毛白杨人工林物候特征和生长对施肥的可塑性响应*
2023-08-09李广德刘国彬郭丽琴姚砚武
王 烨 李广德 刘国彬 廖 婷 郭丽琴 姚砚武 曹 均
(1. 北京市农林科学院林业果树研究所 北京 100093;2. 国家开放大学农林医药教学部 北京100039)
表型可塑性可定义为相同基因型在不同环境下产生的不同表型,是生物对环境的适应(Sultan, 2000)。植物在生长发育过程中对外界的反应可以是主动或者被动的(Valladareset al., 2007)。被动过程是指受个体发育影响,植物表型会随着年龄增加和个体大小而发生变化,称为表观可塑性,如植物的根冠比会随着年龄的增加而逐渐减小。植物的个体发育可表示为异速生长关系,生物体多种特征之间普遍存在异速生长。异速生长是生物体不同特征之间的一种固定的不成比例生长关系,是由物种遗传性所决定的一种固有特性(Weiner, 2004)。主动过程是指植物为适应外界环境而发生的表型改变,称为真实可塑性(McConnaughayet al., 1999)。表型可塑性的关注点是植物对外界的反应,但有研究认为可观察到的真实可塑性却不常见(Fisheret al., 2005),即使处于变化的环境中,植物的表型改变主要还是由个体发育控制(Mülleret al., 2000)。
施肥是重要的森林集约经营技术措施(胡建文等,2020),尤其对于速生树种。研究表明,施氮肥可提高短轮伐林40.3%的产量(Fabioet al., 2018);同时,施肥后能在短期内引起土壤养分的显著增加,为应对环境的变化,植物的一些表型可能会产生可塑性响应。例如,施肥后会引起植物阶段性生长的改变(Possenet al., 2021);根系生物量分配比例往往会随着土壤氮养分的减少而增加(Giardinaet al., 2003),在氮养分充足时,光合产物则更多的分配到地上部分(王力朋等,2012)。也有研究认为,施肥引起植物表型变化是因为加快了植物个体发育,并非因可获得资源的增加而做出表型调整策略(Coyleet al., 2005)。由此可见,环境因子是否能引起植物表型的“真实改变”这个问题依然存在争论。
前期研究发现,施肥能够提高毛白杨林分的胸径生长量和地上部分生物量,对地下生物量则无明显影响(席本野等, 2017),从而引起了地上地下生物量分配比例的改变,这种现象可能是植物生长表型对养分添加的真实响应。为进一步探明植物生长对施肥响应的作用机制,本研究以我国速生丰产林重要造林树种毛白杨(Populus tomentosa)为研究对象,设置不同施氮量处理试验,针对以下目标开展研究:1)分析施肥对毛白杨叶生长和胸径生长物候特征的影响;2)运用异速生长方程分析施肥对毛白杨生物量分配模式的影响。该研究发现将为我国杨树及其它树种用材林高效养分管理技术的制定与优化提供理论依据。
1 研究地概况
研究地位于鲁西北中部黄泛冲积平原,山东省聊城市高唐县(36°58′N,116°14′E)。气候类型为暖温带半湿润季风气候:年平均气温为13.8 ℃,春季干旱,夏季炎热,月平均气温最高在7 月,约为27.2 ℃,月平均气温最低出现在1 月份,约为-1.5 ℃。多年平均降水量589.3 mm,主要集中于7—8 月。年均蒸发量(水面)2 249 mm。全年日照总时数2 652 h,无霜期191~217天。试验地土壤类型为潮土,土壤质地在 0~120 cm土层为粉土,120 cm 以下为粉壤土。试验地 0~90 cm浅土层的理化性质见表1。

表1 试验地土壤物理和化学性质Tab. 1 Soil physical and chemical characteristics at the experimental site
试验时间为2010—2012 年,研究对象为三倍体毛白杨人工林。造林时间为2008 年春季,苗木为1 年生三倍体毛白杨无性系B301((P. tomentosa×P.bolleana)×P. tomentosa)裸根苗。采用宽窄行栽植模式造林,窄行株行距(林带内)为1.5 m×1.5 m,宽行(带距)行距8 m,种植密度为1 404 株·hm-2。2008—2009年对林分进行常规灌溉管理。试验开始前,林分平均胸径5.0 cm,平均树高6.67 m。采用滴灌系统进行灌溉施肥,滴灌管布设在宽行内距树体60 cm 的区域,滴灌管埋深20 cm,滴头间距为50 cm,流量为2 L·h-1。灌溉水源为地下水。
2 研究方法
2.1 试验设计
施肥试验包括施氮(N)量和施肥频率两个因素,肥料为尿素(46% N)。施N 量设3 个水平,分别为115(N115)、230(N230)和345(N345)kg·hm-2a-1;施肥频率设2 个水平,分别为生长季内施肥4 次(F4)和施肥2次(F2),共计6 个施肥处理,并设置一个不施肥对照CK。前期研究(席本野等, 2017)发现,施肥频率对毛白杨生长和生物量无显著影响,与施肥量也无交互作用,因此本研究将2 个施肥频率进行合并,仅分析施肥量因素。试验采用完全随机区组设计,重复3 次,每个重复小区含80 棵树,小区外侧2 条林带为保护行,共计21 个小区。施肥时先把肥料溶于水配置成含氮肥料溶液(浓度为86 g· L-1),然后将肥料溶液注入到滴灌系统中,施肥时间为每年5 月初和7 月初。为保证植物的正常生长,除施肥外,在4—7 月自林木展叶时起,对所有处理采取每隔10 天灌水20 mm 的灌溉制度,同时保持对照接受的灌溉量与施肥处理相同。
2.2 试验方法
2.2.1 胸径生长物候特征 2010—2012 年,在各试验小区中间林带随机选定10 株样树,在树干1.3 m 处标记固定测定位置,用胸径尺定期测量生长季内(4—9 月)胸径,每隔15 d 测量1 次,记录胸径生长的起始时间和停止时间。然后,计算不同时期的胸径平均生长速率:
式中,AGR 代表胸径平均生长速率(mm·d-1),DBHincrement为此时期的胸径生长量 (mm),n是对应时期内的天数 (d)。
2.2.2 叶生长物候特征 2011 年4 月分别在各小区随机布设2 个固定测点进行林分叶面积指数(LAI,m2·m-2)的测定,每个处理共计6 个测点,测点位于中间林带窄行中央。采用WinSCANOPY 冠层分析仪(Régent Instruments Inc., Quebec, Canda)进行测定,测定时间为4 月初—10 月底,约每隔15 天测定1 次,记录不同处理展叶时间、落叶时间和不同时期的LAI。
2.2.3 生物量 分别在2010、2011 和2012 年10 月进行生物量的采样工作。2010 年只对CK 处理采样,2011 和2012 年对全部处理进行采样。根据林木生长调查的结果,发现各处理内林木生长势较为一致(胸径变异系数为5.5%~13%,树高变异系数在6.3%~11%),因此,在每个小区选择1 株(每个处理3 株)代表不同处理的平均标准木进行整株采伐。其中,2010年采伐3 株(CK),2011 年和2012 年各采伐21 株,共计采伐45 株样树。根系样品采集范围为:以树干为中心,边长为 1.5 m 的正方形区域,采集深度至地下60 cm 土层。采伐后,将地上部分分为树干、树枝和叶片3 个器官,将地下部分分成根桩、粗根(直径>2 mm)和细根(直径≤2 mm)3 个器官。在田间对不同器官鲜重进行称量,然后采集代表样品带回实验室,在70 ℃下烘干至恒质量,测定样品含水量,以计算不同器官的干物质量。
2.2.4 气象数据 在距离试验地约100 m 处的空地上安装一台全自动气象站(Delta-T Devices Ltd.,Cambridge,England),用于获得降雨量等气象数据。
为切实保障服务质量和服务效果,河北省图书馆对选派人员优中选优,从中层干部中选拔副研究馆员以上职称的业务骨干13名,由分管馆长带队,到受援单位进行前期调研,了解基本情况和基层馆需求,结合各馆实际情况,分别制定了对应的工作方案。13名业务骨干以分组的形式到各受援图书馆开展服务,每组选定一名组长,负责整体工作的组织协调和安排部署,轮流、定期上门提供工作指导、业务辅导与讲座培训。最终建立了“前期有调研、过程有日志、完成有总结、结束有评价”的工作机制。
2.3 数据分析
2.3.1 施肥与降雨对胸径生长的交互作用 利用公式(2)分析降雨对施肥效应的影响,计算生长季不同时期施肥处理相对于CK 胸径增长量的提高比例。
式中,IFi表示滴灌施肥处理在时期i的胸径生长量,ICi为CK 处理在时期i的胸径生长量,DIF 表示不同时期施肥处理较CK 胸径增长量的提高比例。将DIF与对应时期降雨量做相关性分析,以研究胸径生长对施肥的响应是否受降雨影响。
2.3.2 生物量异速增长分析 采用异速生长方程分析不同处理毛白杨不同变量之间的异速增长关系:
式中,y和x代表不同变量。将植物分成叶、茎(枝+干)、地上(叶+枝+干)和地下(根桩+粗根+细根)4 个部分,分析生物量在这些部分间的分配关系。首先对上式做自然对数转换,将二者的关系转化为线性关系公式(4):
式中,lnβ 为异速生长常数(allometric constant),表示为在Y 轴上的截距,α 为异速指数(scaling exponent),表示为方程的斜率;当α=1 时,x和y之间是等速增长关系;当α≠1 时,x和y之间则为异速增长关系。异速方程的斜率α 表示的是不同器官在生物量分配上的比例关系,截距β 表示二者在绝对量上的不同。
将数据进行转换后,采用简化主轴方法(reduced major axis, RMA)对上式的 β和 α进行拟合求值。RMA被认为是异速方程的最有效的专用分析方法(Falsteret al., 2006)。用本方法拟合出的β 和α 值被认为比一般线性回归法更能表征2 个变量之间的关系。使用SMATR 程序包(Falsteret al., 2006)运行RMA 分析过程。该程序同时还可以对多条直线的斜率和截距的数值(α 和β)进行差异显著性检验,如数据间差异显著,可进行多重比较。该方法还可以给出α 和β 的95%置信区间。
3 结果与分析
3.1 林分叶生长物候特征
由图1 可知,施肥处理(N115、N230和N345)林分叶面积变化与CK 相似,处理间的差异在2011 年整个生长季内均不显著(P>0.05)。毛白杨从4 月10 日左右开始展叶,4 月中旬至5 月中旬是树冠叶片的迅速增长期:4 月中旬不同处理平均LAI 为0.47 m2·m-2,到5月中旬增至1.62 m2·m-2。5 月中旬—8 月叶面积指数较稳定,变化范围在1.60~1.77 m2·m-2之间。8 月末进入落叶期,叶面积迅速下降。生长期内(4—8 月),N115、N230和N345处理的平均LAI 分别为1.64、 1.63 和1.64 m2·m-2,CK 处理为1.58 m2·m-2,施肥处理仅略大于CK,但未达到显著水平(P>0.05)。然而,在落叶期(8 月底之后),CK 林分的LAI 下降速度则要小于各施肥处理。

图1 2011 年施肥处理与CK 处林分LAI 的动态变化Fig. 1 The dynamic changes of stand LAI in fertilization and control (CK) treatments in 2011
3.2 胸径生长物候特征
试验地毛白杨胸径生长开始于4 月中旬,9 月停止生长(图2a),胸径生长节律呈现明显“快-慢-快”的双峰型生长模式(图2b):5 月初至中旬是第1 个速生期,5 月中旬至6 月生长速率整体下降;7 月初至中旬是第2 个速生期;8 月生长速率下降,8 月中旬至9月初,进入枝条封顶期,停止生长。2012 年同时期的生长速率多数小于2011 年同期。2 年的胸径生长监测数据显示,施肥并没有改变毛白杨的胸径生长物候特征,但施肥处理整体的胸径生长速率大于CK(图2),这种差异在生长季后期更为明显。具体而言,施肥处理的生长速率,在2011 年8 月平均比CK 高71.6%(P<0.05),在2012 年7 月和8 月分别比CK 高30.3%和85.1%(P<0.05),而不同施肥量处理之间的差异不明显(P>0.05)。
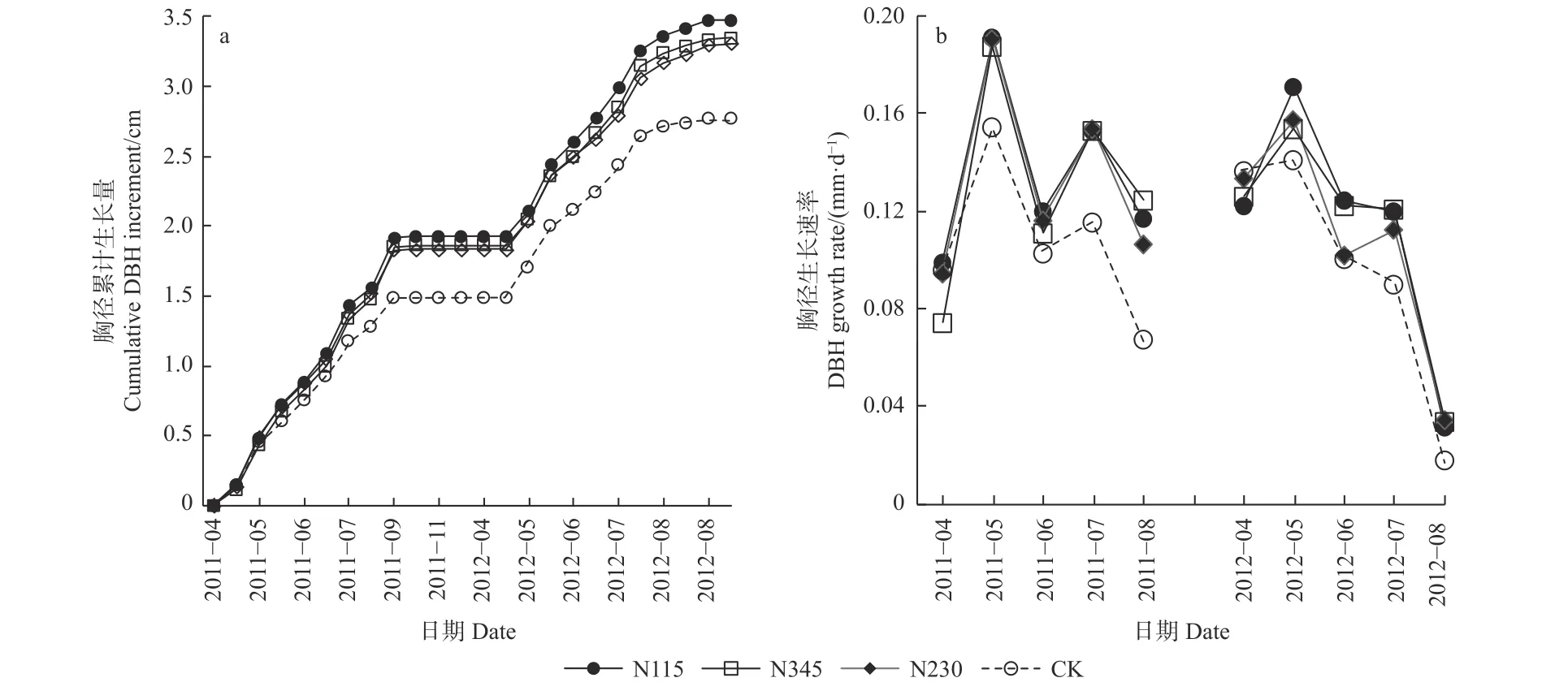
图2 2011—2012 年不同处理(a)胸径累计生长量和(b)胸径生长速率Fig. 2 The cumulative DBH increments (a) and the periodic DBH increment rates in different treatments in 2011 and 2012
2010、2011 和2012 年试验地的总降雨量分别为791.2、593.6 和799.2 mm,最大降雨量出现在7 月和8 月,这2 个月的降雨量占总降雨量的33%和38%(图3)。施肥处理与降雨对胸径生长有明显的交互作用,二者呈显著的二次曲线关系(R2=0.59,P<0.001)。具体表现为当月降雨量在0~200 mm 范围内时,DIF的值随着降雨量的增加逐渐增加,而当月降雨量接近400 mm 时,DIF 的值又迅速下降(图4)。

图3 2010—2012 年月降雨量分布Fig. 3 Monthly precipitation distribution in 2010—2012

图4 施肥与降雨对胸径生长的交互作用Fig. 4 The interaction effect between fertilization and precipitation on the growth of DBH
3.3 异速生长分析
由表2 可知,毛白杨不同器官之间的异速生长关系极显著(P<0.01)。CK 和施肥处理的异速方程斜率α 无显著差异(P>0.05),即施肥未改变异速方程中的异速指数。当不同处理间的斜率和截距均无显著差异时,以处理间的共同斜率和截距进行合并分析。CK 和施肥处理“地上—地下”的方程斜率均为0.7,二者的“茎干—胸径”和“叶—胸径”的共同斜率分别为2.4 和 3.5,均与1 有显著差异(P<0.05)。当异速指数α≠1 时,表示变量之间的增长关系为异速增长,即随着年龄的增长,生物量分配比例受个体发育的控制而发生改变。CK 和施肥处理的“叶—地下”的方程斜率分别为1.2 和1.0,与1 无显著差异(P>0.05)。当异速指数α=1 时,表示这两者为等速增长关系,即随着年龄的增长,叶和地下这2 个部分的分配比例不会发生相对变化。
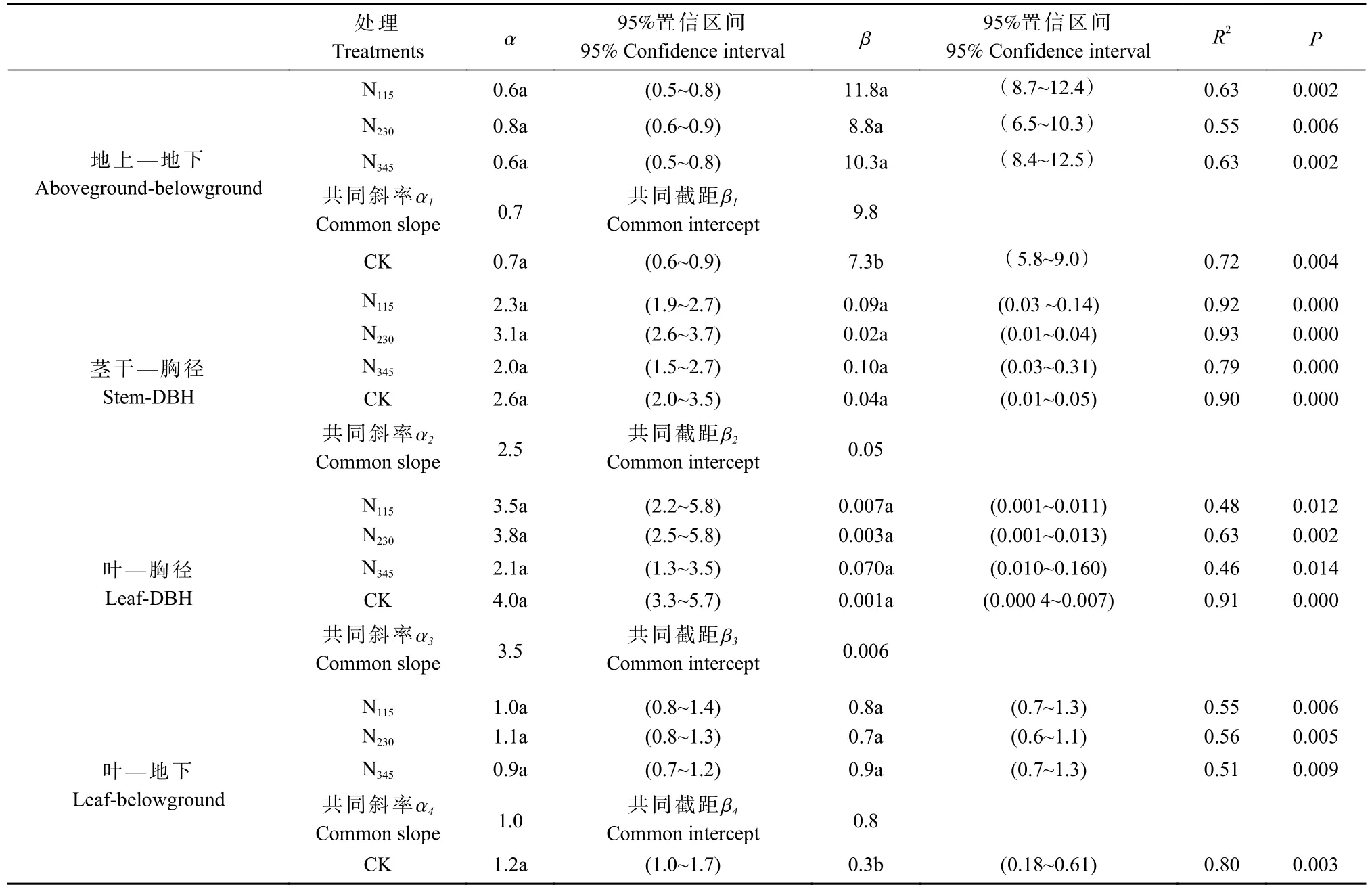
表2 不同处理生物量异速生长方程差异显著性检验①Tab. 2 Test for significant differences in slope and intercept of the allometric equation of biomass among different treatments
由于“叶—地下”的生物量分配为等速增长,所以生物量分配比例的改变取决于异速生长常数β。如表2 所示,施肥虽然没有改变生物量分配异速方程的斜率,但施肥处理“叶—地下”的异速生长常数β 与CK 有显著差异(P<0.05),施肥处理之间无显著差异(P>0.05)。各施肥处理“叶—地下”生物量分配异速方程β 值比CK 显著高出0.4~0.6,表明当拥有相同根系生物量时,施肥比CK 有着更多的叶生物量,即施肥使得生物量分配从根向叶转移。此外,“地上—地下”的方程截距也有不同程度地沿y 轴上升。
4 讨论
4.1 生长物候特征对施肥的可塑性响应
植物的物候特征会受到多种环境因子如气候和管理措施的影响,而具有一定的可塑性(Rossiet al.,2006; Duchesneet al., 2012; Primiciaet al., 2013; Allenet al., 1993; Gouldet al., 2012)。施N 肥能明显改变木本幼苗和草本植物的物候特征(Olietet al., 2011; Xiaet al., 2013),然而,本研究结果显示,施N 肥以及改变施N 量并未改变毛白杨林分叶生长和木质部生长的物候特征。在试验前,笔者曾假设施肥可能会显著提高毛白杨在6 月(最高耗水月份)的胸径生长速率,但试验结果并不支持这个假设。此外,本研究发现施肥使得毛白杨胸径生长物候特征发生了表观可塑性的改变,即施肥处理和CK 虽有相似的胸径生长节律,但施肥处理在各时期的胸径生长速率高于CK,且这种差异在生长季后期(7—8 月)达到显著水平。栓皮栎的木质部生长受N 添加和生长期交互作用影响显著,但对同属树种麻栎的影响不显著(Yuet al., 2019);在马尾松的施肥研究中,也有学者发现施肥对茎干生长物候特征并无明显影响(Zhanget al., 2017)。上述不同研究结果出现差异的原因可能是草本植物和幼苗对土壤养分的添加更为敏感(Nasholmet al., 2009)。有研究表明,物种和木质部结构的差异会导致植物茎干生长物候对环境变化的敏感性不同(Chenet al., 2022)。当木本植物的茎干生长节律对养分增加没那么敏感时,水分却能对其有显著作用(D’Orangevilleet al.,2022)。灌溉后火炬松生长季封顶时间会推迟,由于生长期的延长,灌溉处理的林木生长量也更高(Gonzalez-Beneckeet al., 2010)。针对毛白杨的灌溉研究表明,灌溉提高了毛白杨胸径生长量,但同样对生长物候特征无影响(席本野等, 2012; Liet al., 2020)。因此,本研究认为毛白杨生长物候特征主要受树种基因型或者其他环境因素的控制,而不受施肥灌溉的影响。
本研究发现,施肥对不同时期胸径生长的促进作用不是独立的,而是会受到降雨的影响。当月降雨量在200 mm 范围以内时,施肥对林木生长的促进效应(即施肥相对于CK 胸径生长量的提高比例)与降雨量呈正相关关系,而当月降雨量大于300 mm 时,这种促进作用又急剧下降。过多降雨引起施肥促进效应下降的原因,可能是土壤水分含量过高造成土壤通气性下降,从而导致根系缺氧而抑制了根系对养分的吸收(Schuur, 2003)。Van den Driessche 等(2003)在针对山杨幼苗的施肥试验中也发现,山杨生长对施肥的响应受降雨的影响较大。除了降雨,其他环境因子也曾被发现与施肥对植物的物候有协同效应,最常见的为温度。例如,在土壤增温和施肥的协同作用下,黑云杉木质部的生长时间会延长(Lupiet al.,2012)。由此可见,环境因子与林地管理措施之间存在复杂的交互作用,因此今后在制定和优化相关森林管理技术措施时,应将环境因素对技术措施应用效果的影响考虑在内。例如,在试验地条件下,当月降雨量大于300 mm 时,应考虑排水措施来提高肥效,或者应避免在降雨频繁时进行施肥。
4.2 生物量分配对施肥的可塑性响应
当毛白杨林分的LAI 和胸径生长物候特征对施肥的响应仅在表观的可塑性时,其生物量分配模式对施肥的响应则表现较为复杂。本研究的异速生长分析结果表明,施N 肥仅引起了毛白杨叶片和根系的生物量分配发生了“真实改变”:在具有相同质量根系的情况下,施肥林分拥有比CK 林分更多的叶片生物量,林木茎干的生物量分配则受个体发育的影响更大。总体来说,施肥能部分改变毛白杨生物量的分配模式,但这种作用会受到个体发育的严格控制。许多在其他植物种类上[阔叶树种(Heet al., 2020)、针叶树种(Kinget al., 1999)和草本植物(Zhouet al., 2014)]开展的类似研究结果也表明,植物生物量分配模式对施肥产生的响应会因器官不同而异。除土壤养分以外,在植物对水分和光照响应的研究中也有类似发现(McCarthyet al., 2007; Schallet al., 2012)。这些研究结果表明,植物生物量分配的个体发育作用和对环境资源的响应之间并不相互排斥(Niklas, 2006)。然而有些研究认为,施肥后会提高植物的生长速率,加速了植物的个体发育进程从而引起生物量分配模式的变化,并非施肥本身直接导致的结果(Coyleet al.,2005; Colemanet al., 2004; Coyleet al., 2008)。造成这些研究结论差异的一个重要因素是不同植物不同器官生物量的异速增长关系并不是相同的。在本研究中,毛白杨叶和地下部分的异速生长关系为等速生长,也就是叶和根的生物量分配不会随着年龄的变化而改变。当个体发育的作用可忽略时,生物量分配的任何改变都可视为真实的改变(McConnaughayet al.,1999),表明毛白杨的叶器官对施肥具有较强的表型可塑性响应。
本研究还发现,毛白杨茎干生物量和胸径、地上与地下生物量分配的关系为异速增长关系,即支配这些变量变化的主要驱动力是个体发育(Weiner, 2004)。异速增长关系是植物在长期的进化过程中形成的稳定分配策略,是基因型决定的结果,对可获得资源(光、水和养分)的变化不敏感(Weiner, 2004; Mülleret al.,2000)。因此,在本研究中,施肥处理与CK 茎干生物量分配比例的差异是由于施肥加快了茎干生长速率而引起的,并非茎干生物量分配发生了“真实变化”。茎干生物量分配的“表观变化”也验证了胸径生长物候特征的“表观变化”,对于这种现象笔者也进一步提出一个有待验证的假设:如果植物茎干生长节律发生显著变化,那么茎干生物量分配的真实改变则有可能发生。
5 结论
毛白杨生长和物候特征对施肥的响应是复杂的,施肥虽然能明显提高不同阶段的胸径生长速率,但不能改变胸径的生长节律。毛白杨茎干生物量和胸径、地上和地下之间的生物量分配关系为异速分配,主要由个体发育作用控制。由此表明,施肥能引起毛白杨茎干生长速率和生物量分配的表观变化,但施肥后茎干生物量分配比例的变化是由于施肥加速了林木的个体发育而造成。同时,施肥能引起叶和地下生物量分配的真实变化,使林木将更多的生物量分配到叶器官中,且该作用并不依赖于林木的个体发育。综上,施肥条件下,毛白杨的生长表型不仅会发生真实的可塑性变化,而且也会受到个体发育改变的影响。本研究发现不仅可帮助深入理解速生树种对环境养分变化的响应和适应策略,而且还可为“基于植物可塑性响应策略”的速生丰产林高效养分管理技术的优化(例如,应根据林分收获目标的差别来调整施肥策略)提供理论依据。
