不同品种石榴枝条的抗寒性评价
2023-07-18雷梦瑶高小峰白清敏左卫芳李玉英
雷梦瑶,高小峰,白清敏,邓 珂,左卫芳,李玉英
(1.南阳师范学院 河南省软籽石榴工程研究中心,河南省南水北调中线水源区流域生态安全国际联合实验室,河南 南阳 473061;2.南阳市农业科学院,河南 南阳 473000;3.南阳市农村能源环境保护管理站,河南 南阳473000;4.南阳农业职业学院 农业工程学院,河南 南阳 473000)
石 榴(Punica granatumL.)属 于 石 榴 科(Punicaceae)石榴属(Punica)落叶灌木或乔木[1]。根据籽粒软硬程度分为硬籽石榴和软籽石榴,其中软籽石榴因引自热带地区,耐寒性较差,在低温下生长发育缓慢,开花结果能力下降,制约着其在生产中的推广和应用[2-3]。河南省南阳市地处亚热带季风气候和温带季风气候过渡带,气候独特,是石榴的适宜种植区[4]。南阳市淅川县作为南水北调中线核心水源区,为了保障水质安全,建设高效生态产业带,截至2020 年,软籽石榴种植面积达到3 333 hm2,但个别年份受极端严寒天气影响,部分地方栽植的软籽石榴会遭受较为严重的冻害。冻害是限制软籽石榴生长和地理分布的重要环境因子之一[5],严重的冻害使整个植株、枝条和花芽受冻导致死亡,给广大果农造成严重的经济损失,因此,筛选本土化耐寒石榴品种意义重大。
作为广泛栽培的果树种类,石榴抗寒生理特性研究一直被科研人员所关注。罗华等[6]对山东省枣庄市峄城区农家品种与6个新品种石榴的抗寒性进行了综合评价;毕润霞等[7]利用电导法对石榴抗寒性进行了评价;姚方等[8]分析了以色列软籽品种等8个石榴品种枝条在4 ℃到-15 ℃的低温下冷冻24 h后生理指标的变化,并进行抗寒性排序;王新宇[9]研究了不同胁迫时间和低温对突尼斯软籽石榴生理指标的影响;王庆军等[10-11]分析了山东省枣庄市峄城区3 个石榴品种在不同低温下的半致死温度(LT50)和抗寒性排序,接着对24 个原产于不同省份的不同石榴品种进行低温处理,并结合隶属函数法对其抗寒性进行等级划分;张艳侠等[12]探究了山东、新疆等地5 个石榴品种枝条在-20~-8 ℃低温下相对电导率的变化及其抗寒能力。
淅川县石榴栽培面积大,种类多,但是关于主栽品种抗寒特性的报道还比较少,果农们在引进新的石榴品种时无从参考。石榴越冬时,枝条何时受到冻害威胁,又如何通过生理变化来适应低温变化,是需要关注的问题。鉴于此,以淅川县4个石榴品种1年生休眠枝条为试材,通过模拟低温环境,分析不同温度下相对电导率的变化情况,配合Logistic方程计算LT50,并通过分析石榴在低温胁迫下丙二醛(MDA)含量,超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)活性等8 个生理指标的变化,结合隶属函数法来综合评价其抗寒性,为抗寒石榴品种的选育与栽培提供理论依据。
1 材料和方法
1.1 材料采集
供试材料取自河南省淅川县仁和康源软籽石榴基地和豫淅红软籽石榴基地。2022 年3 月中旬,对生长良好、无病害的突尼斯、中农红、红双喜和千层花1 年生健康枝条进行采集,枝条直径为0.7~0.8 cm,截取长度均为15 cm,用湿润纱布包好放置在塑料袋中,迅速带回实验室备用。
1.2 材料处理
将枝条依次用自来水、蒸馏水冲洗3遍后,用吸水纸擦干,石蜡封闭枝条两端,每种枝条分成7 组,每组10条,用保鲜膜包裹做好标记后将枝条分别置于4 ℃、0 ℃、-3 ℃、-6 ℃、-9 ℃、-12 ℃和-15 ℃低温下进行冷冻处理,冷冻时温度下降幅度为2 ℃/h,到达目标温度后持续24 h,然后解冻至室温放置3 h,进行相关指标的测定。
1.3 测定指标及方法
1.3.1 相对电导率 把低温处理后的枝条剪成0.5 cm 长的小段,量取1 g,放入50 mL 的试管中,加入去离子水25 mL,于25 ℃摇床上摇90 min。然后用电导率仪测其初电导率值,随后置于沸水浴中20 min,取出恢复室温后测定其终电导率值,计算相对电导率(REC),REC=初电导率值/终电导率值×100%[13-14]。
LT50的测定:将不同温度处理下的相对电导率和处理温度之间的关系用Logistic方程进行拟合,通过计算拐点温度来确定LT50[15]。
1.3.2 其他指标 采用酸性茚三酮显色法测定脯氨酸(Pro)含量[16],蒽酮比色法测定枝条内可溶性糖(SS)含量[17],考马斯亮蓝法测定可溶性蛋白(SP)含量[18],MDA 含量、POD 活性、CAT 活性和SOD 活性指标的测定分别采用MDA 植物测定试剂盒、POD 测定试剂盒、CAT 测定试剂盒和SOD 测定试剂盒(均购自南京建成生物工程研究所)。
1.4 数据处理
用SPSS 19.0 和Excel 软件对试验数据进行误差、显著性分析和Logistic 方程回归分析[19]。用隶属函数法对4 个石榴品种的抗寒性进行综合评价,运用公式(1)(2)求出综合隶属度,综合隶属度越大说明抗寒能力越强,反之抗寒能力越弱[20]。隶属函数计算公式:
式中,f(xij)指的是i树种的j项指标的隶属函数值,Xij为指标的测定值,Xjmax和Xjmin分别为j项指标的最大值和最小值。与抗寒性呈正相关的SOD、POD、CAT 活性和脯氨酸、可溶性糖、可溶性蛋白含量用公式(2)计算,与抗寒性呈负相关的MDA 含量和相对电导率用公式(1)计算[21]。
2 结果与分析
2.1 低温胁迫对石榴生理生化指标的影响
2.1.1 相对电导率 4 个石榴品种的枝条经低温处理后,其相对电导率随低温胁迫的加剧呈“S”形增长趋势(图1)。4~-6 ℃时4 个品种的相对电导率整体缓慢上升,在-6 ℃时,千层花最低,突尼斯最高。-6 ℃降到-9 ℃时各品种相对电导率急剧增加,说明此温度范围内细胞膜受损程度较大,其中千层花增幅最大,为104.3%,红双喜增幅最小,为55.3%。且在-9 ℃时,突尼斯相对电导率最高,红双喜最低。-9~-15 ℃时,除红双喜外其余3 个品种的相对电导率趋于平稳甚至有所下降,推测-9 ℃以下细胞膜已被严重破坏。

图1 低温胁迫下4个石榴品种枝条相对电导率的变化Fig.1 Changes in relative electrical conductivity in branches of four pomegranate varieties under low temperature stress
2.1.2 POD 活性 如图2 所示,随着温度的降低,4个石榴品种的枝条POD 活性整体呈上升趋势。其中,红双喜在0~-3 ℃和-9~-12 ℃时的POD 活性均急剧上升,且除-9 ℃之外的其余温度下,红双喜的POD 活性均显著高于其他3 个品种。在4~-15 ℃时突尼斯和中农红的POD 活性整体上升幅度较大,-15 ℃时突尼斯显著高于中农红。
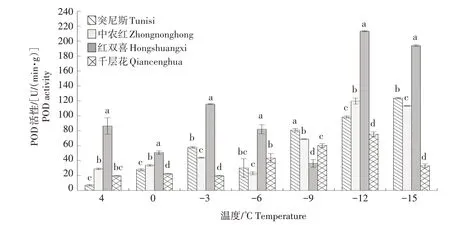
图2 低温胁迫下4个石榴品种枝条POD活性的变化Fig.2 Changes in POD activity in branches of four pomegranate varieties under low temperature stress
2.1.3 脯氨酸含量 如图3所示,随着温度的降低,4个石榴品种枝条的脯氨酸含量整体呈先下降后上升的趋势。在4~0 ℃各品种脯氨酸含量均有所下降,0~-3 ℃时又急剧上升,-3 ℃时突尼斯达到峰值,且高于其他3个品种。4~-12 ℃时,红双喜和千层花的脯氨酸含量均有所上升,在-12 ℃时达到峰值,其中,红双喜略高于千层花。
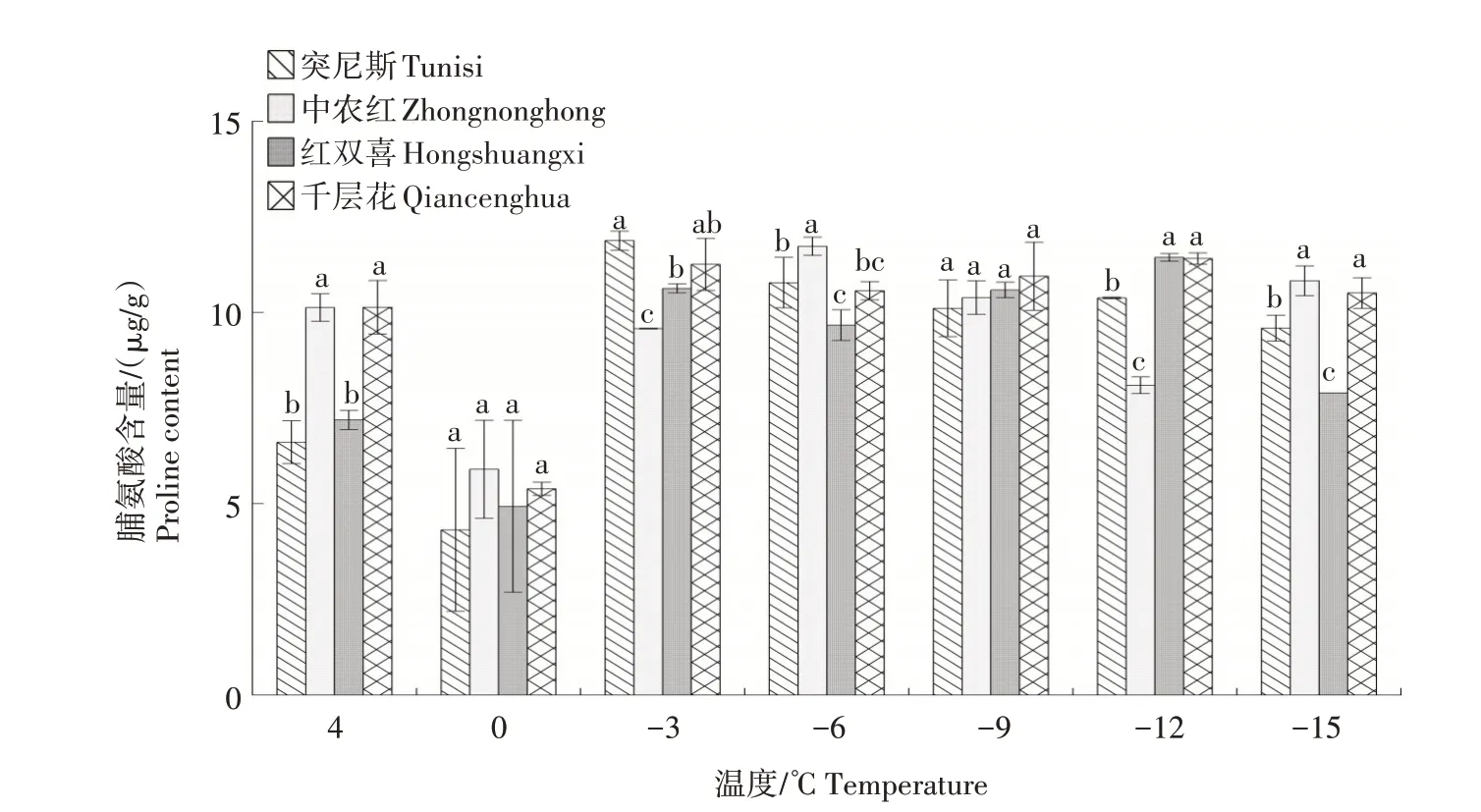
图3 低温胁迫下4个石榴品种枝条脯氨酸含量的变化Fig.3 Changes in proline content in branches of four pomegranates varieties under low temperature stress
2.1.4 可溶性蛋白含量 如图4 所示,随着温度的降低,4个品种的可溶性蛋白含量波动幅度较小,整体维持在0.15~0.25 mg/g。其中,千层花的可溶性蛋白含量在-6~-9 ℃时急剧上升,增幅为27.5%,中农红在-12~-15 ℃时增长幅度较大,为33.1%。突尼斯、中农红和千层花可溶性蛋白含量峰值分别出现在-15 ℃、-6 ℃和-12 ℃。
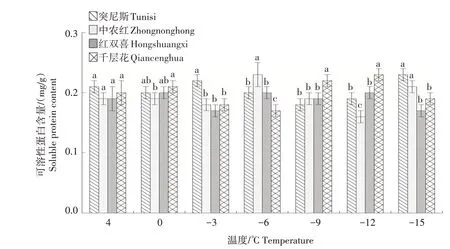
图4 低温胁迫下4个石榴品种枝条可溶性蛋白含量的变化Fig.4 Changes in soluble protein content in branches of four pomegranate varieties under low temperature stress
2.1.5 可溶性糖含量 如图5 所示,随着温度的降低,4 个石榴品种枝条的可溶性糖含量整体呈现上升趋势。在-3~-6 ℃时,各品种可溶性糖含量急剧上升,其中突尼斯和中农红增幅较大。-6 ℃时,突尼斯的可溶性糖含量达到峰值,但略低于中农红。-6~-9 ℃时突尼斯、中农红和千层花的可溶性糖含量均有所下降,而红双喜持续上升并在-9 ℃达到峰值,显著高于其他品种。-9~-15 ℃时中农红和千层花的可溶性糖含量逐渐上升,在-15 ℃达到峰值。
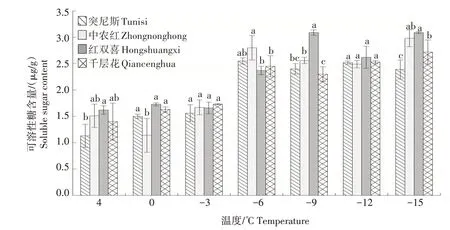
图5 低温胁迫下4个石榴品种枝条可溶性糖含量的变化Fig.5 Changes in soluble sugar content in branches of four pomegranate varieties under low temperature stress
2.1.6 SOD 活性 由图6 可以看出,随着温度的降低,4 个石榴品种枝条的SOD 活性整体波动较大。当温度在4~-3 ℃时,4 个品种的SOD 活性均呈现先降低后升高的变化趋势。在-3~-9 ℃时,红双喜、千层花的SOD 活性先降低后升高,与突尼斯、中农红的变化趋势相反,-9 ℃时,千层花SOD 活性最高。-9~-12 ℃时,4 个品种的SOD 活性逐渐降低,-12~-15 ℃时,红双喜持续降低。
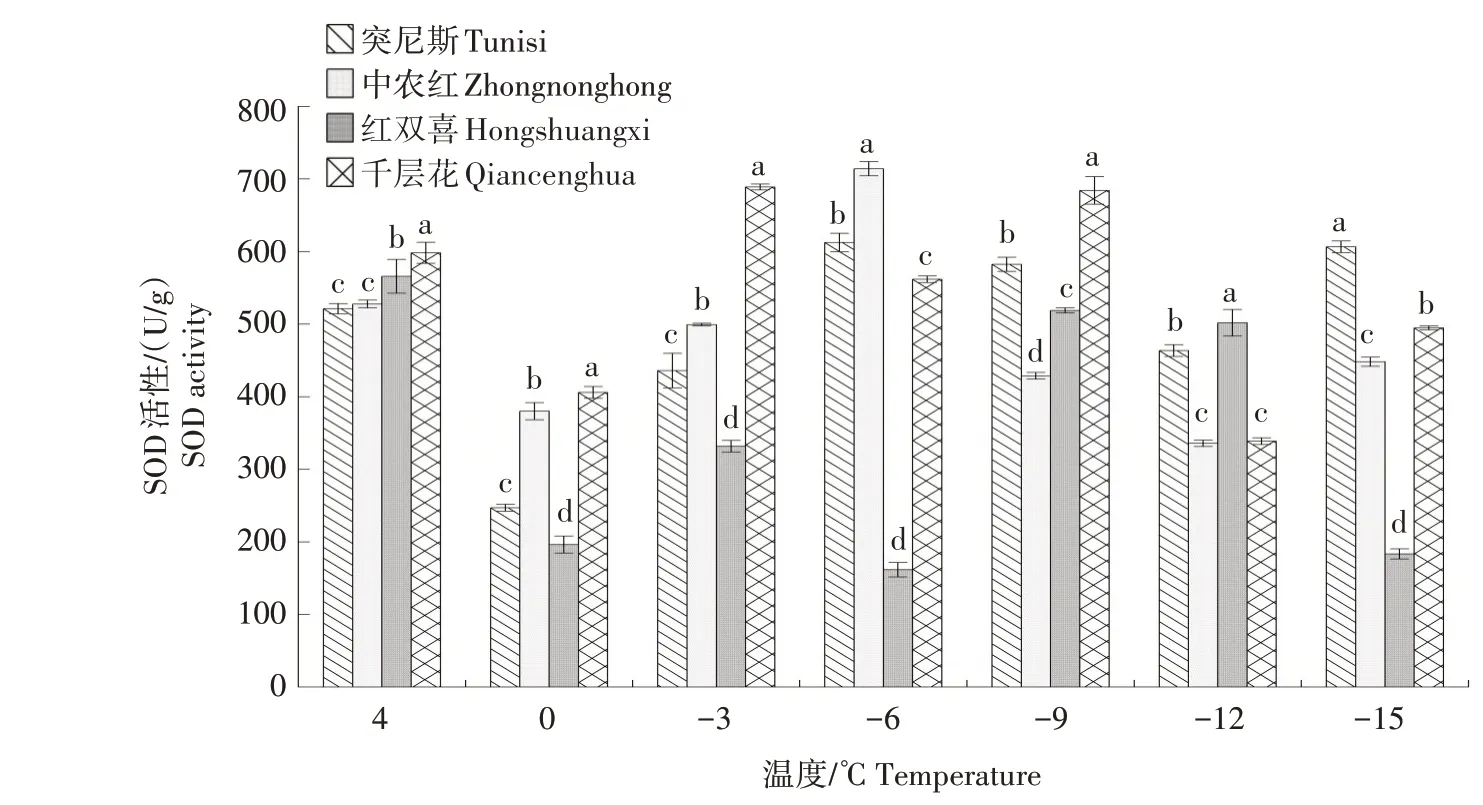
图6 低温胁迫下4 个石榴品种枝条SOD活性的变化Fig.6 Changes in SOD activity in branches of four pomegranate varieties under low temperature stress
2.1.7 MDA 含量 如图7 所示,随着温度的降低,除突尼斯外,其他3 个品种枝条的MDA 含量整体呈先上升后下降的趋势。在4~-6 ℃时,各品种的MDA 含量均有所增加。-6~-12 ℃时千层花的MDA含量逐渐下降,且始终低于其他3 个品种。红双喜的MDA 含量在-9 ℃时达到峰值,-9~-12 ℃时逐渐下降。突尼斯和中农红的MDA 含量变化幅度较为相似且幅度较小。
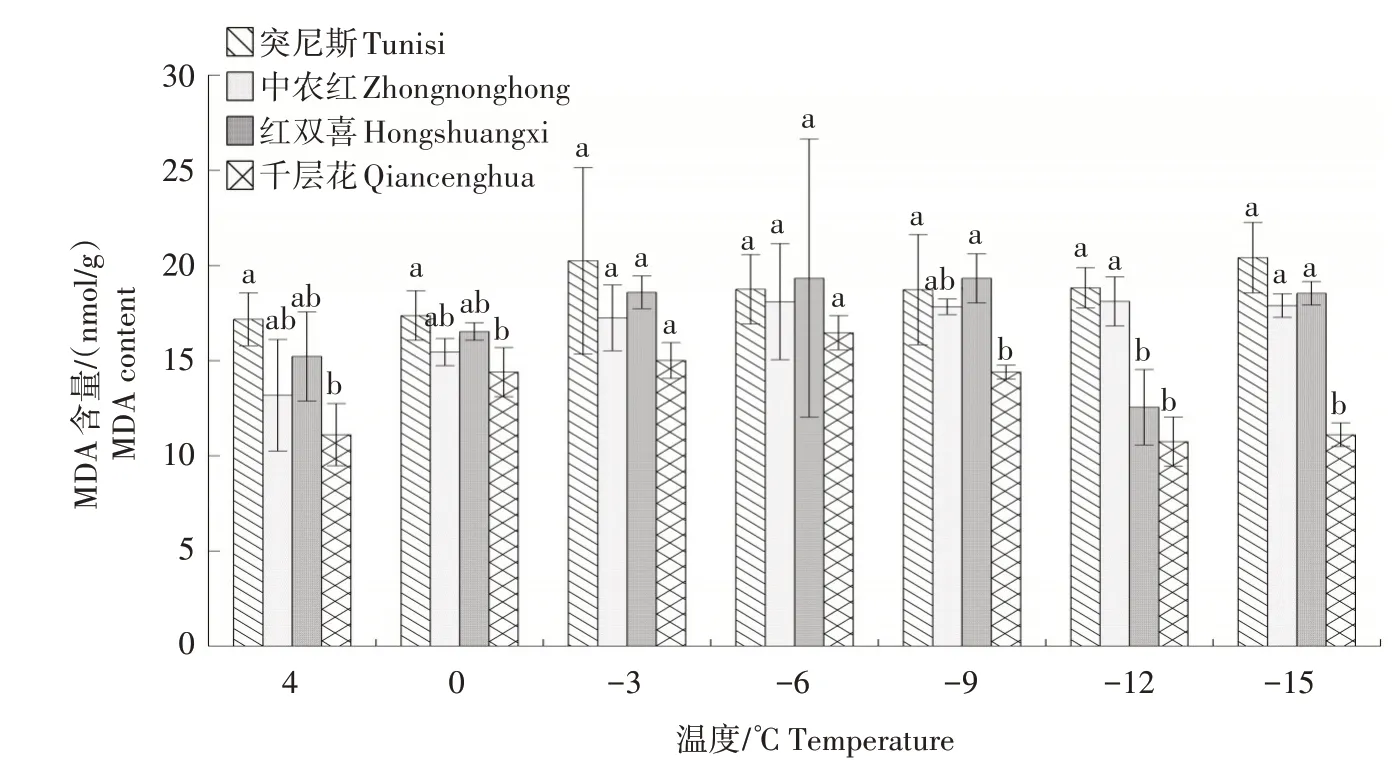
图7 低温胁迫下4个石榴品种枝条MDA含量的变化Fig.7 Changes in MDA content in branches of four pomegranate varieties under low temperature stress
2.1.8 CAT 活性 由图8 可知,随着温度的降低,4个品种枝条的CAT 活性整体呈先升高后降低的趋势。千层花的CAT 活性在4~-6 ℃时持续升高,并在-6 ℃时达到峰值,显著高于其他品种,随后出现降低趋势。突尼斯、中农红和红双喜的CAT 活性在4~-12 ℃时有所升高,-12 ℃时达到峰值,其中突尼斯和红双喜的CAT活性均显著高于中农红。
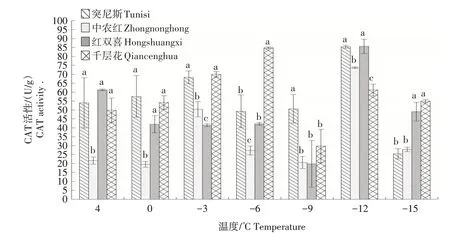
图8 低温胁迫下4个石榴品种枝条CAT活性的变化Fig.8 Changes in CAT activity of four pomegranate varieties under low temperature stress
2.2 低温胁迫下4个石榴品种的LT50
通过计算Logistic曲线拐点温度,比较各品种的LT50,在一定程度上鉴定不同石榴品种抗寒性强弱[22]。4 个石榴品种的LT50如表1 所示,千层花抗寒性最强,其次是中农红,温度分别比红双喜低4.67 ℃和2.62 ℃。4 个石榴品种抗寒性强弱依次为千层花>中农红>突尼斯>红双喜。
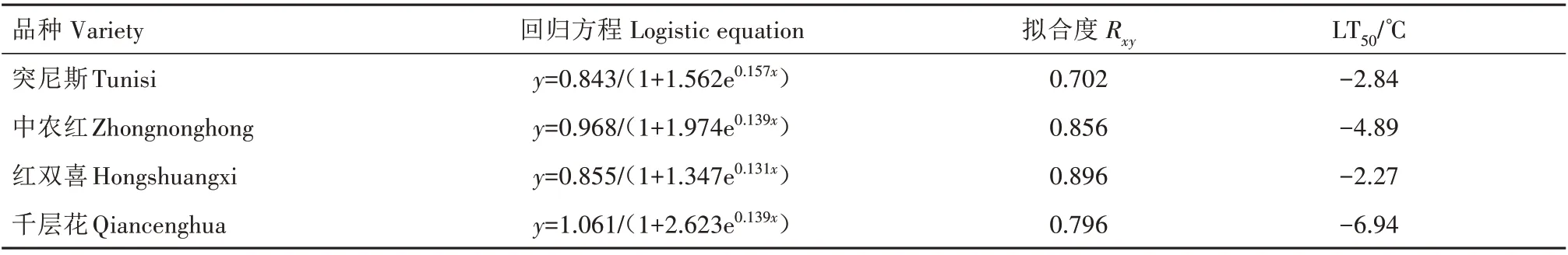
表1 低温胁迫下4个石榴品种的LT50Tab.1 LT50 of four pomegranate varieties under low temperature stress
2.3 石榴生理指标间相关性分析
如图9所示,脯氨酸含量与相对电导率、SOD 活性和可溶性糖含量呈极显著正相关(P<0.01),相对电导率与CAT 活性、可溶性蛋白含量呈负相关,与MDA 含量呈正相关,但未达到显著水平,说明低温胁迫时,细胞膜被破坏,电解质外渗,CAT 与可溶性蛋白起到保护作用。CAT 与POD 活性呈显著正相关(P<0.05),但与可溶性蛋白含量呈显著负相关。POD 活性与可溶性糖含量呈极显著性正相关,相关系数为0.46。由此可见,各抗寒指标间存在一定联系,不能以单个指标判断其抗寒性。
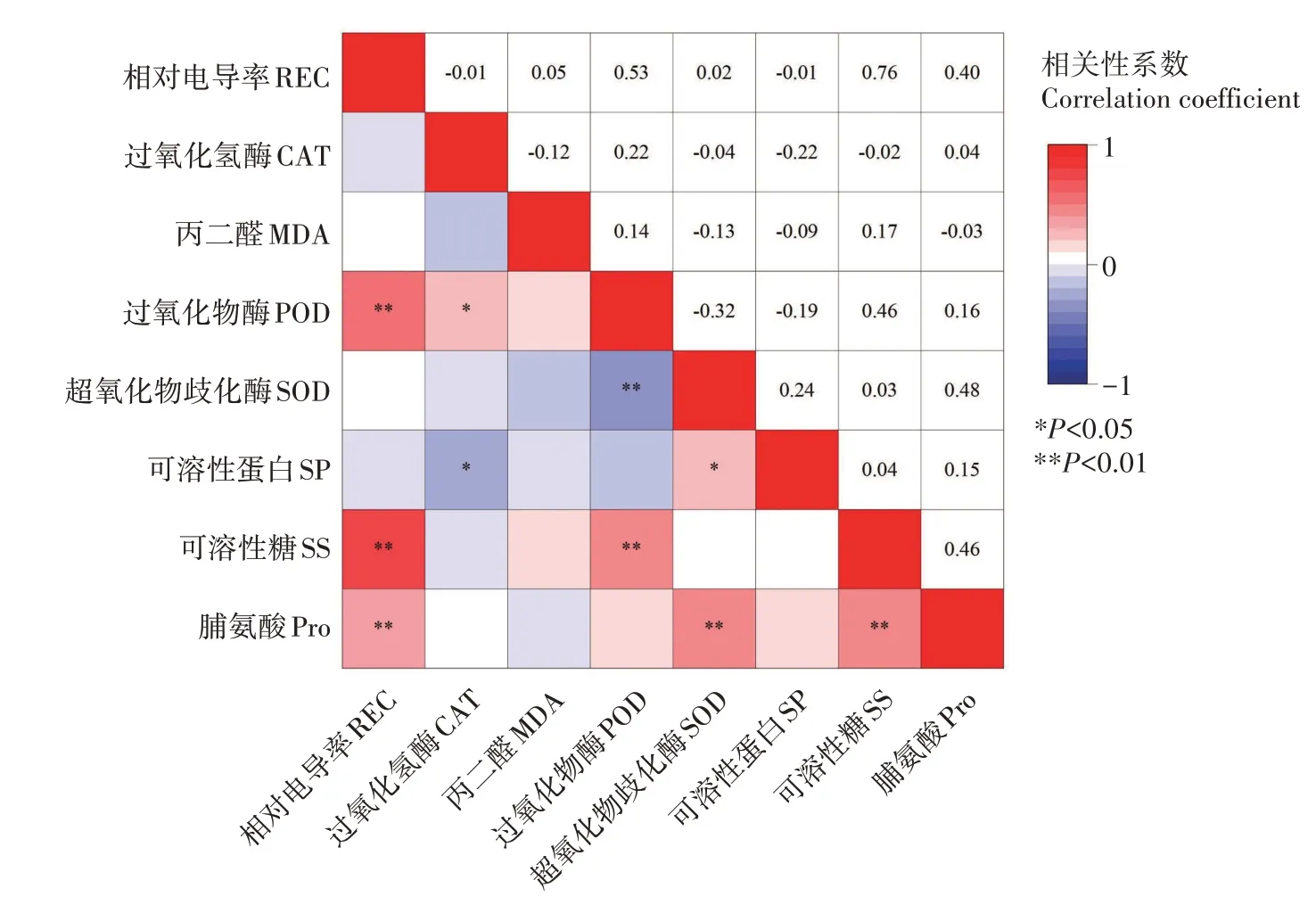
图9 石榴生理指标间相关性分析Fig.9 Correlation analysis between physiological indicators of pomegranate
2.4 4个石榴品种抗寒性综合评价
如表2 所示,对石榴抗寒性密切相关的8 个生理指标即脯氨酸含量、可溶性糖含量、可溶性蛋白含量、相对电导率、SOD 活性、POD 活性、CAT 活性和MDA 含量进行综合评价,结果表明,千层花的综合隶属度最大,为0.56,中农红综合隶属度最小,为0.45。根据隶属函数值大小,对4 个石榴品种的抗寒性进行排序,依次为千层花>红双喜>突尼斯>中农红。

表2 4个石榴品种生理指标抗寒隶属函数值及综合评价Tab.2 Cold resistant subordinate function values and comprehensive evaluation of physiological indexes of four pomegranate varieties
3 结论与讨论
3.1 低温胁迫与相对电导率之间的关系
植物本身具有抗逆机制,即通过调节一系列生理生化和分子生物学变化来适应寒冷、干旱和高盐等非生物胁迫[23]。植物的抗寒能力强弱是影响植物生长发育、决定其地理分布的重要因素,自然情况下植物受到低温胁迫时,会随着温度的不断降低,发生植物细胞膜透性、渗透调节物质、酶活性和外观形态等方面的改变[24]。
相对电导率能反映细胞膜的渗透性和破坏程度[25],植物细胞膜本身具有选择透过性,在细胞的正常代谢活动中发挥了重要作用。前人研究表明,当抗寒力较弱的植物受到低温胁迫时,细胞膜透性增大,从而引起胞内电解质外渗,细胞受损严重,导致相对电导率增加[26],因此,电导法已经成为反映植物抗寒能力的主要方法,广泛用于各种植物抗寒力的研究。李玲等[27]运用电导法鉴定橡胶树幼苗不同组织及品种间抗寒性强弱,得出结果与恢复生长法结果一致;井俊丽等[28]使用电导法协同Logistic 方程对9 种苹果砧木抗寒性进行比较;裘珍飞等[29]应用电导法评价了5 种桉树抗寒性。本研究中,随着温度的降低,4个品种石榴枝条相对电导率逐渐上升,呈近似“S”形变化曲线,这与郭伟珍等[30]研究结果一致。利用电导法结合Logistic 方程得出LT50值的大小,能更直观地判断果树的抗寒性强弱。本研究通过测定4 个石榴品种1 年生枝条在低温胁迫下相对电导率大小,初步判断其抗寒性强弱(LT50)分别为千层花(-6.94 ℃)>中农红(-4.89 ℃)>突尼斯(-2.84 ℃)>红双喜(-2.27 ℃)。
3.2 低温胁迫与MDA含量之间的关系
MDA 含量能初步反映膜系统损伤程度和植物抗逆性[31],当细胞膜的过氧化程度加重,导致其产物MDA 大量积累,植物细胞膜结构遭到破坏,细胞代谢紊乱,严重会导致死亡,所以MDA 含量与植物抗寒性呈负相关。姜良宝[32]发现,随着胁迫温度的降低,耐寒性较强的梅花的MDA 含量缓慢升高,而耐寒性最弱的梅花的MDA 含量呈现先升后降的变化趋势。在本研究中,低温胁迫下4 个石榴品种的MDA 含量变化整体呈先逐渐增加后减少的变化规律,这与郑元等[33]研究的3 个仁用杏品种枝条MDA含量先上升后下降的变化趋势一致,可以推断耐寒石榴品种可以忍耐MDA含量在一定范围的增加,耐寒性最强的千层花的MDA 含量始终低于其他3 个品种,表明耐寒性强的石榴品种忍耐细胞膜过氧化胁迫的能力也高。
3.3 低温胁迫与渗透调节物质含量之间的关系
植物受到低温刺激后细胞膜受损,通过合成和积累脯氨酸、可溶性糖和可溶性蛋白等重要的渗透调节物质来抵御逆境[34],起到保护细胞膜的作用[35]。本研究发现,随着温度降低,耐寒性不同的石榴品种可溶性蛋白和可溶性糖含量都有所增加,可能是细胞遭受低温胁迫后积累形成新的蛋白质,糖类大分子物质积累合成可溶性糖,细胞保水能力增强,对细胞膜起保护作用。当温度持续下降时,红双喜和千层花可溶性蛋白含量逐渐降低,表明其渗透调节代谢系统遭到破坏,可溶性蛋白分解速率加快,导致含量逐渐下降。低温胁迫下,脯氨酸的增加可以保持细胞内渗透平衡,从而减少细胞膜的破坏来抵御低温伤害[36]。本研究中脯氨酸含量变化与白淼等[37]报道的脯氨酸含量整体呈先升后降的变化趋势有所不同,可能由于本研究持续低温的缘故,也有前人[38]研究认为抗寒性与脯氨酸含量之间并没有很强的相关性。
3.4 低温胁迫与抗氧化酶活性之间的关系
胞内活性氧大量积累也会诱导抗氧化防御系统,主要包括SOD、POD 和CAT 等抗氧化酶,其对活性氧进行清除,保护细胞免遭伤害,因此与植物抗寒性呈显著的正相关关系。闫彪[39]通过对油菜幼苗进行不同时间的低温处理,其中的SOD、POD 和CAT 活性均表现出先升高后降低的趋势。杨雪梅等[40]以皮亚曼、牡丹、泰山红和泰山三白为材料,研究发现,4 个石榴品种枝条中相对电导率和POD 活性随着处理温度降低而逐渐升高,SOD 和CAT 活性呈先升高后降低的变化趋势,这与本研究结果基本一致。本研究测得的SOD 和CAT 活性在一定温度范围内随着温度下降而逐渐升高,说明该阶段内低温诱导SOD 和CAT 活性增加,清除细胞内产生的活性氧,保护细胞膜系统不被伤害,而随着温度持续下降,高浓度的活性氧导致酶蛋白分子破坏,SOD和CAT 活性降低,膜脂过氧化程度加重。POD 活性整体随着温度降低逐渐升高,这可能与POD 在植物中具有双重性有关,在低温胁迫前期,清除H2O2,表现为保护效应,为细胞活性氧保护酶系统的成员之一,后期参与活性氧的生成,表现为伤害效应,破坏细胞膜系统,使枝条遭受冻害[41]。
综上,石榴的抗寒性受多种因素共同影响,应结合渗透调节物质及抗氧化酶等指标,运用隶属函数法进行综合评价,本研究得出淅川县4 个品种石榴枝条的抗寒性依次表现为千层花>红双喜>突尼斯>中农红。
