碳氮投入对4种不同土地利用方式土壤磷含量及胞外酶活性的影响
2023-07-08赵媛媛靳嘉雯金予溦江佳艺郝祥蕊张德龙
赵媛媛,靳嘉雯,陈 硕,张 帅,金予溦,江佳艺,郝祥蕊,张德龙,陈 清
(1.中国农业大学 资源与环境学院,农田土壤污染防控与修复北京市重点实验室,北京 100193;2.上海农乐生物制品股份有限公司,上海 201419)
磷素是植物生长发育必需的大量营养元素之一。在农业种植体系中,由于化肥的不断施用,全球范围内土壤磷素累积水平均显著提升[1]。磷肥的不断施用在获得作物高产的同时,也造成土壤磷素大量累积而形成残留态磷,以及增加磷素向深层土壤或周围水体迁移的环境风险[2]。对于自然生态系统而言,存在普遍的磷素限制现象[3],这会导致不同土地利用的土壤磷素循环存在差异性。
在农田土壤中,由于大量养分的投入导致土壤磷素水平较高[4],高磷残留土壤的无机磷的比例相对较高,而有机磷比例相对较低,为9.6%~13.8%[5-6],高量有机肥的施用导致土壤的C/P、C/N或N/P比很低,周年种植造成土壤有机碳氮磷的矿化量很高。全球尺度下土壤和微生物生物量的C∶N∶P比值平均分别为186∶13∶1和60∶7∶1[7],或者为287∶17∶1和42∶6∶1[8],我国所有类型土壤的C∶N∶P比值平均为134∶9∶1[9]。而受高养分投入的影响,山东地区的部分设施菜田土壤的平均C∶N∶P比例只有25∶2∶1[10]。因此,土壤C/P也是影响土壤磷素转化及其对外源养分投入的响应存在差异的重要因素。
在高投入体系中,土壤有机碳成为微生物活性的主要限制因子,而外源有机碳的投入可以在一定程度上增加磷素生物有效性[11]。有机物料的施用为微生物提供了碳源和矿物质,提高了微生物和磷酸酶的活性,促进有机磷的转化。土壤中外源碳的投入将影响有机磷矿化、有机碳的降解以及微生物活性。有研究发现,有机碳的降解与有机磷化合物关系紧密。Spohn等[12]通过短期试验证实,有机磷矿化可能是微生物对碳源获取的副作用,微生物对碳源的需求能驱动微生物矿化葡萄糖-6-磷酸中的磷。在外加碳源的土壤培养试验中研究发现,加入蔗糖和玉米秸秆增加了胞外酶的分泌,包括β-葡萄糖苷酶、纤维二糖水解酶、几丁质酶、亮氨酸氨肽酶和木聚糖酶,而在其他研究中,还包括蛋白酶和磷酸酶[13]。Tomar等[14]的研究发现,土壤微生物呼吸与酶活性存在相关性,且与脱氢酶和酚氧化酶存在显著正相关,表明微生物生长和细胞外酶有良好对应的关系[15]。
氮肥投入影响了土壤有机碳的变化,增加了土壤微生物量,促进了土壤呼吸。适量施用氮肥还能加速磷酸酶酶促土壤有机磷化合物的水解,生成更多植物可利用的无机态磷,增强土壤磷的供应能力等[16]。研究发现,养分投入会影响磷在土壤—植物系统中的循环及周转,如影响微生物量磷的再矿化作用、非生命的磷的胞外酶水解作用及生物矿化过程等。外加氮源将影响土壤有机磷或磷素的形态转化,对土壤理化性质和酶的活性也会产生影响。适量施用氮肥能加速磷酸酶酶促土壤有机磷化合物的水解,生成更多植物可利用的无机态磷,增强土壤磷的供应能力[17]。除此之外,含氮元素的有机物的添加能有效影响土壤溶液中胞外酶的活性。如β-葡萄糖苷酶、纤维素二糖水解酶的活性随着有机物的添加有明显的提升[18]。
针对不同土地利用方式的土壤,其土壤磷水平以及碳磷比均存在较大的差异,外源碳氮投入会对土壤碳氮磷循环相关的胞外酶活性产生影响,进而影响磷含量;因此,本研究探究了4种土壤中外源碳氮添加对土壤胞外酶、速效磷和有机磷含量的影响,对合理、有效地预测农田土壤磷素累积状态、提高作物产量和保护生态环境具有重要的意义。
1 材料和方法
1.1 供试土壤
试验所采用的4种土壤为:设施菜田土壤、粮田土壤、草原土壤和森林土壤,设施菜田土壤采自北京房山区西场村金地汇农农业合作社设施菜田,于收获期采集,该地气候类型为典型的大陆性季风气候,年平均气温11.6 ℃,降水量603 mm,种植过程中的鸡粪和尿素用量分别为3 t/hm2和150 kg/hm2,种植农作物为番茄;粮田土壤采自设施菜地附近的土壤,于玉米收获期采集;草原土壤采自内蒙古草原生态研究站,位于内蒙古多伦县,海拔1 324 m(42°02′N,116°17′E),原生植被是一个有羊草和冰草的典型草原社区;森林土壤采自北京森林生态系统定位研究站,位于北京西郊门头沟区东灵山,海拔1 100 m(115°36′E,39°48′N),森林包括一些典型的温带地区次生落叶阔叶林和人工针叶林。土壤采集后过筛(2 mm)风干,去除土壤中的可见植物残体和土壤动物,待培养。试验前测得4种土壤的理化性质如表1。

表1 土壤基本物理性质Tab.1 Basic physical properties of soil
1.2 试验设计
试验以葡萄糖作为碳源,以硝酸钾作为氮源,共设4个处理,分别为对照(CK)、葡萄糖(G,500 mg/kg土壤)、硝酸钾(N,300 mg/kg土壤)、葡萄糖和硝酸钾(GN,葡萄糖500 mg/kg土壤和硝酸钾300 mg/kg土壤)。碳氮添加量以风干土质量计。首先进行预培养:即将4种风干过筛(2 mm)的供试土壤分别称取12份100 g于250 mL玻璃培养瓶中,加水至土壤最大田间持水量的60%,覆盖带孔透气的封口,放入恒温培养箱(25 ℃,湿度60%)中进行为期7 d的预培养,使土壤趋于稳定状态。然后进行正式培养:根据处理分别称取葡萄糖和/或硝酸钾,与土壤充分混匀,再次置于恒温培养箱中,每个处理3次重复,随机排列,培养期间用称质量法补充水分。
1.3 测试项目与方法
1.3.1 气体采集 在培养的第1,2,5,10,18,55天进行气体(CO2)采集。采集时,将土壤从培养箱中取出,将培养瓶取出进行通风放置10 min,然后用封口膜密封在常温暗处静置2 h,分别在封口的前后各取20 mL气体,储存于气袋中,然后再次将培养瓶放回培养箱内培养。气体用气相色谱仪(Agilent 7890A,美国)进行测定,分析CO2前后浓度的差异及其与所加物料的关系。
1.3.2 土壤样品采集与测定 培养的第55天取气后采集土壤样品,将罐子中所有土壤取出并分为2份:其中1份为称取20 g置于4 ℃保存,于7 d内测定磷酸酶的活性;其余风干,测定相关理化性质(土壤pH值、土壤有机碳、土壤总氮、土壤总磷、土壤有机磷、速效磷)。土壤pH值利用水提取土壤溶液(土水比1.0∶2.5)用酸度计测定。土壤总磷(TP)通过浓硫酸和H2O2消煮法测定。土壤有机磷利用灼烧法测定[19]。土壤Olsen-P采用0.5 mol/L NaHCO3(pH=8.5)溶液提取(土水比1∶20),并用钼锑抗比色法测定[20]。土壤总碳(TC)、总氮(TN)和有机碳(OC)含量通过干烧法,利用元素分析仪测定(Vario Micro Cube,Elementar)。土壤酶活性通过荧光微型板酶检测方法进行测定,同时调节缓冲液的不同pH值(Turner,2010),碱性磷酸酶(ALP,EC 3.1.3.1)以4-甲基伞形基磷酸为底物,配制缓冲溶液pH值为11.0,葡萄糖苷酶(BG,EC 3.2.1.21)以4-甲基伞形酮基β-D-葡萄糖苷为底物,配制缓冲溶液pH值为6.0,最后在激发波长为365 nm、发射波长为450 nm的条件下用多功能酶标仪(SpectraMax i3x)读取荧光值,所有试剂均采购于Sigma Aldrich公司。
1.4 数据分析
试验数据以Excel 2007软件整理,采用SPSS软件进行统计分析,用LSD法进行显著性检验,显著性水平P<0.05,并进行相关分析,得出相关值后评价。
2 结果与分析
2.1 碳氮投入对土壤磷含量的影响
2.1.1 土壤有机磷含量 4种土壤中有机磷的含量对碳氮不同投入响应不同(图1),且不同土地利用方式下,土壤有机磷含量差异大。结果表明,与CK处理相比,添加碳源显著提高了菜田、森林土壤有机磷的含量,分别提高了89.7%,40.6%;添加碳源、添加氮源和同时添加碳源、氮源均显著提高了森林土壤有机磷含量,分别提高了40.6%,33.6%,46.8%,且同时添加碳源和氮源时,土壤有机磷含量最高。

不同小写字母代表同一类型土壤中不同处理间达到显著差异水平(P<0.05)。A.菜田;B.粮田;C.草地;D.森林。图2—5同。Different lowercase letters represent significant differences between different treatments in the same type of soil(P<0.05).A.Greenhouse vegetable fields;B.Grain field;C.Grassland;D.Forest.The same as Fig.2—5.
2.1.2 土壤速效磷含量 4种土壤中速效磷的含量对碳氮不同投入响应不同(图2),且不同土地利用方式下,土壤速效磷含量差异明显。与CK处理比较,N处理菜田的土壤速效磷显著增加了14.2%。其余3种土壤在添加碳源、添加氮源以及同时添加碳源、氮源后,土壤速效磷含量均有下降,其中森林土壤在碳氮投入后下降最明显,3个处理组中土壤速效磷分别下降了14.5%,14.0%,9.4%。

图2 不同处理对不同土地利用方式的土壤有效磷含量的影响Fig.2 Effects of different treatments on soil olsen-P content under different land use patterns
2.2 碳氮投入对土壤胞外酶活性的影响
2.2.1 土壤碱性磷酸酶活性 4种土壤中ALP的活性对碳氮不同投入响应不同(图3)。结果表明,与CK处理相比,碳氮添加处理对菜田和粮田土壤ALP活性没有显著影响,单独碳源添加处理显著降低了草地和森林土壤的ALP活性,分别降低了35.9%,25.5%,与此同时,氮添加也显著降低了草地和森林土壤的ALP活性,分别降低了28.8%,24.8%。草地和森林土壤在添加碳源的基础上添加氮源后与仅添加碳源或氮源相比,均提高了土壤ALP活性,这种变化在森林土壤中更为显著。

图3 不同处理对不同土地利用方式的土壤碱性磷酸酶活性的影响Fig.3 Effects of different treatments on soil alkaline phosphatase activity under different land use
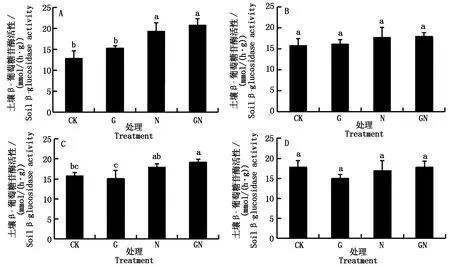
图4 不同处理对不同土地利用方式的土壤β-葡萄糖苷酶活性的影响Fig.4 Effects of different treatments on soil β-glucosidase activity under different land use
2.2.2 土壤β-葡萄糖苷酶活性 4种不同处理方式下,不同类型土壤β-葡萄糖苷酶活性对碳氮投入的响应不同(图 4)。结果表明,与CK处理相比,氮源添加显著提高了菜田土壤的β-葡萄糖苷酶活性,提高了50.7%。碳、氮同时添加处理对于4种土壤的β-葡萄糖苷酶活性均有增加,且菜田和草地土壤中这种差异更为显著,分别增加了62.6%,21.6%。且碳氮添加与仅添加碳源或氮源相比,均提高了土壤β-葡萄糖苷酶活性,这种变化在菜田和草地土壤中更为显著。
2.3 土壤理化性质
土壤培养55 d后,可观察到不同处理的土壤理化性质发生了改变(表2)。添加氮源后,菜田、粮田土壤pH值均显著降低,而在草地土壤pH值则显著增加,但森林土壤pH值无显著变化。不同土地利用方式下,添加氮肥处理均增加了土壤全氮含量,其中以菜田、草地和森林土壤最为显著。氮源和碳源的添加对全磷和OC/TP则没有显著影响。已有文献中证明,pH值的改变会直接或者间接影响土壤磷素有效性,然而本研究中虽然pH值发生显著变化,但是其增幅和降幅均在0~7.96%,加之土壤本身的缓冲性,不足以引起土壤磷素有效性的改变。因此,可以认为本试验中速效磷含量的变化来源于氮肥本身。

表2 碳氮投入对不同土壤理化性质的影响Tab.2 Effects of carbon and nitrogen inputs on physicochemical properties of different soils
2.4 碳氮投入对土壤CO2释放量的影响
在持续55 d土壤培养过程中,第1,2,5,11,18,55天采得气体的累积排放总量见图5。不同土地利用方式下,土壤微生物呼吸量相对于对照均无显著差异或有增加。不添加有机物料的处理中,菜田和粮田的土壤微生物呼吸量均比草地和森林的土壤微生物呼吸量低。在4种处理中,添加了碳源的土壤微生物呼吸量与对照相比显著增加,添加了氮源的土壤微生物呼吸量与对照相比变化不显著。在草地土壤(TN=1.97 g/kg)及森林土壤(TN=2.99 g/kg)的高氮条件下,在碳源添加的基础上,外加氮源明显抑制了土壤微生物呼吸,CO2累积排放量分别降低了19.7%和16.5%,而在菜田、粮田土壤中未观察到显著差异。

图5 4种处理条件下4种土壤CO2累计释放量Fig.5 Four kinds of soil CO2 cumulative release under four kinds of treatment conditions
3 结论与讨论
3.1 碳氮投入对不同土地利用方式的土壤磷含量变化的影响
与森林土壤对比,菜田、粮田和草地3种土壤的C/N较低,碳为相对限制因素,氮源的添加可能通过改变微生物的群落结构进而改变微生物对碳源的利用,受到碳限制的微生物会使用磷酸化化合物的有机部分作为碳源[21]。许多近年来文献结果表明,外源碳氮的添加会导致土壤微生物的呼吸量增加引发起爆效应[22-25]。一般而言,这个过程会改变土壤有机碳的分解[26],而对于外源添加不稳定或者活性的碳源等物质,则会通过起爆效应的正效应增加土壤有机碳的分解[18,27],第一种是表观分解,即微生物只是利用了外源投入的能源物质,而并未分解土壤中有机碳,这种情况下,可能并不会影响到土壤中养分的循环过程;第二种是实际的分解,即土壤微生物利用外源能源物质的同时,也分解了土壤中的有机碳,则会显著改变土壤中其他元素的转化过程[28],而这可以在一定程度上驱动土壤磷素转化过程。有研究表明,添加葡萄糖和纤维素显著促进了土壤微生物的生长以及对土壤磷素的活化[29],通过施用外源碳促进土壤微生物对磷素的固持进而降低其在土壤剖面的迁移[30]。本研究结果显示,与CK相比,添加葡萄糖处理显著降低了草地和森林土壤速效磷含量,可能是由于碳源加入驱动了微生物对磷的固定;而对于速效磷含量本就较高的设施菜田而言,葡萄糖添加并未显著影响其土壤速效磷含量,但是显著增加了有机磷含量,因此,土壤碳磷循环是存在耦合关系的。对于氮素添加后,可能导致磷酸酶的活性受到抑制,从而阻碍了有机磷的矿化进程,因此,添加氮源的处理比对照处理的有机磷含量更高。这与于群英[17]的研究一致,适量施用氮肥能加速磷酸酶促进土壤有机磷化合物的水解,生成更多植物可利用的无机态磷,会增强土壤磷的供应能力。
有研究表明,土壤有机质和pH值是影响土壤磷素转化的关键因素,需要指出的是,对于氮肥而言,其长期效应会导致土壤酸化,这会分别通过生物过程和非生物过程进而影响土壤磷素转化,本试验尽管氮源添加显著影响了土壤pH值,但并未因此显著影响土壤磷素含量。本研究结果表明,外源碳的添加均显著增加了培养初期土壤的呼吸量,在培养的第7天趋于平稳,与此同时,碳氮共同添加的处理显著低于单施碳源处理,即硝酸铵抑制了土壤CO2的释放量,已有研究结果表明,在添加葡萄糖的同时添加硝态氮或者铵态氮均会抑制起爆效应[31],土壤有机碳和微生物生物量之间存在正相关关系[30],葡萄糖这一类活性的碳源添加到土壤中后,会增加微生物的生长或者改变微生物的群落结构,而随着微生物的生长,会逐渐消耗对土壤无机氮的利用进而降低氮的含量,此时微生物会通过分泌相关的酶分解土壤中的有机碳进而获取用于自身生长的元素[32]。然而,当碳氮一起添加时,氮源则可以直接被微生物利用,且土壤微生物不必通过分泌酶去分解有机碳[26,33],因此,会降低起爆效应[34],有研究表明,碳源的添加可使土壤速效磷含量降低而微生物量磷增加,降低活性磷含量以及潜在的磷素淋失风险[30],也有研究指出,添加葡萄糖提高了土壤微生物活性、土壤呼吸、微生物量磷和磷酸酶活性,短期内添加葡萄糖使得土壤微生物大量增殖消耗土壤有效磷,但长期来看并不会影响土壤有效磷含量[35]。
3.2 碳氮投入对不同土地利用方式土壤胞外酶活性的影响
土壤酶的活性在土壤肥力的评价中起着重要作用[36]。酶是土壤有机磷矿化作用中的重要催化剂,主要来源于植物根系和微生物代谢物,以及由其他动植物分解过程中产生。提高土壤碱性磷酸酶的活性,可促进土壤有机磷矿化进而满足作物或微生物对磷素的需求[37]。土壤磷酸酶酶促作用可以表征土壤磷素有效化强度[38]。在本试验中,草地和森林土壤的氮素含量较高,而农田土壤的却较低,可能由于本研究中的设施菜田和粮田土壤种植年限均较短,且设施菜田土壤质地为沙壤土,夏季揭膜过程中氮素发生了淋失,而草地和森林土壤可能是长期氮沉降导致。结果表明,在设施菜田土壤中,外源碳源的添加对β-葡萄糖苷酶活性没有显著影响,但是氮源添加显著增加了该土壤β-葡萄糖苷酶活性,与此同时,碳氮同时添加显著增加了草地土壤β-葡萄糖苷酶活性,因此,在本试验的设施菜田土壤中添加氮源有利于碳的循环过程,而草地中由于氮素含量较高,在补充碳源基础上,氮源添加有助于提升其β-葡萄糖苷酶活性。在此过程中,土壤微生物通过分泌与碳磷循环相关的胞外酶,以平衡元素之间的化学计量关系[39],进而影响土壤磷素含量。有研究表明,适量施用氮肥会加速磷酸酶促进土壤有机磷化合物的水解,生成更多植物可利用的无机态磷,增强土壤磷的供应能力[16]。而对于森林土壤而言,则是相反的过程,即由于ALP活性的降低抑制了有机磷的矿化,进而无机磷的释放量随之降低。菜田土壤中OC/TP相对较低,在碳源添加的基础上,氮源的添加会促进有机碳的降解,进而影响到有机磷的含量,而这种作用可能来源于β-葡萄糖苷酶活性的增加。有研究结果表明,长期氮肥施用显著抑制了ALP活性,而这可能通过降低土壤有机磷的矿化进而增加土壤有机磷的累积[5]。也有研究表明,氮肥添加显著提升了土壤ALP的活性,这可能是由于在磷素缺乏的情况下,氮肥施用促进了根系或土壤微生物对磷酸酶的分泌[40],并促进了作物对无机磷的吸收,同时间接增加了土壤有机磷的比例。Chen等[18]通过向土壤中添加蔗糖后,发现显著增加了土壤胞外酶,包括β-葡萄糖苷酶活性,此过程也增加了土壤磷酸酶活性,而这种变化是由于增加了土壤微生物的生长导致的[41]。然而,针对碳氮添加如何通过调控土壤碳氮磷化学计量以及酶的化学计量进而影响土壤磷素形态转化的机制还需要进一步探讨,这可以为理解不同养分含量土壤磷素调控和管理提供理论参考。
碳源添加显著增加了菜田土壤有机磷含量;同时添加碳氮显著提升了森林土壤有机磷含量;碳源添加显著降低了草地土壤Olsen-P含量;添加氮源显著增加了菜田土壤的Olsen-P含量,降低了森林土壤的Olsen-P含量。
碳氮投入对菜田和粮田土壤碱性磷酸酶没有显著影响,但氮素添加提高了菜田土壤β-葡萄糖苷酶活性;在草地和森林土壤中,氮素投入抑制了碱性磷酸酶活性,而在碳源基础上添加氮素则提高了碱性磷酸酶和β-葡萄糖苷酶活性。
不添加碳源条件下,氮素添加对土壤CO2释放量没有影响,添加碳源后,菜田土壤有增加CO2释放的趋势,同时,氮素抑制了高碳体系土壤CO2的释放量。
综上所述,不同土地利用方式的土壤磷素含量和酶活性对碳氮添加的响应存在差异性,通过外源添加碳源可以一定程度上通过调控微生物对酶的分泌,实现“以碳促磷”,进而增加土壤磷素供应能力。
