苦瓜皂苷对饲喂高糖饲料松浦镜鲤幼鱼生长、消化及肝脏健康的影响
2023-07-12范泽吴迪李晨辉李晋南刘庆武王连生
范泽,吴迪,李晨辉,2,李晋南,刘庆武,王连生
(1.中国水产科学研究院黑龙江水产研究所,黑龙江省水生动物病害与免疫重点试验室,黑龙江 哈尔滨 150070;2.农业农村部水产品质量安全控制重点实验室,北京 100141;3.黑龙江省双鸭山市农业技术推广总站,黑龙江 双鸭山 155100)
淀粉是养殖鱼类饲料中最重要的糖类来源,具有节约饲料蛋白质的作用。鱼类自身“先天性糖尿病体质”使得糖类节约饲料蛋白效应未能有效发挥[1]。鱼类摄食高糖饲料后消化能力下降[2]、肝脏损伤[3,4],免疫力减弱[5,6]。而一些植物提取物如皂苷类、类黄酮类、黄酮类等有潜在的防治这类代谢疾病的作用[7,8]。近年来,天然植物中皂苷类化合物的作用越来越受到重视。皂苷绿色、无毒害,且具有降血脂、抗氧化、抗菌抗炎症等功能,成为值得深入开发与利用的生物活性物质。
苦瓜皂苷是苦瓜中的有效成分之一,在抗氧化、提升免疫力、降低胆固醇、抑菌等方面效果显著[9]。王先远等[10]发现,苦瓜皂苷可显著增强人体超氧化物歧化酶,谷胱甘肽活力,其机理可能与肿瘤坏死因子(TNF)的作用有关。苦瓜皂苷对高血糖模型小鼠具有明显降低血糖作用和抗氧化能力的恢复作用;灌胃120 mg/kg 苦瓜皂苷30 d 降血糖作用和恢复抗氧化能力的效果较好。目前苦瓜皂苷在水产动物营养免疫方面的研究还未见相关报道。
Kelch 样环氧氯丙烷相关蛋白1(Keap1)-核因子E2 相关因子2(Nrf2)/抗氧化反应元件(ARE)信号通路是水产动物抵抗内、外源氧化损伤的关键信号通路。在信号通路中,Nrf2 是促进下游抗氧化基因转录的关键信号蛋白,而Keap1 是Nrf2 的负调节蛋白[11]。研究证实:功能性营养物质可对水产动物Keap1-Nrf2/ARE 信号通路进行调节[12]。但苦瓜皂苷对Keap1-Nrf2/ARE 信号通路的调控机制尚未研究。
作为典型的杂食性鱼类,尽管与肉食性鱼类相比,鲤对于糖类的利用能力更胜一筹[1]。松浦镜鲤是我国主要的鲤养殖品种,具有良好的抗病和抗逆能力[13]。本实验室前期研究发现,摄食高糖饲料后松浦镜鲤肠道皱襞高度降低,肠道对营养物质的消化吸收减弱,并伴有肝脏病理损伤[14]。基于此,本实验通过研究不同水平苦瓜皂苷对松浦镜鲤肠道消化酶、肝脏抗氧化相关酶活性、基因表达及肝脏组织形态的影响,以期为初步揭示苦瓜皂苷在高糖饲料条件下改善鱼体健康的潜在机制,及苦瓜皂苷作为功能性营养物质在提高鱼类对淀粉高效利用上提供科学依据,并为缓解高糖饲料引发的鱼类健康问题提供理论基础。
1 材料与方法
1.1 材料
试验饲料以鱼粉、鸡肉粉、豆粕、大豆浓缩蛋白为蛋白质源,鱼油和豆油为脂肪源,以木薯淀粉为糖源,根据先前的研究结果[15],分别设置低糖(10%,LC)和高糖(30%,HC)两个对照组;高糖+4 个不同梯度的苦瓜皂苷水平:30%糖水平+100 mg/kg(MCS1)、400 mg/kg(MCS2)、1 600 mg/kg(MCS3)及6 400 mg/kg(MCS4)。所用饲料成分组成及营养水平见表1。苦瓜皂苷购自西安贝吉诺生物科技有限公司,纯度≥98.5%,其添加浓度依马春宇等[16]对苦瓜皂苷降血糖及抗氧化作用机制的研究结果设置。饲料制作:将所有干性原料经粉碎后,过80 目筛,按逐级放大原则使用混匀机搅拌混匀,然后依据饲料配方比例加入鱼油及豆油,在混匀机中充分混合后用饲料制粒机制成直径为1.50 mm 的沉性颗粒饲料。

表1 饲料组成和营养水平(干物质基础)Tab.1 Ingredient and nutrient levels of the diets used in the experiment(dry matter basis)
试验用松浦镜鲤幼鱼由中国水产科学研究院黑龙江水产研究所呼兰实验站提供,待用基础饲料(购自哈尔滨通威饲料有限公司,含30%蛋白和6%脂肪)暂养14 d 后挑取规格一致、体质健康、初始体质量为(6.43±0.02)g 的松浦镜鲤幼鱼在室内控温循环水养殖系统中开展饲养试验。
1.2 方法
根据试验饲料设置6 个处理组,每个处理组3个重复,每个重复25 尾鱼。试验鱼饲养于室内500 L 控温循环水族箱中,分别投喂6 组饲料,养殖56 d。整个试验期间,每天8:00 am、13:00 pm 和17:30 pm 投喂三次,日投喂率为7%~8%/(体质量·d)。每14 d 测定1 次各网箱鱼的质量,调整投喂量。采用多参数水质分析仪(HQ40D)(美国HACH 公司)监测养殖试验过程中主要水质参数。试验期间,水温变化在28℃~32℃之间,溶解氧量为6.0 mg/L 左右,pH7.6~8.0。
养殖试验结束时,停食24 h。每个重复随机取鱼3 尾,经100 mg/L 的MS-222 麻醉后,解剖取整条肠道,并将肠道内容排空,用0.9%生理盐水清洗干净后,保存于-20℃冰箱,用于测定消化酶;解剖取肝脏用于测定抗氧化相关指标;每个重复再随机取鱼4 尾,经同样麻醉后,剖取肝脏,2 尾鱼得肝脏用于抗氧化相关基因的检测,另2 尾鱼的肝脏(约0.5 cm3)用于测定与观察肝脏组织结构形态。
肠道及肝脏粗酶液的制备:将组织按照组织的0.9%生理盐水为1∶9 的比例配制成10%的组织匀浆液,4℃离心20 min 后吸取上清液,存于-20℃冰箱中备用。
肠道脂肪酶和淀粉酶活性用南京建成生物工程研究所测定试剂盒。其中检测淀粉酶活性的方法为碘-淀粉比色法,检测脂肪酶活性的方法为比浊法。利用酶联免疫试剂盒测定肠道蛋白酶活力。
肝脏抗氧化相关指标(过氧化氢酶CAT、过氧化物酶POD、谷胱甘肽GSH 活性及丙二醛MDA 含量)用南京建成生物工程研究所测定试剂盒。利用酶联免疫试剂盒测定肠道肝脏超氧化物歧化酶SOD 活力。
用于基因表达测定的肝脏经液氮迅速冷冻后,于-80℃冰箱中保存。利用Trizol 法提取各组织总RNA,参照TaKaRa PrimeScriptTM RT reagent Kit with gDNA Eraser(Perfect Real Time)(Code No:RR047A)说明书,将总RNA 反转录为cDNA。根据Genbank 中及相关文献中现有的鲤相关基因的保守序列设计引物。所有引物均由生工生物工程(上海)有限股份公司合成(表2)。实时荧光定量PCR 反应根据SYBRPremix Ex TaqTM(Tli RNaseH Plus)(Code: RR420A)试剂盒说明书进行,采用SYBRGreen 染色法,在Applied Biosystems 7500 Real Time PCR System 实时PCR 检测系统下进行。为了避免扩增批次间的差异,目的基因与其对应的内参基因同时扩增,每个模板均做三个重复,得到三个Ct 值,取其平均值作为最终Ct 值,后运用比较Ct 法(△△Ct)分析比较所得数据,得到各模板中各基因的相对表达量,计算公式为:
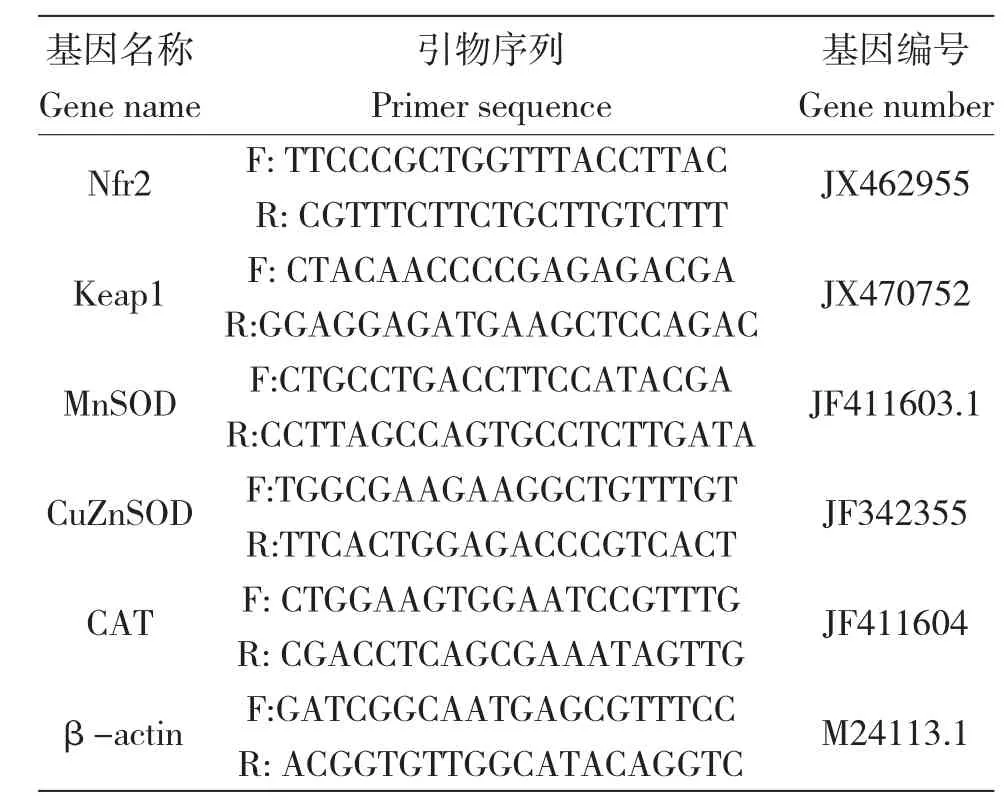
表2 试验所用引物Tab.2 Primers in present study
肝脏组织形态学观察:固定于4%多聚甲醛的肝脏组织通过梯度乙醇脱水、二甲苯透明、石蜡包埋、制成5 μm 切片、苏木精伊红染液(H&E)染色等步骤,制作成肝脏组织切片。切片在光学显微镜(Leica MD 2000B)下观察并拍照。
1.3 数据统计
生长指标的计算:
式中:t 代表试验天数(d);Nt代表实验结束时鱼存活尾数;W0为鱼初始体质量(g);Wt为鱼终末体质量(g);F 代表饲料摄取量干重(g)。
所有数据均以平均值±标准差表示,并利用SPSS23.0 统计软件进行单因素方差分析(One-way ANOVA),若差异达到显著水平(P<0.05),则进行Duncan's 法多重比较。
2 结果与分析
2.1 苦瓜皂苷对松浦镜鲤幼鱼生长指标的影响
由表3 可知,尽管HC 组与LC 组的终末均体质量及特定生长率无显著差异(P>0.05),但二者均低于LC 组。而HC 的饲料效率显著低于LC 组(P<0.05)。在高糖饲料添加苦瓜皂苷组中,MCS3 组鱼的终末均体质量、特定生长率、饲料效率及摄食率最高,显著高于HC 组(P<0.05),且MCS3 组的终末均体质量、摄食率显著高于LC 组(P<0.05)。

表3 苦瓜皂苷对松浦镜鲤幼鱼生长指标的影响Tab.3 Effect of bitter melon Momordica charantia saponins on growth of juvenile Songpu mirror carp in various treatments throughout the experiment
2.2 苦瓜皂苷对松浦镜鲤幼鱼肠道消化酶的影响
由表4 可知,8 周养殖试验结束时,HC 组鲤的蛋白酶及淀粉酶活性均低于LC 组,但并无显著差异(P>0.05),而脂肪酶活性显著高于LC 组。高糖饲料中添加苦瓜皂苷后,蛋白酶及淀粉酶活性先升高后降低,脂肪酶活性先降低后升高。MCS3 组鲤的蛋白酶及淀粉酶活性最高,显著高于HC 组和MCS4组(6 400 mg/kg)(P<0.05),而其脂肪酶活性最低,显著低于HC 组(P<0.05),且三种的活性均与LC 组无显著差异(P>0.05)。
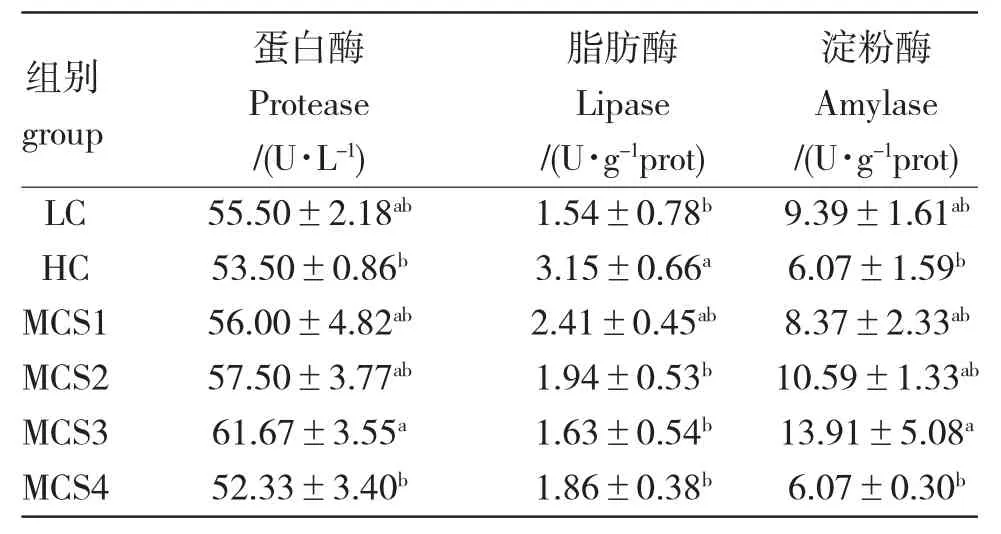
表4 苦瓜皂苷对松浦镜鲤幼鱼肠道消化酶的影响Tab.4 Effect of bitter melon M.charantia saponins on the intestinal digestive enzyme activities of juvenile Songpu mirror carp in various treatments throughout the experiment
2.3 苦瓜皂苷对松浦镜鲤幼鱼肝脏抗氧化指标的影响
由表5 可知,HC 组鲤的的SOD、CAT、POD、GSH 及T-AOC 活性均低于LC 组,其中POD 及T-AOC 活性显著低于LC 组(P<0.05),而MDA 含量却显著高于LC 组(P<0.05)。高糖饲料中添加苦瓜皂苷后,抗氧化酶活性先升高后降低,MDA 含量先降低后升高。MCS3 组的SOD、CAT、POD、GSH 及T-AOC 活性最高,显著高于HC 组(P<0.05),而其MDA 含量却最低,显著低于HC 组(P<0.05)。
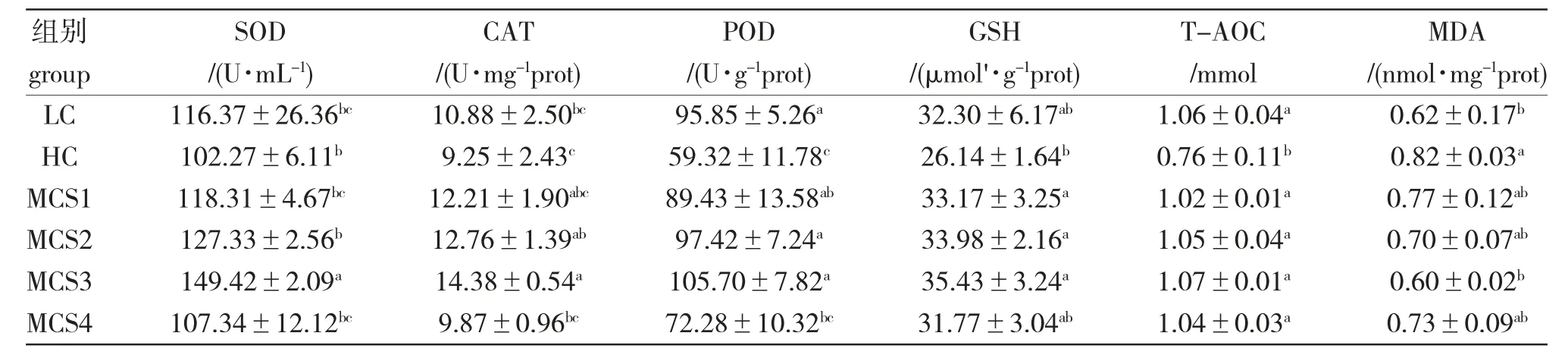
表5 苦瓜皂苷对松浦镜鲤幼鱼肝脏抗氧化指标的影响Tab.5 Effect of saponins of bitter melon M.charantia on the hepatopancreas antioxidant indices of juvenile Songpu mirror carp in various treatments throughout the experiment
2.4 苦瓜皂苷对松浦镜鲤幼鱼肝脏抗氧化相关基因表达的影响
由图1 可知,8 周养殖后,HC 组松浦镜鲤幼鱼中,除Keap1 外其余抗氧化相关基因的表达均显著低于LC 组(P<0.05)。随着高糖饲料中苦瓜皂苷含量的增加,松浦镜鲤幼鱼肝脏抗氧化相关基因的表达均先升高后降低。MCS3 组Nrf2、CuZnSOD、Mn-SOD 和CAT 的表达量最高,显著高于HC 组(P<0.05),MCS3 组(1 600 mg/kg)Keap1 表达量最低,显著低于HC 组,CAT 表达量显著高于LC 组(P<0.05)。
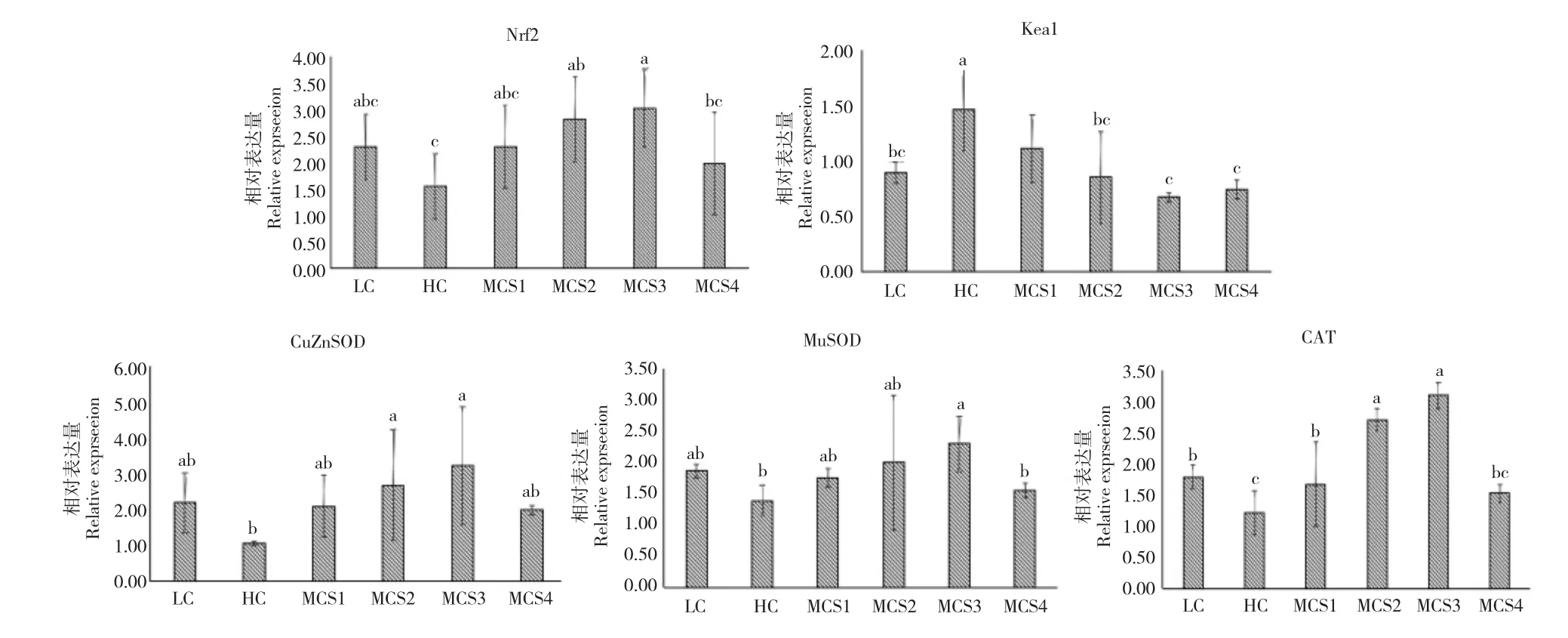
图1 苦瓜皂苷对松浦镜鲤幼鱼肝脏抗氧化相关基因表达的影响Fig.1 Effect of saponins of bitter melon M.charantia on the hepatopancreas antioxidant gene expression levels of juvenile Songpu mirror carp in various treatments throughout the experiment
2.5 苦瓜皂苷对松浦镜鲤幼鱼肝脏组织形态的影响
根据肝脏抗氧化酶活性及相关基因及其表达结果,对比了LC、HC、MCS3 及MCS4 组松浦镜鲤幼鱼肝脏组织形态学。与LC 组相比,HC 组会导致肝脏组织结构模糊,肝细胞严重损伤,出现细胞核缺失及严重脂肪样变;MCS3 组改变了HC 组的病理表现,肝细胞核更清晰,细胞界限明显且排列整齐,与LC 组肝脏组织结构完整度相似,结构较清晰。而在MCS4组则又出现了细胞核缺失,空泡化现象(图2)。
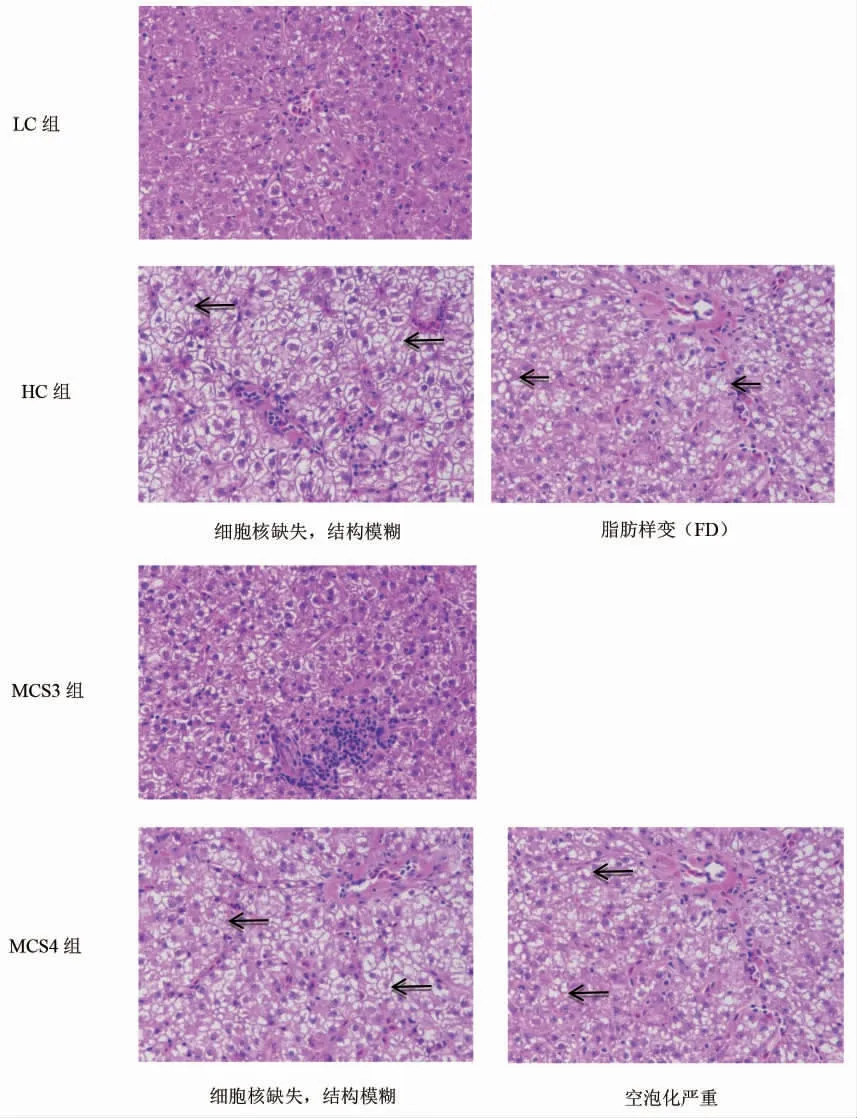
图2 苦瓜皂苷对松浦镜鲤幼鱼肝脏组织形态的影响(H.E 400×)Fig.2 Effect of saponins of bitter melon M.charantia on the hepatopancreas histology of juvenile Songpu mirror carp in various treatments throughout the experiment
3 讨论
饲料中添加适宜水平的糖类可促进鱼类生长,促进对高糖饲料的利用效率[17]。尽管杂食性鱼类对高糖饲料有较好的耐受能力[17],但过量添加依然会抑制鱼体生长[18]。本试验结果表明,高糖饲料组鱼的终末体质量与特定生长率低于低糖组,且高糖组鱼的饲料效率显著低于低糖组,表明高糖是限制鱼体生长的关键因素,这与许霄霄等[19]研究结果基本一致。本试验结果显示,在高糖饲料中添加1 600 mg/kg 苦瓜皂苷与高糖组相比,终末体质量、特定生长率及饲料效率均显著提高,上述指标也高于低糖组。目前对苦瓜皂苷的研究主要集中在人和小鼠上。陈松瑞[20]研究发现,灌胃350 mg/kg 苦瓜皂苷12周后,2 型糖尿病小鼠体质量显著回升,这在一定程度上佐证了苦瓜皂苷的促生长效应,但苦瓜皂苷在鱼类营养中的应用研究仍处于初期,需深入研究其促生长机制。
糖类应用于鱼体生长的首要限制因素即为消化。消化酶活性首先决定了鱼类对饲料营养物质的利用能力。鲤作为典型的无胃鱼类,对饲料营养物质消化吸收主要在肠道中[21]。有研究表明,在饲料的适宜范围内提高糖水平可增强淀粉酶、蛋白酶活性[22,23],而在本试验中高糖组(30%)肠道淀粉酶、蛋白酶活性显著低于低糖组,表明饲料糖水平可能超出了适宜范围,这与李晋南等利用50%高淀粉饲料饲喂松浦镜鲤的研究结果并不一致,可能是由于饲料糖源不同所致。本研究发现,当高糖饲料中补充1 600 mg/kg 苦瓜皂苷后,松浦镜鲤幼鱼肠道淀粉酶、蛋白酶活性较高糖组显著提高。Jr Serrano[24]研究发现,鲤摄食含皂树皂苷的饲料后肠道胰蛋白酶、淀粉酶活性明显提升。这揭示皂苷类物质在促进糖类、蛋白质等在鱼类肠道中的消化发挥重要作用,但具体调控机制有待于深入研究。
高血脂、高血糖是鱼类摄食高糖饲料后典型的应激症状,会导致进一步的组织氧化应激[25]。本研究发现,高糖饲料诱导了松浦镜鲤幼鱼肝脏抗氧化酶SOD、CAT、POD、GSH、T-AOC 活性的降低和脂质过氧化产物MDA 含量的升高,这在对异育银鲫(Carassius auratus gibelio)[4]、大口黑鲈(Micropterus salmoides)[26],白鲫(Carassius auratus cuvieri)[27]和彭泽鲫(Carussius auratus pengsenensis)[28]等研究中均有发现。相关研究发现,饲料中补充功能性添加剂能够缓解高糖饲料诱导的鱼类肝脏氧化应激[29]。已证实,苦瓜皂苷具有良好的抗氧化功效[12,30]。Jiang等[31]发现,苦瓜皂苷对高脂联合链脲菌素模型大鼠具有显著的抗氧化能力的恢复效应。在本研究中,高糖饲料中添加400~1 600 mg/kg 苦瓜皂苷增加了肝脏SOD、CAT、POD、GSH 和T-AOC 活性,减少了肝脏MDA 含量。刘波等[32]研究发现,苦瓜皂苷有良好的抗氧化活性,能够提高小鼠体内SOD 活性,降低非酒精性脂肪肝的发病风险。上述结果揭示苦瓜皂苷可有效缓解了高糖饲料引发的松浦镜鲤幼鱼肝脏抗氧化能力减弱,保护肝脏健康。在三疣梭子蟹(Portunus trituberculatus)的日常饲料基础中添加300 mg/kg 或450 mg/kg 皂树皂苷能增强其SOD、CAT 和谷胱甘肽过氧化物酶的活性[33]。结合本试验结果表明,皂苷类物质可作为改善水产动物抗氧化能力的功能性饲料添加剂。Keap1/Nrf2 信号通路是水产动物抵抗内、外源性氧化损伤关键信号通路[11,34]。本试验发现,高糖饲料中添加1 600 mg/kg 苦瓜皂苷显著增强了松浦镜鲤幼鱼肝脏Nrf2 及下游抗氧化酶(MnSOD、CuZnSOD 及CAT)的基因表达,而Keap1 基因表达被抑制,进一步揭示适宜水平苦瓜皂苷的添加能够减轻高糖饲料对鱼类肝脏的氧化应激损伤,发挥抗氧化剂的作用,且这种作用与Keap1-Nrf 2 信号通路密切相关。
高糖饲料会引起鱼体肝脏肿大,空泡化以及脂肪样病变[29,35],影响肝脏健康。李静辉[36]指出,鲤摄食含15%玉米淀粉的饲料时肝细胞出现水样变和轻微的脂肪样变;摄食含30%玉米淀粉饲料时肝细胞严重脂肪样变。本研究得出与之一致的结论。刘波等[32]发现,当苦瓜皂苷剂量为100 mg/kg 时,非酒精性脂肪性肝病大鼠肝细胞排列规则,结构完整,炎性细胞浸润与脂肪颗粒堆积基本消失。本研究同样发现,添加1 600 mg/kg 苦瓜皂苷后,松浦镜鲤幼鱼肝脏组织形态基本恢复正常,但添加量达到6 400 mg/kg 时,肝脏组织结构损伤加剧。这表明,苦瓜皂苷的添加剂量增加到1 600 mg/kg 能成为缓解高糖摄入诱导的代谢性肝病的有效手段,但组织修复机制需进一步深入研究。
综上,高糖饲料中补充适宜水平苦瓜皂苷可以增强松浦镜鲤幼鱼肠道对糖、蛋白质的消化吸收能力;减少对高糖的氧化应激反应,这一过程与Keap1-Nrf2 信号通路关系密切;同时也可修复高糖诱导的肝细胞受损。本研究发现松浦镜鲤高糖饲料中添加苦瓜皂苷添加水平应在1 600 mg/kg 左右,下一步将在此浓度上下游设计浓度点,以确定其最佳添加水平。当苦瓜皂苷添加水平达到6 400 mg/kg时,会对松浦镜鲤肠道消化及肝脏健康产生不利影响。
