图像中提取表征苗木在梯度氮肥下应激反应能力的株高形态指标,应用可视化方法可呈现不同处理下株高的动态变化。图9 为正常水分处理下T2、T5 处理组簸箕柳苗木株高在时间序列上的变化,图9a、c 为平台采集的彩色图像,图9b、d为应用可视化算法后的株高可视化图像。可以看出,正常水分处理下,T2 处理组簸箕柳苗木上层叶尖逐渐转变为深红色,说明随着时间推移,株高不断增加;T5 处理组42 天后簸箕柳苗木上层叶尖颜色停留在黄色,说明氮肥过高导致植物停止生长发育。株高可视化将簸箕柳苗木在时间序列上的生长变化情况清楚呈现出来,便于直观观察出植物生长发育情况。

图9 簸箕柳苗木随时间变化株高可视化示意Fig. 9 Visual diagram of plant height of Salix suchowensis seedlings changing with time
4 结论
1) 设计并构建面向簸箕柳苗木的多视角表型信息采集平台,通过拍摄多视角图像,避免植物枝叶遮挡等问题,提出近似拼接算法解决高角度和低角度图像的拼接问题,多视角图像下株高和主茎直径提取算法的R2分别为0.96 和0.74。
2) 整个生长周期时间序列下,簸箕柳苗木平均株高和主茎直径受不同水分处理显著影响,正常水分处理下平均株高和主茎直径相较干旱胁迫处理提高47.4%和10.7%。
3) 时间跨度上的簸箕柳苗木序列图像处理结果表明,簸箕柳苗木平均株高和主茎直径在梯度氮肥含量下出现差异性,株高和主茎直径生长随氮肥含量增加呈先增加后降低趋势。
