钙镁叶面肥缓解妃子笑荔枝果肉退糖现象的呼吸机理
2023-07-22杜婧加彭俊杰陈甜甜税贤廖海枝林晓凯周开兵
杜婧加 彭俊杰 陈甜甜 税贤 廖海枝 林晓凯 周开兵


摘要:【目的】探討叶面喷施钙镁肥对妃子笑荔枝果肉糖酵解(EMP)—三羧酸循环(TCA)途径呼吸速率的影响,为荔枝生产提供理论参考。【方法】设置以树冠喷布0.3% CaCl2水溶液(Ca)、0.3% MgCl2水溶液(Mg)、0.3% CaCl2+0.3% MgCl2水溶液(Ca+Mg)的3组处理,以树冠喷布清水为对照(CK),测定、比较不同发育期荔枝果肉风味品质主要指标、EMP-TCA途径呼吸速率及其关键酶活性的动态变化。【结果】荔枝果肉可溶性糖含量在35~56 d呈上升趋势,在花后63 d之后,除Ca+Mg处理的果肉可溶性糖含量未显著下降(P>0.05,下同)而缓解退糖现象外,其他处理均出现退糖现象。所有处理的果肉可滴定酸含量在采收期前后均无显著变化,且相互间无显著差异;Ca处理和Ca+Mg处理的EMP途径呼吸速率显著低于Mg处理和CK(P<0.05,下同),而在TCA途径呼吸速率上,Ca+Mg处理显著低于Ca处理,故Ca+Mg处理通过抑制EMP-TCA途径呼吸速率进而抑制果肉总呼吸速率;采收期前后,Ca+Mg处理的磷酸已糖异构酶(GPI)、琥珀酸脱氢酶(SDH)和苹果酸酶(NADP-ME)等活性均显著低于CK;Ca处理对GPI活性具有抑制作用,对PDC活性呈现促进作用;Ca处理和Mg处理的NADP-ME活性显著高于Ca+Mg处理和CK。【结论】Ca+Mg处理通过抑制妃子笑荔枝果肉GPI、SDH和NADP-ME活性而分别抑制EMP和TCA途径,进而降低EMP-TCA途径有氧呼吸速率,减少果肉糖分呼吸损耗,从而减缓果肉退糖现象。
关键词:妃子笑荔枝;钙镁叶面肥;糖酵解(EMP);三羧酸循环(TCA);呼吸速率;可溶性糖;可滴定酸
中图分类号:S667.1 文献标志码: A 文章编号:2095-1191(2023)02-0356-09
Abstract:【Objective】The effects of spraying the calcium and magnesium foliar fertilizer on the respiration rate of Feizixiao litchi pulp glycolysis(EMP)-tricarboxylic acid cycle(TCA) pathway was explored to provide reference for litchi planting. 【Method】During the period of fruit development, crowns of Feizixiao plants were sprayed with 0.3% magnesium chloride(Mg),0.3% calcium chloride(Ca) and 0.3% CaCl2+0.3% MgCl2(Ca+Mg), and water as control(CK),indicators of pulp flavor and quality at different developmental stages, the dynamic changes of EMP-TCA pathway respiration rate and key enzyme activities were detected and compared. 【Result】The soluble sugar content of litchi pulp showed an upward trend from 35 to 56 d. After 63 d after flowering, except for the soluble sugar content of pulp treated with Ca+Mg did not decrease significantly (P>0.05, the same below), and sugar withdrawal occurred in all other treatments. The titrata-ble acid content of all treated pulp did not change significantly before and after the harvest period, and there was no significant difference between them. The respiration rate of EMP pathway of Ca treatment and Ca+Mg treatment was significantly lower than that of Mg treatment and CK(P<0.05, the same below),while the respiration rate of TCA pathway in Ca+Mg treatment was significantly lower than that of Ca treatment,so Ca+Mg treatment inhibited the total respiration rate of pulp by inhibiting EMP-TCA respiration rate. Around the harvest time, the activities of phosphate hexose isomera-se (GPI), succinate dehydrogenase (SDH) and malic enzyme (NADP-ME) treated with Ca+Mg were significantly lower than those treated with CK. Ca treatment inhibited GPI activity and promoted PDC activity; the activity of NADP-ME in Ca treatment and Mg treatment was significantly higher than that in Ca+Mg treatment and CK. 【Conclusion】In conclusion, Ca+Mg treatment inhibits the EMP and TCA pathways by inhibiting the GPI, SDH and NADP-ME activities of the pulp of Feizixiao litchi, thereby reducing the aerobic respiration rate of EMP-TCA and reducing the respiration loss of sugar in the pulp, thereby alleviating the sugar-dropping of the pulp.
Key words: Feizixiao litchi; foliage spraying of calcium magnesium fertilixer; glycolysis pathway(EMP); tricarboxylic acid circulation(TCA); respiration rate; soluble sugar; titratable acid
Foundation items: National Natural Science Foundation of China(31960570); Hainan University World Class Discipline Construction Fund Special Project(RZZX201906); Hainan Graduate Innovation Project(Hys2019-90)
0 引言
【研究意义】荔枝(Litchi chinensis Sonn.)产业是我国热带南亚热带果业主要支柱产业,其中,妃子笑荔枝是我国的主栽品种之一,也是海南产区栽培面积最大的品种,研究其果实品质发育问题有益于促进荔枝产业和华南区经济发展(倪耀媛和吴素芬,1990)。妃子笑荔枝以果实风味见长,但存在果皮果肉发育不同步的现象,即果肉含糖量最高时果皮着色不良,而果皮完全转红着色时,果肉衰老导致糖类物质氧化分解,糖分下降,果味发涩,出现果肉退糖(果皮滞绿)现象,该问题影响了妃子笑荔枝的市场竞争力和经济价值(张锐等,2014;吕润等,2021)。课题组前期的实验结果表明,在果实生长发育期间,喷施钙镁混合液可减缓妃子笑荔枝果肉的退糖现象(周晓超等,2015),但关于其减缓退糖的代谢生理机制问题研究报道较少。因此,通过深入研究钙镁单一和混合叶面营养对妃子笑荔枝果肉呼吸作用的影响,可有助于了解果肉退糖现象发生时可溶性糖的去向和减缓退糖时的呼吸代谢生理机制,可丰富和发展妃子笑荔枝果实生长发育理论,为果实着色不良的荔枝品种遗传改良和栽培技术优化打下理论基础。【前人研究进展】植物的呼吸代谢是一种复杂且强大的反应体系,能氧化不同的碳底物,不仅能产生能量,还可生成碳骨架支撑蛋白质等物质合成(Vanlerberghe et al.,2020)。植物的呼吸途径主要包括糖酵解(EMP)、三羧酸循环(TCA)、磷酸戊糖(PPP)和电子传递链,其中,机体能量主要由糖酵解—三羧酸循环(EMP-TCA)途径提供(Analin et al.,2020)。在有氧条件下,EMP途径的最终产物丙酮酸进入线粒体进行TCA途径,该途径是糖类、蛋白质和脂肪的共同氧化途径,同时也是与其他代谢途径连接的枢纽,对植物体的代谢调控具有重大意义。相关研究发现,油菜素内酯(BRs)和1-甲基环丙烯(1-MCP)可明显减低杨桃呼吸速率,延缓其成熟衰老(陈宇新,2017);低钙处理显著抑制芒果果实的呼吸速率,提高其果实贮藏性(姚智等,2017)施用氮素可提高甜菜根部呼吸速率從而促进块根增长(于超,2021)。课题组前期研究结果表明,钙镁元素是影响妃子笑荔枝糖酸含量的主要元素(苏阳等,2015),而有机酸的积累对妃子笑荔枝果肉呼吸具有负反馈调节作用(廖海枝等,2021)。糖分是果肉内在营养风味品质的基本构成物质,其在体内的主要分解代谢方式受供氧情况的影响而存在差异,主要包括无氧氧化(EMP)、有氧氧化(TCA)、磷酸戊糖途径(PPP)及糖原分解等,糖类物质经呼吸作用使果肉积累中间产物有机酸和释放CO2而损耗(翁雪成和袁红,2007)。因此,果肉糖类物质的积累可能受呼吸作用调控影响。【本研究切入点】喷施钙镁混合液可减缓妃子笑荔枝果肉的退糖现象,但是其糖分积累与呼吸代谢生理生化机理尚不清楚。【拟解决的关键问题】在前期研究基础上,通过观测荔枝果肉可溶性糖、可滴定酸含量,EMP和TCA途径的呼吸作用相关指标等变化,比较不同施肥处理和对照间差异,分析确定叶面钙镁营养对妃子笑荔枝果肉呼吸作用的影响特点,进一步探析钙镁营养减缓果肉退糖现象的生理生化机理。
1 材料与方法
1. 1 试验地概况
采样地点为海南省临高县金牌农场五队荔枝园(东经109°48′,北纬19°54′),位于海南省西北部,该果园所在产区位于热带季风气候区,其气候特点是高温多雨,光照充足,年平均温23~24 ℃,年平均日照时数2175 h,年平均降水量1100~1800 mm,且雨热同季;土壤为肥沃砖红壤。妃子笑荔枝在该产地主要物候期:花期为2—3月,生理落果期为4月初开始,4月下旬进入果实膨大期,5月中旬进入果实成熟期。
1. 2 试验设计
于2021年4月18日选择生长势一致且无任何不良表现的16年生妃子笑荔枝树20株,分为4组,单株小区,重复5次。3个处理分别是:喷施0.3% CaCl2水溶液、0.3% MgCl2水溶液、0.3% CaCl2+0.3% MgCl2水溶液,分别简记为Ca处理、Mg处理、Ca+Mg处理;以喷施清水为对照(CK)。假种皮包满种子且果蒂出现红色标志着荔枝果实生理落果期结束,此时开始田间叶面喷肥处理,即4月下旬开始叶面喷肥。喷肥时间为2021年4月18日(花后35 d)、4月25日(花后42 d)和5月2日(花后49 d)的上午8:00—9:00。标准参照果的选取:于4月18日分别在每株样树树冠外围间隔大约70°(以树干为圆心)的5个位置,随机选取生长一致且中庸的5个果实作为取样参照果,挂牌标记。此后取样时以标准果的平均纵径、平均横径和着色状态为参照,在树冠外围以相同方法选取类似大小和着色状态的果实作为样品果。从稳果开始至果皮全红时为止,即在花后35 d(4月18日)、42 d(4月25日)、49 d(5月2日)、56 d(5月9日)、63 d(5月16日)、70 d(5月23日)取果样,每次取果25~30个,在田间放入液氮罐速冻并带回实验室,储存于-80 ℃超低温冰箱中备用。
1. 3 测定指标及方法
果肉可溶性糖含量测定参考Wang等(2006)的方法,稍作修改:称取0.5 g果肉,微波炉杀酶30 s,加入乙醇研磨、离心,上清液蒸干、定容至10 mL,经0.45 μm滤膜过滤后待测。将果糖、葡萄糖与蔗糖含量之和作为可溶性总糖含量,使用Waters2695高效液相色谱仪测定糖分含量。采用糖酸仪进行可滴定酸含量测定,测定时果汁稀释至50倍,以柠檬酸作为计算标准进行检测。
果肉EMP-TCA途径呼吸速率使用便携式O2/CO2顶空分析仪(Dansensor CheckPoint3)进行测定(Sivakumar and Korsten, 2010),单位为mL CO2/(kg·h)(Wang et al.,2010);使用10 mmol/L氟化钠作为EMP途径专一性抑制剂,使用50 mmol/L丙二酸作為TCA途径专一性抑制剂。测定时间均在上午10:00左右,环境温度维持在28 ℃左右,每次先测得样品总呼吸速率,真空渗入抑制剂后测定各途径的剩余呼吸速率,由总呼吸速率减去各途径剩余呼吸速率得出单一途径呼吸速率观测值(史青纯,2009;宋丛丛,2021)。
磷酸己糖异构酶(GPI)和丙酮酸激酶(PK)酶液提取参考何亚辉(2007)、史青纯(2009)的方法,略作修改。称取果肉2 g,冰浴环境下加入Tris-HCl缓冲液,研磨、离心,取上清液,用酶标仪在520 nm波长下测定吸光度(OD值)。以果糖为标准物通过标准曲线计算样品酶活性。
琥珀酸脱氢酶(SDH)和丙酮酸脱氢酶(PDC)参考Zhang等(2008)、宋丛丛(2021)的方法制备酶液。称取2 g荔枝果肉,加入含有0.005 mol/L EDTA、0.5 mol/L蔗糖、1 mg/mL牛血清蛋白、0.05 mol/L PBS缓冲液的提取介质,冰浴后研磨。测定酶活性前酶液与反应液1∶1混匀,在30 ℃环境下保温10 min,在600 nm测定吸光度的变化。通过标准曲线计算样品中各酶活性。
苹果酸脱氢酶(NAD-MDH)和苹果酸酶(NADP-ME)等活性测定参考罗安才等(2003)、史青纯(2009)的方法。称取2 g荔枝果肉,加入Tris-HCl缓冲液,冰浴研磨后离心取上清液备用;上清液加入由0.2 mol/L Tris-HCl、0.1%聚乙二醇辛基苯基醚、5 mol/L抗坏血酸合成的反应液,定容至5 mL。参考Sadka等(2000)的方法测定酶活,略有改动。用酶标仪在340 nm波长下测定各酶吸光度(OD值),通过标准曲线计算样品中各酶活性(Sadka et al.,2000;罗安才等,2003)。
1. 4 统计分析
采用SAS 9.1.3对数据进行统计分析。采用ANOVA过程作方差分析,DUNCAN法作多重比较分析。
2 结果与分析
2. 1 妃子笑荔枝果肉可溶性糖和可滴定酸含量变化
2. 1. 1 可溶性糖 如图1所示,所有处理的荔枝果肉可溶性糖含量均在前期呈上升趋势,后期则存在一定差异。其中,Ca处理上升至49 d后无明显变化,Mg处理和CK在花后56 d达峰值;除Ca+Mg处理,CK和Mg处理均出现退糖现象。花后42 d,Ca+Mg处理显著最高(P<0.05,下同),而Mg处理显著最低;花后56 d,Ca处理显著最低;至花后70 d,Ca+Mg处理显著最高,而其他处理较56 d时均呈现下降或平缓趋势。综上,Ca+Mg处理具有促进可溶性糖积累的效果,且在花后56 d之后未出现退糖现象,其他处理均出现退糖现象。
2. 1. 2 可滴定酸 如图2所示,各处理的荔枝果肉可滴定酸含量动态变化均呈L形,即花后35~56 d急剧下降,花后56 d之后无明显变化。花后42 d,Ca+Mg处理的果肉可滴定酸含量显著低于其他处理;花后49 d,Ca处理显著最高;其余时间各处理间无显著差异。由此可见,各处理在试验前期果肉可滴定酸积累量均呈降低趋势,Ca处理积累量相对较高,在后期则各处理间的果肉可滴定酸含量无显著差异(P>0.05,下同),说明妃子笑荔枝果实成熟时果肉风味品质主要决定于果肉可溶性糖含量。
2. 2 妃子笑荔枝果肉不同途径呼吸速率变化
2. 2. 1 总呼吸速率 如图3所示,各处理的荔枝果肉总呼吸速率均在花后56 d之前上升,之后Ca处理和Ca+Mg处理一直下降,Mg处理和CK均在63 d上升至最高值后下降。花后35 d,各处理间差异不显著;花后42 d,Mg处理和Ca处理显著高于CK和Ca+Mg处理,前二者之间和后二者之间无显著差异;花后49 d,Mg处理显著最高,其余处理间无显著差异;花后63 d,各施肥处理的总呼吸速率排序依次为Mg>Ca>Ca+Mg,CK与Mg处理和Ca处理间差异不显著;花后70 d,Mg处理显著高于其余处理,Ca+Mg处理显著最低,Ca处理与CK间无显著差异。综上,Mg处理对总呼吸速率表现出一定的促进作用,而Ca+Mg处理呈现抑制作用。
2. 2. 2 MEP途径呼吸速率 如图4所示,各处理的荔枝果肉EMP途径呼吸速率动态变化趋势存在明显差异。其中,CK的果肉呼吸速率呈上升—下降—上升的变化趋势;Ca处理的果肉呼吸速率在前期上升,至花后49 d之后下降,后无明显变化;Mg处理和Ca+Mg处理的果肉呼吸速率上升至花后42 d之后下降,但前者至花后63 d上升,后者至花后49 d之后上升。花后49~56 d、花后70 d,CK均高于施肥处理;花后49 d,Mg处理与Ca处理间差异不显著,且显著高于Ca+Mg处理;花后56和70 d,Ca处理与Ca+Mg处理间差异不显著,且显著低于Mg处理;花后63 d,Ca处理和Ca+Mg处理显著低于CK和Mg处理,前两者和后两者之间无显著差异。综上,施钙镁肥处理对EMP途径的呼吸速率均表现出不同程度的抑制作用,且Ca+Mg处理抑制作用最明显,Ca处理的抑制作用次之。
2. 2. 3 TCA途径呼吸速率 如图5所示,各处理的荔枝果肉TCA途径呼吸速率动态变化特点不同。其中,CK的TCA途径呼吸速率呈现2次上升—下降的交替变化趋势,在花后70 d上升;Ca处理和Ca+Mg处理均呈现先上升后下降的变化趋势;Mg处理在前期上升,至花后56 d后小幅下降,至末期回升。花后42 d,4个处理按照CK、Ca+Mg处理、Mg处理、Ca处理依次递减;花后49 d以后,Ca处理显著高于其他处理;Ca+Mg处理的TCA途径呼吸速率在花后49 d和63 d呈现次高;花后49~56 d,Mg处理显著低于Ca+Mg处理,花后56 d显著最低;花后63 d,CK显著最低,且Mg处理与Ca+Mg处理间无显著差异;花后70 d,Ca+Mg处理的TCA途径呼吸速率较低,Ca处理和Mg处理显著高于Ca+Mg处理,CK与Ca+Mg处理间差异不显著。综上所述,不同处理对TCA途径呼吸速率的影响不同;Ca处理和Mg处理在前期对TCA途径呼吸速率表现出抑制作用,前者和后者分别在中后期和末期表现出一定的促进作用;在花后56 d前后,Ca+Mg处理对TCA途径呈现抑制作用。说明Ca+Mg处理在成熟期前后抑制TCA途径是其减缓退糖现象的原因之一,Ca处理和Mg处理仍然呈退糖现象原因之一可能是TCA途径在后期增强。
2. 3 妃子笑荔枝果肉呼吸作用相关酶活性变化
2. 3. 1 GPI活性 如图6所示,Ca+Mg处理的GPI活性呈现上升—下降—平缓趋势;Ca处理呈现上升—下降的交替变化趋势。其中,花后42 d,Ca+Mg处理与Ca处理差异不显著,Mg处理与CK差异不显著,前二者显著高于后二者;花后49 d,Ca+Mg处理显著高于CK和Ca处理,并与Mg处理差异不显著,Ca处理与CK差异不显著;花后56 d,Ca处理与CK差异不显著,Mg处理与Ca+Mg处理差异不显著,前二者显著高于后二者;花后63 d,施钙镁肥处理均显著低于CK,且3个处理间差异不显著;花后70 d,Mg处理显著最高,其余处理间差异不显著。综上所述,Ca处理和Ca+Mg处理在前期促进GPI活性,在后期Ca+Mg处理呈现抑制酶活性作用;Mg处理则在花后49和70 d起促进酶活性作用,在其他时间也抑制酶活性。说明Ca+Mg处理在完全成熟期抑制EMP途径是通过抑制GPI活性而达到。
2. 3. 2 PK活性 如图7所示,Mg处理、Ca+Mg处理和CK的荔枝果肉PK活性均呈现上升—下降的交替变化趋势,Ca处理在后期有所上升。其中,花后42 d,各处理按照Mg处理、Ca处理、Ca+Mg处理、CK依次递减;花后56 d,Ca处理的PK活性显著最高,其他处理相互间差异不显著;花后63 d,Ca处理的PK活性显著最低,其余处理间差异不显著;花后70 d,Ca+Mg处理的PK活性显著高于其他处理,其他处理相互间差异不显著。总之,花后49 d之前,施钙镁肥处理均促进了PK活性;Ca处理呈现在中期促进和在末期抑制PK活性的趋势,Ca+Mg处理在采收前后则有一定的促进作用。说明PK不是Ca+Mg处理抑制TCA途径的关键酶。
2. 3. 3 PDC活性 如图8所示,不同處理的荔枝果肉PDC活性动态变化趋势差异明显。其中,CK和Ca处理呈2次下降—上升的交替变化趋势;Ca+Mg处理在花后42 d后上升,至56 d后下降;Mg处理呈上升—下降—上升的变化趋势,且全程显著最高;花后42~63 d,Ca、Ca+Mg、CK相互间差异不显著,Mg处理显著最高;花后70 d,Ca处理和Mg处理显著高于CK和Ca+Mg处理,前二者和后二者分别差异不显著。综上,Mg处理对PDC活性呈促进作用,Ca处理只在末期对PDC活性具有促进作用,Ca+Mg处理未表现出明显抑制作用。说明PDC酶活性并非是是Ca+Mg处理抑制TCA途径的主要原因。
2. 3. 4 SDH活性 如图9所示,各处理的荔枝果肉SDH活性具有不同的动态变化趋势。其中,CK呈上升—下降—上升的趋势;Ca处理呈2次上升—下降的交替过程;Mg处理在前期无明显变化,后期呈上升趋势;Ca+Mg处理呈现上升—下降的趋势。花后42 d,Ca处理显著高于其他处理,其他处理间无显著差异;花后49 d,施钙镁肥处理均显著低于CK,且按照CK、Ca、Ca+Mg和Mg处理依次递减;花后56 d,Ca处理的酶活性显著最高,Mg处理显著最低,Ca+Mg处理和CK位于中间且差异不显著;花后63 d,Ca+Mg处理与Mg处理差异不显著,Ca处理与CK差异不显著,前二者显著高于后二者;花后70 d,Ca+Mg处理显著低于其他处理,其他处理相互间差异不显著。总之,Ca处理在花后35~56 d呈促进酶活趋势;Mg处理在中期抑制酶活性,自花后63 d表现出促进作用;Ca+Mg处理在采收期活性降低则可能是Ca+Mg处理抑制TCA途径的原因之一。
2. 3. 5 NAD-MDH活性 如图10所示,各处理的荔枝果肉NAD-MDH活性动态变化趋势存在明显差异。其中,CK和Ca+Mg处理呈2次下降—上升交替过程;Ca处理呈近似L形的下降趋势;Mg处理呈下降—上升—下降的趋势。花后42 d,Ca处理的果肉NAD-MDH活性显著最高,Ca处理和Mg处理显著高于CK和Ca+Mg处理,后二者间差异不显著;花后49 d,Ca处理的果肉NAD-MDH活性显著最低,其余2个施肥处理与CK间差异不显著;花后56 d,Mg处理和Ca+Mg处理显著高于CK和Ca处理,前二者和后二者间分别差异不显著;花后63 d,CK的果肉NAD-MDH活性显著最低,3个施肥处理间差异不显著;花后70 d,Ca+Mg处理显著高于其他处理,CK又显著高于Ca处理和Mg处理,Ca处理与Mg处理间则差异不显著。综上所述,Ca处理在前期对NAD-MDH酶活性有促进作用,后期呈现抑制作用;Ca+Mg处理在中、后期呈现出促进酶活性的作用。说明NAD-MDH在Ca+Mg处理抑制TCA途径中不是关键酶。
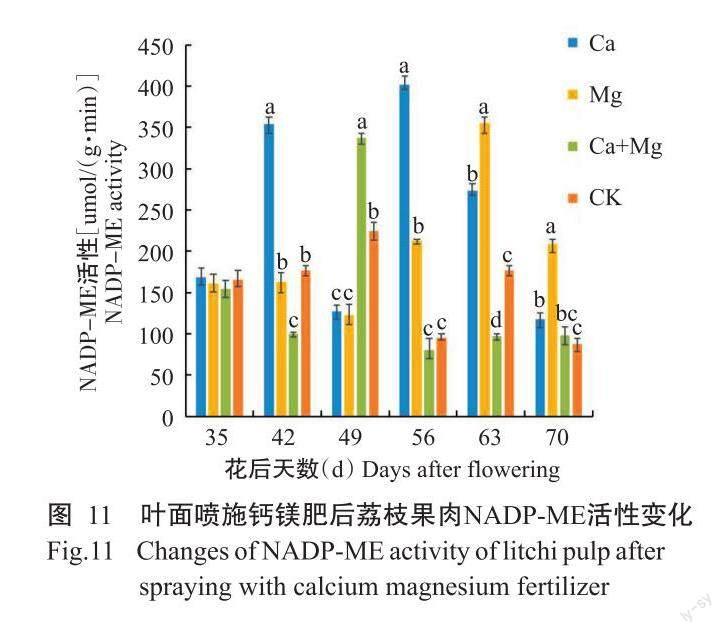
2. 3. 6 NADP-ME活性 如图11所示,各处理的荔枝果肉NADP-ME活性动态变化趋势明显不同。其中,Ca处理和CK呈2次上升—下降的交替变化趋势;Mg处理前期呈下降趋势,至花后56 d上升,在末期下降。花后42 d,Ca处理的果肉NADP-ME活性显著最高,Ca+Mg处理显著低于其他处理,Mg处理与CK差异不显著;花后49 d,Ca+Mg处理显著最高,CK次之,Ca处理与Mg处理差异不显著;花后56 d,Ca处理显著最高,Mg处理次之,Ca+Mg处理和CK最低且两者间差异不显著;花后63 d,各处理间差异显著,且按照Mg处理、Ca处理、CK和Ca+Mg处理的顺序递减;花后70 d,Mg处理显著最高,Ca+Mg处理与Ca处理和CK差异不显著,Ca处理与CK差异显著。总之,Mg处理前期抑制但后期促进酶活性,Ca处理整体促进酶活,Ca+Mg处理在56~63 d抑制酶活。说明Ca+Mg处理在采收期前抑制NADP-ME活性是其抑制TCA途径呼吸速率原因之一。
3 讨论
本研究结果表明,Ca+Mg处理减缓了妃子笑荔枝果肉的退糖现象,而Ca处理和Mg处理则与CK一样仍然出现退糖现象,该结果与本课题组之前在不同试验点和不同年份所取得的试验结果(张锐等,2014;周晓超等,2015;王展等,2017)一致。说明叶面喷施钙镁混合肥可减缓妃子笑荔枝果肉退糖现象,因而可作为改良的施肥技术在生产实践中加以应用。
植物的呼吸代谢途径有多种方式,EMP作为完全有氧呼吸的第一步,是将葡萄糖和糖原降解为丙酮酸并伴随ATP产生的一系列反应(何亚辉,2007),呼吸作用消耗糖分的多少主要取决于EMP呼吸速率的高低。本研究结果表明Ca+Mg处理和Ca处理在果实采前均抑制EMP呼吸速率,这为二者减缓果肉退糖现象出现提供了可能性。GPI催化6-磷酸葡萄糖的异构化反应,催化其转变为6-磷酸果糖,是EMP途径的关键酶,PK催化磷酸烯醇式丙酮酸转化为丙酮酸,是EMP途径的最后一步不可逆反应,这2种酶普遍被认为是EMP途径的限速酶(李立梅等,2018),植物呼吸中的EMP途径主要受GPI和NaF的调节(郑集和陈钧辉,2003)。本研究结果表明,在果实成熟前后,Ca+Mg处理抑制了荔枝果肉GPI活性,Ca处理次之,从而导致二者在花后63 d之后EMP途径呼吸速率降低。在EMP酶系中,PK通过将ADP和PEP转化为ATP和丙酮酸以催化糖酵解途径的末端反应,在糖酵解的调节中起着至关重要的作用(宋丛丛,2021)。同时,Ca处理具有在中期促进和后期抑制PK酶活性的趋势,Ca+Mg处理在末期则呈现一定的促进作用,说明影响EMP途径的最关键限速酶是GPI。综上所述,Ca+Mg处理和Ca处理通过在果实采前抑制GPI活性而抑制EMP呼吸速率。
呼吸作用的第二大途径就是TCA途径,是需氧生物体内普遍存在的呼吸代谢途径,在有氧条件下,利用糖酵解的终产物丙酮酸,在丙酮酸脱氢酶酶系的催化下氧化脱羧,进行彻底的氧化分解。该途径是三大营养素(糖类、脂质、氨基酸)的最终代谢通路和共同氧化途径,在上述过程中产生的部分中间产物,是合成其他许多重要物质的前体,并起着与其他代谢途径相互联系、相互转变的纽带作用(李冬彪,2017)。TCA途径的第1个酶是线粒体PDC,其将丙酮酸转化为乙酰辅酶A。SDH是参与TCA呼吸通路和线粒体电子传输链的唯一酶,先后催化琥珀酸氧化为富马酸,将泛醌(Q)还原为泛醇(QH2)(Chumyam et al., 2016)。NAD-MDH也是TCA中的关键酶,NAD-MDH催化苹果酸仲醇基脱氢氧化成羰基,生成草酰乙酸。本研究结果表明,Ca+Mg处理在采收期抑制SDH和NADP-ME活性;Ca处理和Mg处理则在末期促进PDC和NADP-ME活性,抑制NAD-MDH酶活性,说明本文影响TCA途径最关键的限速酶可能是SDH和NADP-ME。Ca+Mg处理通过抑制SDH和NADP-ME等活性而抑制TCA途径,引起该途径呼吸速率降低,这与前人的研究结果(李霞,2004;李宏博,2005)基本一致。相关研究也表明,NO与SDH发生作用,通过抑制琥珀硫激酶和琥珀酸脱氢酶活性控制TCA途径(马春媚,2013)。有研究表明,PDC活性会直接影响进入TCA途径物质的多少,进而控制CO2释放量,最终影响呼吸速率(李熙萌等,2016)。Ca处理可能通过增强PDC活性而促进TCA途径,存在促使呼吸速率升高的现象。综上,Ca+Mg处理通过抑制果肉SDH和NADP-ME酶活性而抑制果肉TCA途径呼吸速率,Ca处理通过在果肉成熟期促进SDH和NADP-ME酶活性,并在采收期促进PDC活性进而提高果肉呼吸速率,使得该途径呼吸速率显著高于Ca+Mg处理。
由于植物呼吸作用主要以EMP-TCA途径为主,其决定总呼吸速率的高低(张强,2020)。基于上述讨论可知,Ca+Mg处理通过抑制果肉GPI及SDH和NADP-ME等活性,分别抑制果肉EMP和TCA途径,进而抑制EMP-TCA途径完全有氧呼吸作用,因此出现本研究中Ca+Mg处理果肉总呼吸速率降低的结果,因而减少了果肉糖分的损耗而减缓果肉退糖现象;Ca处理虽然在果实采前抑制EMP,但其同时促进了TCA途径,因而与Mg处理和CK均未降低总呼吸速率,从而引起果肉退糖现象。
4 结论
妃子笑荔枝叶面喷施0.3%的钙镁混合肥可减缓其果肉退糖现象,原理是Ca+Mg混合叶面处理通过抑制果肉主要呼吸途径的呼吸速率进而减少果肉糖分消耗,从而减缓退糖现象。生产中钙镁混合肥可作为妃子笑荔枝果实品质发育调节的改良施肥技术加以采用。
参考文献:
陳宇新. 2017. 油菜素内酯延缓杨桃品质劣变的效果及其对呼吸代谢和内源激素变化的影响[D]. 广州:华南农业大学. [Chen Y X. 2017. Exogenous application of BRs to delay carambola fruit quality deterioration and its effects on respiratory metabolism and endogenous hormone changes[J]. Guangzhou: South China Agricultural University.]
何亚辉. 2007. 丙酮酸脱氢酶系测活方法的比较研究与创新[D]. 武汉:华中师范大学. [He Y H. 2007. Comparision and development of measuring methods for Pyruvate Dehydrogenase complex[D]. Wuhan:Central China Normal University.]
李冬彪. 2017. 网纹甜瓜幼苗光合作用和根呼吸代谢对根际高CO2胁迫后恢复的响应[D]. 沈阳:沈阳农业大学. [Li D B. 2017. Responses of photosynthesis and root respiratory metabolism of netted muskmelon to root-zone high CO2 concentration restoration after stress[D]. Shenyang: Shenyang Agricultural University.]
李宏博. 2005. GA对变温层积条件下解除辽东楤木种子休眠的作用及其机制研究[D]. 沈阳:沈阳农业大学. [Li H B. 2005. The research on the effect of GA on breaking dormancy of Aralia elate seed and the mechanism of that under altering temperature stratification[D]. Shenyang: Shenyang Agricultural University.]
李立梅,李鑫,赵秀香,毛赫,孙景花,吴元华. 2018. 西瓜感染黄瓜绿斑驳花叶病毒后糖分代谢与倒瓤的关系[J]. 植物保护学报,45(3):447-454. [Li L M, Li X, Zhao X X, Mao H, Sun J H, Wu Y H. 2018. The relationships between sugar metabolism and blood-flesh of watermelon infected with cucumber green mottle mosaic virus[J]. Journal of Plant Protection, 45(3):447-454.]doi:10. 13802/j.cnki.zwbhxb.2018.2016208.
李熙萌,杨琼,李征珍,刘海鸥,冯金朝. 2016. 胡杨(Populus euphratica)叶片呼吸作用对地下水埋深的响应[J]. 生态科学,35(3):29-36. [Li X M, Yang Q, Li Z Z, Liu H O, Fen J Z. 2016. Response of foliage respiratory cha-racteristics of Populus euphratica to underground water table[J]. Ecological Sciene,35(3):29-36.] doi:10.14108/j.cnki.1008-8873.2016.03.005.
李霞. 2004. 呼吸代谢对甜樱桃自然休眠的调控及破眠技术研究[D]. 泰安:山东农业大学. [Li X. 2004. Control of respiratory metabolism on sweat cherry dormancy and studies on technical of dormancy release[D]. Taian: Shan-dong Agricultural University.]
廖海枝,林晓凯,杨成坤,杜婧加,彭俊杰,周开兵. 2021. 钙镁叶面肥对‘妃子笑荔枝果肉柠檬酸积累的影响[J]. 热带作物学报,42(12):3537-3544. [Liao H Z,Lin X K,Yang C K,Du J J,Peng J J,Zhou K B. 2021. Effects of foliar calcium and magnesium fertilizer on citric acid accumulation of ‘Feizixiao litchi fruit[J]. Chinese Journal of Tropical Crops,42(12):3537-3544.] doi:10.3969/j.issn.1000-2561.2021.12.021.
羅安才,杨晓红,邓英毅,李纯凡,向可术,李道高. 2003. 柑橘果实发育过程中有机酸含量及相关代谢酶活性的变化[J]. 中国农业科学,36(8):941-944. [Luo A C, Yang X H, Deng Y Y, Li C F, Xiang K S, Li D G. 2003. Organic acid concentrations and the relative enzymatic changes during the development of citrus fruits[J]. Scientia Agricultura Sinica,36(8):941-944.]
吕润,梁彩红,邹海平,李伟光,白蕤. 2021. 海南妃子笑荔枝精细化农业气候区划研究[J]. 热带农业科学,41(12):117-122. [Lü R, Liang C H, Zou H P, Li W G, Bai R. 2021. Precise agricultural climate regional planning of litchi Feizixiao in Hainan[J]. Chinese Journal of Tropical Agriculture, 41(12):117-122.] doi:10.12008/j.issn.1009-2196.2021.12.018.
马春媚. 2013. 一氧化氮对桃果实三羧酸循环相关酶活性及蛋白质结构的影响[D]. 泰安:山东农业大学. [Ma C M. 2013. Effect of nitric oxide on activities and structure of enzymes in tricarboxylic acid cycle[D]. Taian:Shandong Agricultural University.]
倪耀媛,吴素芬. 1990. 荔枝栽培[M]. 北京:农业出版社:8-9. [Ni Y Y, Wu S F.1990. Litchi cultivation[M]. Beijing: China Agriculture Press: 8-9.]
史青纯. 2009. 外源水杨酸处理对采后黄冠梨果实呼吸途径的影响[D]. 保定: 河北农业大学. [Shi Q C. 2009. Effects of salicylic acid on respiration pathway of postharvest Huang-guan pear(Pyrus bretschneideri Rehd.)[D].Baoding:Agricultural University of Hebei.]
宋丛丛. 2021. NO处理对不同肉质桃果实冷藏过程中呼吸作用的影响[D]. 北京:中国农业科学院. [Song C C. 2021. Effects of NO treatment on the respiration of different fleshy peach fruit during cold storage[D]. Beijing: Chinese Academy of Agriculture Scienses.] doi:10.27630/d.cnki.gznky.2021.000557.
苏阳,周晓超,高丹,周开兵. 2015. ‘妃子笑荔枝果肉中主要风味物质与钾钙镁含量的关系[J]. 热带作物学报,36(6):1131-1135. [Su Y,Zhou X C,Gao D,Zhou K B. 2015. Studies on the relationship between the main flavor components and the contents of K,Ca and Mg in flesh of Feizixiao litchi(Litchi chinensis Sonn. cv Feizi-xiao)[J]. Chinese Journal of Tropical Crops,36(6):1131-1135.] doi:10.3969/j.issn.1000-2561.2015.06.019.
王展,李世军,袁孟玲,周开兵. 2017. 叶面喷施镁肥对妃子笑荔枝果皮着色及钾钙镁含量的影响[J]. 南方农业学报, 48(5):854-860. [Wang Z, Li S J, Yuan M L, Zhou K B. 2017. Effects of magnesium foliar application on pericarp coloring of Litchi chinensis Sonn. cv. Feizixiao and contents of potassium, calcium and magnesium in pericarp[J]. Journal of Southern Agriculture,48(5):854-860.] doi:10.3969/j.issn.2095-1191.2017.05.016.
翁雪成,袁红. 2007. 蒽酮—硫酸法测定荔枝核中可溶性多糖含量[J]. 杭州师范学院学报(医学版),27(2):107-108. [Weng X C, Yuan H. 2007. Determination of soluble polysaccharide content in litchi kernel by anthrone-sulfuric acid method[J]. Journal of Hangzhou Teachers College (Medical Edition),27(2):107-108.]
姚智,李华东,白亭玉,康专苗,王艺蓉,仇海威,林电. 2017. 叶施钙肥对芒果果实呼吸、乙烯释放及矿质元素含量的影响[J]. 中国土壤与肥料,(2):95-101. [Yao Z, Li H D, Bai T Y, Kang Z M, Wang Y R, Qiu H W, Lin D. 2017. Effect of foliar application of calcium fertilizer on respiration,ethylene release and mineral contents of mango fruit[J]. Soil and Fertilizer Sciences in China,(2):95-101.] doi:10. 11838/sfsc.20170215.
于超. 2021. 呼吸代謝对甜菜块根生长的作用机理研究[D]. 呼和浩特:内蒙古农业大学. [Yu C. 2021. Effects me-chanism of respiratory metabolism on taproot growth in sugar beet(Beta vulgaris L.)[D]. Hohhot: Inner Mongolia Agricultural University.] doi:10.27229/d.cnki.gnmnu. 2021.000023.
张强. 2020. 钙与1-MCP调控甜瓜后熟软化机理及近冰温贮藏技术研究[D]. 乌鲁木齐:新疆大学. [Zhang Q. 2020. Mechanism of calcium and 1-MCP regulating post-ripe-ning softening and near freezing temperature storage technology of melon[D]. Urumqi: Xinjiang University.] doi:10.27429/d.cnki.gxjdu.2020.001434.
张锐,韩丽婷,王娟,周开兵. 2014. 两个荔枝品种果皮着色和果肉糖酸积累特点的比较[J]. 福建农林大学学报(自然科学版),43(4):374-378. [Zhang R, Han L T,Wang J,Zhou K B. 2014.The comparison of the differences in pericarp colouring and accumulations of sugar and acid in flesh between two litich cultivars[J]. Journal of Fujian Agriculture and Forestry University(Natural Science Edition),43(4):374-378.] doi:10.13323/j.cnki.j.fafu(nat.sci.).2014.04.010.
鄭集,陈钧辉. 2003. 普通生物化学[M]. 第3版. 北京: 高等教育出版社. [Zheng J, Chen J H. 2003. General biochemistry[M]. The 3rd Edition. Beijing: Higher Education Press.]
周晓超,苏阳,张锐,周开兵. 2015. 叶面喷布钾、钙和镁肥对妃子笑荔枝果皮着色的调节效果[J]. 西南农业学报,28(4):1713-1718. [Zhou X C,Su Y,Zhang R,Zhou K B. 2015. Effects of K,Ca and Mg applied in foliar nutrients on pericarps coloring of Litchi chinensis Sonn. cv. Feizi-xiao[J]. Southwest China Journal of Agriculture Sciences,28(4):1713-1718.] doi:10.16213/j.cnki.scjas.2015.04.056.
Analin B, Mohanan A, Bakka K, Challabathula D. 2020. Cytochrome oxidase and alternative oxidase pathways of mitochondrial electron transport chain are important for the photosynthetic performance of pea plants under salinity stress conditions[J]. Plant Physiology and Biochemistry,154:248-259. doi:10.1016/j.plaphy.2020.05.022.
Chumyam A, Shank L, Uthaibutra J, Saengnil K. 2016. Effects of chlorine dioxide on mitochondrial energy level and redox status of ‘Daw longan pericarp during storage[J]. Postharvest Biology and Technology,116:26-35. doi:10.1016/j.postharvbio.2016.01.002.
Sadka A, Dahan E, Cohen L, Marsh K B. 2000. Aconitase activity and expression during the development of lemon fruit[J]. Physiologia Plantarum,108(3):255-262. doi:10.1034/j.1399-3054.2000.108003255.x.
Sivakumar D, Korsten L. 2010. Fruit quality and physiological responses of litchi cultivar McLeans Red to 1-methylcyclopropene pre-treatment and controlled atmosphere storage conditions[J]. LWT-Food Science and Technology,43(6):942-948. doi:10.1016/j.lwt.2010.02.001.
Vanlerberghe G C,Dahal K,Alber N A,Chadee A. 2020. Photosynthesis,respiration and growth:A carbon andenergy balancing act for alternative oxidase[J]. Mitochond-rion, 52:197-211. doi:10.1016/j.mito.2020.04.001.
Wang H C, Huang H B, Huang X M,Hu Z Q. 2006. Sugar and acid compositions in the arils of Litchi chinensis Sonn.:Cultivar differences and evidence for the absence of succinic acid[J]. Journal of Horticultural Science & Biotechnology,81(1):57-62. doi:10.1080/14620316.2006. 11512029.
Wang Y F, Wang P, Xia J D, Yu T, Lou B G, Wang J,Zheng X D. 2010. Effect of water activity on stress tole-rance and biocontrol activity in antagonistic yeast Rhodosporidium paludigenum[J]. International Journal of Food Microbiology, 143(3): 103-108. doi:10.1016/j.ijfoodmicro.2010.07.035.
Zhang Y H, Wang Z M, Huang Q,Shu W. 2008. Phosphoenolpyruvate carboxylase activity in ear organs is related to protein concentration in grains of winter wheat[J]. Journal of Cereal Science,47(12):386-391. doi:10.1016/j.jcs.2007.04.011.
(责任编辑 邓慧灵)
