盐碱胁迫下不同倍性水稻4 种转录因子家族基因的表达模式分析
2023-07-18瞿小杰刘金海彭赞文
黄 海,瞿小杰,刘金海,彭赞文
(梧州学院,广西 梧州 543002)
水稻(Oryza sativaL.)是我国主要的粮食作物之一,对保障国家粮食安全具有极其重要的作用。但是,我国土壤盐碱化情况比较严重,土壤盐碱含量过高会使水稻出现生理性干旱,导致离子毒害、渗透胁迫、氧化胁迫和营养胁迫,影响水稻生长发育进而影响其产量和品质[1]。多倍体化是驱动植物进化的主要动力[2]。多倍体植物由于染色体加倍或者不同染色体组间的相互结合,使基因组结构和基因表达模式不断发生变化,并且基因剂量得到增加,有时会增加次生代谢物质含量,进而促使多倍体植物的抗逆性增强,更容易适应外界环境变化[3]。对于多倍体水稻而言,染色体加倍后的四倍体水稻与二倍体相比,具有植株茎秆粗壮[4]、籽粒大[5]、蛋白质含量高[6]、抗性强等优点[7]。经过长期演化,水稻形成一系列防御机制以响应盐碱胁迫。其中,在分子水平上,一些转录因子可以调控水稻对盐碱胁迫的抗性[8]。水稻中至少存在1 611 个转录因子基因,约占基因组的2.6%,一些转录因子家族拥有上百个基因,比如bZIP、MYB、AP2/ERF 和WRKY 等家族,这些转录因子参与水稻生长发育及对生物和非生物胁迫抗性的调控,具有极其重要的生物学功能[9-12]。前人研究发现,bZIP、MYB、AP2/ERF、WRKY转录因子家族中的一些基因可以响应水稻干旱、高盐、低温等非生物胁迫,超表达这些基因可以不同程度地提高转基因水稻的抗旱性、耐盐性和耐冷性等[9-12]。目前,关于抗逆相关转录因子对盐碱胁迫的响应机制研究主要集中于二倍体水稻[9-13],在四倍体水稻上的研究鲜有报道。为此,以二倍体水稻桂育12及其同源四倍体水稻为试验材料,研究二倍体、四倍体水稻中bZIP、MYB、AP2/ERF 和WRKY 转录因子家族基因在盐碱胁迫条件下的表达模式变化,为解析不同倍性水稻的耐盐碱机制奠定基础。
1 材料和方法
1.1 试验材料
供试水稻材料为二倍体水稻桂育12(以下简称为桂育12D,由广西壮族自治区农业科学院水稻研究所提供)及其同源四倍体水稻(以下简称为桂育12T,经过秋水仙素加倍得到)。
1.2 水稻培养及处理
选取大小均匀、籽粒饱满的水稻种子,用0.3%NaClO 浸泡消毒15 min,然后用蒸馏水清洗,之后将种子放置在铺有2 层滤纸的培养皿中,35 ℃暗培养3 d,最后置于16 h光照(28 ℃)/8 h黑暗(25 ℃)的人工气候箱中用水稻营养液在育苗盒中培养[14]。待幼苗长至三叶一心期时,分别用pH 值11.39 的NaOH(A)和100 mmol/L NaCl(S)配制的营养液胁迫处理6 h,对照组(CK)使用正常营养液培养6 h,每个材料设置3次重复。
1.3 RNA提取及实时荧光定量PCR(qRT-PCR)
RNA 提 取 参 照TransZol Up Plus RNA Kit 试 剂盒说明书进行;RNA 反转录参照TransScript®One-Step gDNA Removal and cDNA Synthesis SuperMix试剂盒说明书进行,反应程序:42 ℃15 min,85 ℃5 s。qRT-PCR 参 照TransStart®Top Green qPCR SuperMix 试剂盒说明书进行,反应体系20 μL:ddH2O 7 μL,2×TransStart Top Green qPCR Super Mix 10 μL,上、下游引物(0.1 ng/μL)各1 μL,反转录产物1 μL;反应程序:94 ℃30 s;94 ℃15 s、60 ℃5 s 、72 ℃10 s,45 个循环。上述试剂盒均购自北京全式金生物技术有限公司,每个处理设3 个生 物 学 重 复 。内 参 基 因 为GAPDH(Glyceraldehyde-3-phosphate dehydrogenase),GAPDH及bZIP、MYB、WRKY、AP2/ERF 转录因子家族基因的引物设计参考qPrimerDB 数据库[15],所有基因引物序列详见表1。基因的相对表达量计算采用 2-△△Ct法[16]。
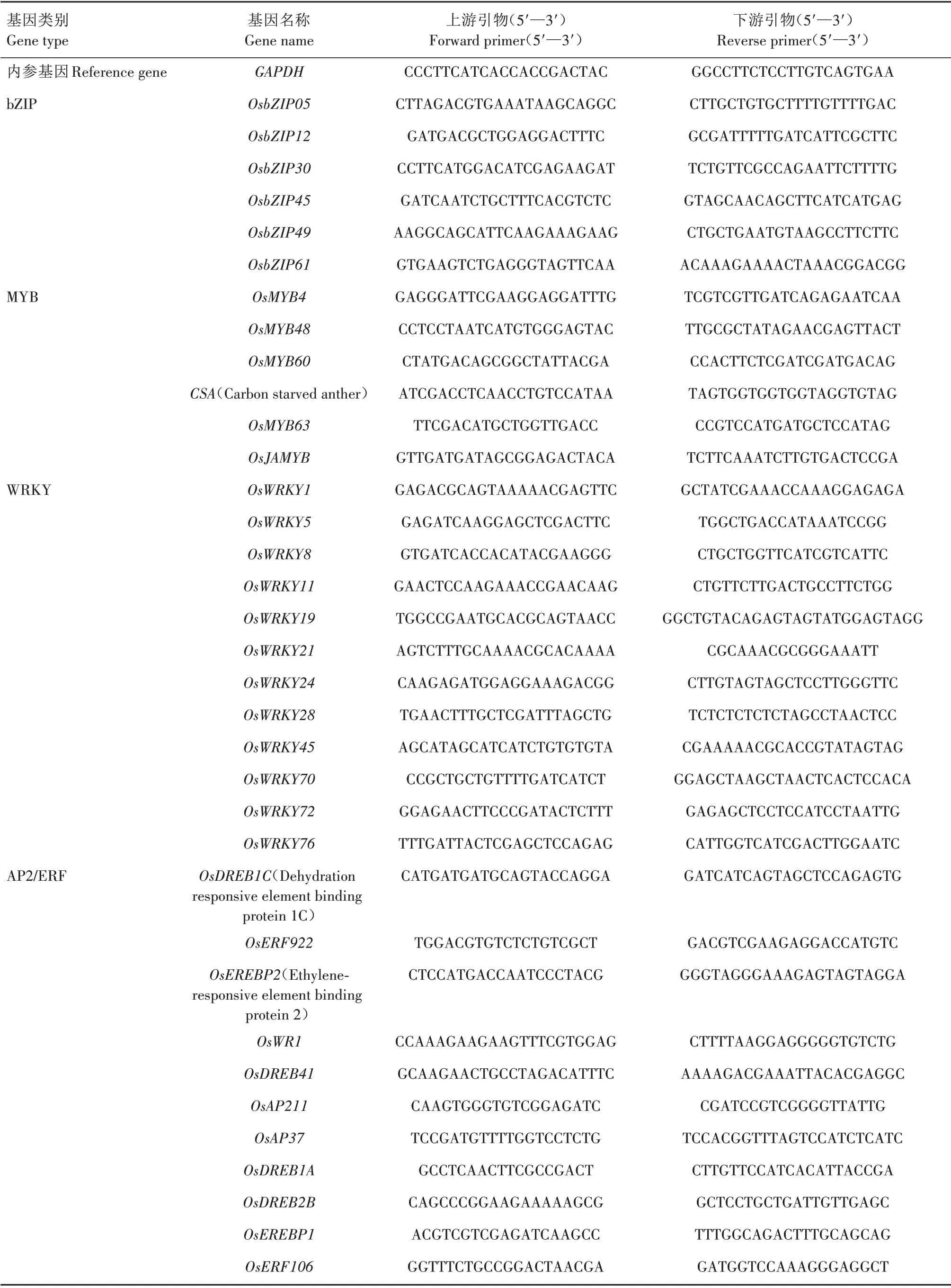
表1 qRT-PCR 所用引物Tab.1 Primers used for qRT-PCR
1.4 数据统计分析
使用Excel 2021 软件整理试验数据,采用DPS 17.10软件中Tukey法进行多重比较分析。
2 结果与分析
2.1 盐碱胁迫对不同倍性水稻bZIP转录因子家族基因表达的影响
bZIP 是由一个亮氨酸拉链二聚体结构域和一个DNA 结合域组成的一类重要的植物转录因子,参与植物生长、胚乳发育、叶片衰老以及各种生物和非生物胁迫的响应等[17-19]。本研究选取的6 个bZIP 转录因子家族基因在盐碱胁迫下的表达情况如图1 所示。由图1 可以看出,无胁迫条件下,桂育12D 中的OsbZIP30和OsbZIP61基因表达量均极显著高于桂育12T,而OsbZIP49基因相反,其他基因无极显著差异。在盐胁迫条件下,桂育12D 和桂育12T 中OsbZIP05和OsbZIP12基因表达量均极显著提高;桂育12T中OsbZIP30和OsbZIP45基因表达量均极显著提高;对于OsbZIP49基因,在桂育12T 中表达量极显著下降,在桂育12D 中表达量提高但未达到极显著水平,而OsbZIP61基因与OsbZIP49基因完全相反。在碱胁迫条件下,桂育12D 中OsbZIP12、OsbZIP45、OsbZIP49和OsbZIP61基因表达量均极显著提高,但在桂育12T 中除OsbZIP49基因表达量极显著下降、OsbZIP61基因表达量极显著提高外,其他基因均无极显著差异;桂育12D 和桂育12T 中OsbZIP05和OsbZIP30基因表达量均无极显著差异,OsbZIP61基因表达量均极显著提高,且桂育12T 中提高幅度大于桂育12D。综上,多数bZIP 转录因子家族基因表达量会受盐胁迫诱导上调,且在四倍体水稻中受诱导上调幅度大于二倍体水稻;在碱胁迫条件下,多数bZIP 转录因子家族基因表达量在二倍体水稻中提高,在四倍体水稻中无极显著差异。

图1 盐碱胁迫下不同倍性水稻bZIP转录因子家族基因的表达分析Fig.1 Expression analysis of bZIP transcription factor family genes in different ploidy rice under saline-alkali stress
2.2 盐碱胁迫对不同倍性水稻MYB转录因子家族基因表达的影响
MYB 转录因子因其N 端保守的MYB 结构域而得名,是一类存在最广、功能最强的转录因子,既能调控植物的生长发育,又可以参与植物对高盐、干旱、极端温度、病虫害侵袭等逆境胁迫的抗性反应[20]。本研究选取的6 个MYB 转录因子家族基因在盐碱胁迫下的表达情况如图2所示。由图2可以看出,无胁迫条件下,桂育12D中CSA基因表达量极显著高于桂育12T,而OsMYB4和OsJAMYB基因相反,其他基因无极显著差异。在盐胁迫条件下,桂育12D 和桂育12T 中OsMYB4和OsJAMYB基因表达量均极显著提高,OsMYB48和OsMYB63基因表达量均极显著下降;桂育12D 中CSA基因表达量极显著下降,但在桂育12T 中极显著提高。在碱胁迫条件下,桂 育12D 和 桂 育12T 中OsMYB4、OsMYB48、OsMYB60和OsMYB63基因表达量均无极显著差异,CSA基因表达量极显著下降,OsJAMYB基因表达量极显著提高,且在桂育12T 中的提高幅度高于桂育12D。总体来看,OsMYB60基因表达量在盐碱胁迫条件下均无极显著差异。综上,在盐胁迫条件下,不同倍性水稻中同一MYB 基因对盐胁迫的响应有所差别;在碱胁迫条件下,OsJAMYB和CSA基因均被极显著诱导表达,且在四倍体水稻中的提高或者下降幅度大于二倍体水稻,其他MYB基因无极显著差异。

图2 盐碱胁迫下不同倍性水稻MYB转录因子家族基因的表达分析Fig.2 Expression analysis of MYB transcription factor family genes in different ploidy rice under saline-alkali stress
2.3 盐碱胁迫对不同倍性水稻WRKY转录因子家族基因表达的影响
WRKY 转录因子因存在高度保守的WRKYGQK 结构域而得名,在植物形态建成、物质代谢、生物及非生物胁迫响应的过程中起到重要的调控作用[21-22]。本研究选取的12 个WRKY 转录因子家族基因在盐碱胁迫下的表达情况如图3 所示。由图3 可以看出,无胁迫条件下,桂育12T 中OsWRKY1、OsWRKY5、OsWRKY8、OsWRKY11、OsWRKY24、OsWRKY28、OsWRKY45、OsWRKY72和OsWRKY76基因表达量均极显著高于桂育12D,其他基因无极显著差异。在盐胁迫条件下,桂育12D和 桂 育12T 中OsWRKY1、OsWRKY8、OsWRKY11、OsWRKY24、OsWRKY70和OsWRKY72基因表达量均提高但未达到极显著水平;桂育12D 中OsWRKY5基因表达量极显著提高,OsWRKY19、OsWRKY21、OsWRKY45和OsWRKY76基因表达量均无极显著差异,但在桂育12T 中均极显著下降。在碱胁迫条件下,桂育12D 中OsWRKY1、OsWRKY8、OsWRKY11、OsWRKY21、OsWRKY24、OsWRKY28、OsWRKY45、OsWRKY72和OsWRKY76基因表达量均极显著提高,OsWRKY5、OsWRKY19和OsWRKY70基因表达量均无极显著差异;桂育12T 中OsWRKY8、WRKY45、OsWRKY70、OsWRKY72和OsWRKY76基因表达量均极显著下降,其他基因表达量均无极显著差异。综上,在二倍体水稻中大多数WRKY 基因对盐胁迫无极显著响应,但在四倍体水稻中,盐胁迫抑制较多WRKY 基因的表达;在碱胁迫条件下,二倍体水稻中多数WRKY 基因表达量提高,而在四倍体水稻中多数下降,表明碱胁迫下WRKY 基因在二倍体、四倍体水稻中发挥的作用相反。

图3 盐碱胁迫下不同倍性水稻WRKY转录因子家族基因的表达分析Fig.3 Expression analysis of WRKY transcription factor family genes in different ploidy rice under saline-alkali stress
2.4 盐碱胁迫对不同倍性水稻AP2/ERF转录因子家族基因表达的影响
AP2/ERF 转录因子以含有高度保守的AP2/ERF 结构域而得名,该结构域具有DNA 结合功能,与植物生长发育、生物合成以及逆境应答密切相关[23-24]。本研究选取的11个AP2/ERF转录因子家族基因在盐碱胁迫下的表达情况如图4 所示。由图4可以看出,无胁迫条件下,桂育12D 中OsDREB1C、OsEREBP1、OsEREBP2、OsAP211、OsERF106、OsERF922和OsWR1基因表达量与桂育12T 均无极显著差异;桂育12T 中OsDREB1A基因表达量极显著高于桂育12D,OsDREB2B、OsDREB41和OsAP37基因相反。在盐胁迫条件下,桂育12D 和桂育12T中OsDREB1A、OsDREB1C和OsAP37基因表达量均极显著提高;桂育12D中OsEREBP2和OsERF922基因表达量均极显著提高,在桂育12T 中虽提高但未达到极显著水平;桂育12D 中OsDREB2B、OsEREBP1、OsERF106和OsWR1基因表达量均提高但未达到极显著水平,OsDREB41、OsAP211基因表达量均下降但未达到极显著水平;桂育12T 中OsDREB41、OsAP211和OsWR1基因表达量均极显著提高。在碱胁迫条件下,桂育12D 和桂育12T 中OsDREB41基因表达量均无极显著差异;桂育12D中OsAP211基因表达量提高但未达到极显著水平,OsWR1基因表达量下降但未达到极显著水平,在桂育12T中也均无极显著差异;桂育12D中OsDRE2B、OsEREBP1、OsAP37、OsERF106和OsERF922基因表达量均无极显著差异,但在桂育12T中极显著提高;桂育12D中OsDREB1C、OsEREBP2基因表达量均极显著提高,但在桂育12T 中无极显著差异;桂育12D中OsDREB1A基因表达量极显著提高,但在桂育12T 中极显著下降。综上,本研究所选的AP2/ERF基因多数受盐胁迫诱导表达量提高,但在不同倍性水稻中对盐胁迫的响应不同;在碱胁迫条件下,二倍体水稻中大多数AP2/ERF 基因表达量无极显著差异,而在四倍体水稻中较多AP2/ERF 基因表达量极显著提高。

图4 盐碱胁迫下不同倍性水稻AP2/ERF转录因子家族基因的表达分析Fig.4 Expression analysis of AP2/ERF transcription factor family genes in different ploidy rice under saline-alkali stress
3 结论与讨论
水稻是重要的粮食作物,提高水稻抗逆性、扩大水稻种植面积对确保我国粮食安全具有重要作用[25-26]。许多研究证实,转录因子在植物对非生物胁迫的响应中发挥了重要的枢纽作用[27],但多针对于二倍体水稻,四倍体水稻在此方面的研究则鲜有报道。因此,在盐碱胁迫下研究二倍体、四倍体水稻转录因子家族基因表达模式的变化,对探究不同倍性水稻的耐盐碱机制具有一定指导意义。
研究发现,OsbZIP05[28]和OsbZIP12[29]基因的表达与水稻耐盐性存在正相关关系。本研究发现,二倍体、四倍体水稻中OsbZIP05和OsbZIP12基因被盐胁迫诱导表达并达到极显著水平,且在四倍体水稻中受盐胁迫诱导程度更高;在碱胁迫条件下,二倍体水稻中OsbZIP12基因表达量极显著提高,而在四倍体水稻中无极显著差异。表明OsbZIP05和OsbZIP12基因主要在盐胁迫条件下二倍体、四倍体水稻和碱胁迫条件下二倍体水稻中发挥功能,在碱胁迫条件下四倍体水稻中没有发挥主要功能。另有研究发现,OsbZIP45基因可以正调控水稻耐旱性[30]。在本研究中,二倍体水稻中OsbZIP30和OsbZIP45基因表达量在盐胁迫前后无极显著差异,而在四倍体水稻中极显著提高,表明OsbZIP30和OsbZIP45基因在二倍体水稻对盐胁迫的响应中不发挥主要功能,可能在四倍体水稻中正向响应盐胁迫;在碱胁迫条件下,四倍体水稻中OsbZIP30和OsbZIP45基因表达量无极显著差异,OsbZIP45基因表达量在二倍体水稻中提高,表明OsbZIP45基因可能在碱胁迫条件下二倍体水稻中发挥了一定功能。二倍体水稻中OsbZIP49基因的表达受盐胁迫诱导,四倍体水稻中OsbZIP49基因的表达受盐胁迫抑制,碱胁迫与盐胁迫趋势一致,表明该基因在二倍体、四倍体水稻对盐碱胁迫的应激反应中发挥了相反的作用;而盐胁迫条件下OsbZIP61基因表达量的变化趋势与OsbZIP49基因相反,在碱胁迫条件下二倍体、四倍体水稻中均极显著提高,推测该基因在水稻对盐碱胁迫的应激反应中发挥的作用可能与OsbZIP49基因不同,有待进一步研究。
研究发现,OsJAMYB基因与茉莉酸介导的非生物胁迫途径相关,在水稻苗期,高盐胁迫能诱导OsJAMYB基因的表达,进而激活一些防御相关基因的转录表达[31]。本研究发现,二倍体、四倍体水稻中OsMYB4和OsJAMYB基因在盐胁迫条件下的表达量均高于对照组,且OsJAMYB基因的表达量差异在二倍体、四倍体水稻中均达到极显著水平;在碱胁迫条件下,OsJAMYB基因变化规律与盐胁迫一致,但OsMYB4基因表达量无极显著差异,表明OsJAMYB基因在盐碱胁迫条件下可能均发挥功能,而OsMYB4基因可能仅在盐胁迫条件下发挥功能。另外,二倍体、四倍体水稻中OsMYB63基因的表达量被盐胁迫极显著抑制,且对四倍体水稻的抑制程度更高,在碱胁迫条件下无极显著差异,表明该基因可能仅负调控水稻耐盐性。研究发现,OsMYB60[32]和CSA[33]基因可以通过调控水稻表皮角质蜡的生物合成和ABA(脱落酸)稳态正向调控水稻幼苗对非生物胁迫的耐受性。但在本研究中,OsMYB60基因表达量在盐碱胁迫条件下均无极显著差异,推测该基因可能在水稻对盐碱胁迫的响应中不发挥功能;而二倍体水稻中CSA基因的表达受盐碱胁迫极显著抑制,四倍体水稻中CSA基因表达量在盐胁迫条件下极显著提高,在碱胁迫条件下均极显著下降,表明二倍体、四倍体水稻CSA基因对盐胁迫的应激反应不同。研究发现,OsMYB48-1[34-35]基因受高盐胁迫诱导表达,超表达OsMYB48-1基因增强了转基因水稻植株对ABA 的敏感性,从而提高了水稻耐盐性。但在本研究中,二倍体、四倍体水稻中OsMYB48基因的表达均被盐胁迫抑制并达到极显著水平,在碱胁迫条件下无极显著差异,说明盐胁迫会抑制该基因的表达,这与前人[34-35]研究结果不一致,推测可能是本试验材料中OsMYB48基因可能是OsMYB48-1基因的另一等位基因,它们在水稻对盐碱胁迫的响应中发挥了不同的作用。
研究发现,OsWRKY8、OsWRKY11、OsWRKY24、OsWRKY45和OsWRKY72基因参与调节水稻对渗透胁迫、非生物胁迫的应答[36-40]。在本研究中,OsWRKY8、OsWRKY11、OsWRKY24、OsWRKY70和OsWRKY72基因在二倍体、四倍体水稻中的表达量受盐胁迫诱导上调但未达到极显著水平,OsWRKY1基因的变化趋势与之类似,推测在二倍体、四倍体水稻对盐胁迫的响应中OsWRKY1基因与它们发挥了相同的功能。四倍体水稻中OsWRKY5、OsWRKY19、OsWRKY21、OsWRKY28和OsWRKY76基因表达量极显著下降,与OsWRKY45基因变化趋势一致,推测可能在四倍体水稻抗盐应激反应中发挥了一定功能。OsWRKY76基因在水稻萌发过程中对耐碱性可能起负调控作用[41]。在碱胁迫条件下,四倍 体 水 稻 中OsWRKY8、OsWRKY45、OsWRKY70和OsWRKY72基因表达量均极显著下降,与OsWRKY76基因变化趋势一致,表明这4 个基因可能与OsWRKY76基因发挥了一样的功能,并且OsWRKY8、OsWRKY45、OsWRKY72和OsWRKY76基因表达量在四倍体水稻中均极显著下降,在二倍体水稻中均极显著提高,推测它们对碱胁迫的应答机制相反。综上,本研究中四倍体水稻中多数WRKY 基因的表达均被盐碱胁迫抑制,所以四倍体水稻WRKY 基因对盐碱胁迫的响应机制有待于进一步探索。
研究发现,OsDREB1A和OsDREB1C基因参与调控植物对冷胁迫的耐受性[42]。本研究中,OsDREB1A和OsDREB1C基因在盐胁迫条件下二倍体、四倍体水稻中和碱胁迫条件下二倍体水稻中的表达均被诱导且达到极显著水平,推测OsDREB1A和OsDREB1C基因可能在多种非生物胁迫的响应中存在冗余功能。过表达OsEREBP1基因能提升水稻生物和非生物胁迫的耐受性[43]。在本研究中,碱胁迫条件下四倍体水稻中OsEREBP1基因表达量极显著提高,表明OsEREBP1基因在四倍体水稻应对碱胁迫中发挥了一定作用。OsEREBP2[44]和OsERF922[45]基因分别是盐胁迫的正、负调控因子,它们的表达受盐胁迫强烈诱导。本研究中,OsEREBP2和OsERF922基因表达量在盐胁迫条件下二倍体水稻中极显著提高。在四倍体水稻中虽提高但未达到极显著水平。碱胁迫条件下,二倍体水稻OsEREBP2基因表达量极显著提高,OsERF922基因无极显著差异;但在四倍体水稻中OsEREBP2基因表达量下降但未达到极显著水平,而OsERF922基因表达量极显著提高。推测OsEREBP2和OsERF922基因可能也参与了碱胁迫应答,但响应机制尚不明确。此外,OsWR1[46]基因主要在水稻叶片中表达,且受干旱、ABA 和盐胁迫的诱导。本研究中,二倍体、四倍体水稻中OsWR1基因的表达均被盐胁迫诱导,在四倍体水稻中达到极显著水平,提高幅度大于二倍体水稻,在碱胁迫条件下无极显著差异,说明四倍体水稻中OsWR1基因的表达只受盐胁迫诱导。对于OsDREB41和OsAP211基因,它们的表达在二倍体水稻中均被盐胁迫抑制,在四倍体水稻中则均被极显著诱导,而在碱胁迫条件下无极显著差异,表明OsDREB41和OsAP211基因可能只在盐胁迫中发挥功能。
综上所述,在盐碱胁迫条件下,本研究中所选的转录因子家族基因在二倍体、四倍体水稻中的表达模式是不同的,主要体现在表达程度以及表达趋势上,可能是因为:虽然bZIP、MYB、WRKY 和AP2/ERF 基因均与非生物胁迫相关,但它们在盐碱胁迫响应过程中扮演角色的重要性不同[47];另外,转录因子可能不通过单一基因直接调控水稻耐盐碱性,而是通过多基因协同或拮抗机制形成一个复杂的调控网络,进而通过基因的多重表达来抵御盐碱胁迫[48]。本研究中,在无胁迫条件下,四倍体水稻中一些转录因子基因表达量极显著高于二倍体,如OsbZIP49、OsMYB4、OsJAMYB、OsWRKY1和OsDREB1A基因等;在受到盐碱胁迫后,二倍体、四倍体水稻中的多数转录因子基因被胁迫极显著诱导表达,且在四倍体水稻中提高或下降程度更高,如OsbZIP05、OsbZIP12、OsJAMYB、OsMYB48基 因等;四倍体水稻中也有少数转录因子基因在受到盐碱胁迫后表达趋势与二倍体水稻完全相反,如OsbZIP49、OsWRKY8、OsWRKY45、OsWRKY76和OsDREB1A基因等。导致这些现象的原因可能是染色体加倍致使一些基因剂量加倍,使二倍体、四倍体水稻在正常条件下基因的表达产生明显差异。在胁迫条件下,由于在多倍体基因组中,多数重复基因仍保持着与二倍体基因组中相似或相同的功能,染色体加倍带来基因剂量效应的优势被突显,进而使四倍体水稻中胁迫相关基因增量响应,提高基因的表达程度,增强四倍体水稻的抗性[49];并且染色体加倍后的四倍体水稻中部分基因可能触发了RNAi 机制,导致四倍体水稻中一些转录因子基因的表达量在正常条件下极显著低于二倍体水稻,并且在胁迫前后二倍体、四倍体水稻中出现相反的趋势。此外,染色体加倍过程中会发生不同程度的染色体结构变异,包括染色体的倒位、异位、缺失、重复和变异,进而使重复基因发生突变、沉默和重组,甚至产生与原基因功能完全不同的新基因[50]。本研究为解析多倍体水稻中bZIP、MYB、AP2/ERF 和WRKY基因的耐盐碱机制奠定了基础。
