我国主要玉米病毒病的鉴定及分类研究进展
2023-06-04顾天潇冯陈尉郭枭花滟泓贺振张坤陈佳欢
顾天潇 冯陈尉 郭枭 花滟泓 贺振 张坤 陈佳欢

摘要:玉米是重要的粮食作物,和人们的生活息息相关,但病害种类复杂,其中病毒病害存在极大威胁,可能会导致玉米产量损失惨重,当前病毒病的有效防治手段主要是利用高抗性品种,但该技术耗时费力,且依赖性高;在田间及时发现病毒病,尽早处理田间病株,是控制病毒病发生的有效手段,因此准确地识别玉米病毒病害,尽早进行检测鉴定并做出相关防治措施,可以最大程度降低损失。本文归纳了我国玉米上的主要病毒病害,主要包括引发国内玉米粗缩病、玉米矮花叶病、玉米红叶病、玉米鼠耳病以及玉米褪绿斑驳病的病原,归纳国内侵染玉米并造成危害的主要病毒,如水稻黑条矮缩病毒(RBSDV)、玉米矮花叶病毒 (MDMV)、大麦黄矮病毒(BYDV)、黄瓜花叶病毒(CMV)等病毒,简述这些病毒的传播介体及主要传播途径,从源头上为预防玉米病毒病提供参考;同时整理相关病毒的基本信息,如病毒分类、病毒粒子特性、病毒基因组结构、病毒侵染植物后引发的症状等;旨在加深人们对生产上常见的玉米病毒病的认识和理解,以期为准确快速识别玉米病毒病提供参考,为田间防控提供理论支持。
关键词:玉米病毒病;症状;病毒粒子;基因组结构;介体昆虫;传播途径;病害防治
中图分类号:S435.131.4+9 文献标志码:A 文章编号:1002-1302(2023)09-0001-08
基金项目:国家自然科学基金(编号:81903764、31801699);江苏省自然科学基金优秀青年基金项目(编号:BK20220116);江苏省自然科学基金青年项目(编号:BK20180904);广东省植物保护新技术重点实验室开放课题(编号:植重2021-07);江苏省研究生科研与实践创新计划(编号:SJCX22_1792)。
作者简介:顾天潇(1999—),男,甘肃张掖人,硕士研究生,从事植物病毒学研究。E-mail:984520812@qq.com。
通信作者:陈佳欢,博士,讲师,硕士生导师,从事药用植物天然抗菌活性成分挖掘研究,E-mail:092015@yzu.edu.cn;张 坤,博士,副教授,硕士生导师,从事植物病毒学研究,E-mail:zk@yzu.edu.cn。
玉米(Zea mays L.)属禾本科玉蜀黍属,是我国极为重要的粮食作物。我国玉米生产量占世界的 20%,是全球第二大玉米生產和消费国[1]。在我国,玉米总产量仅低于水稻和小麦,是重要的粮食来源,同时也是工业原料(工业发酵等)和药品成分的来源,与农业、畜牧业联系密切[2]。
玉米耐旱、耐寒,其环境适应性很强,在贫瘠土地上生长状况良好[3]。但在玉米生长过程中,会遭受多种病原物侵害,继而引发各种病害,导致玉米产量或者品质下降。玉米病害的发生受多方面因素影响,如病原物特性、病原物的类型、植物生长状况、环境条件等。造成玉米病害的病原物十分繁杂,包括真菌、细菌、线虫和病毒等。迄今为止,我国已发现并报道了40余种玉米病害[4],常见病害有30余种[5]。病毒性病害对玉米的生产造成了极大的危害,通常被称为玉米“癌症”[6]。在实际生产中,玉米病毒病的田间鉴定是及时发现病害类型并挽回经济损失的关键,但由于不同病毒会引起相似的症状,同时单一玉米植株上可能侵染多种病毒等情况,因此对于玉米病毒病的准确识别还存在难点。本文深入总结了常见玉米病毒病的病原特性,症状、病毒分类地位、病毒基因组情况、传播侵染途径、病毒蛋白的编码特征等,这有助于加深对病毒病的认识,也有利于田间一线生产人员尽早精准识别玉米病毒病害,及时采取相关的防控策略,避免病毒病害的大流行大暴发,为实际生产中玉米病的识别及防控提供理论参考。
1 呼肠孤病毒科
1.1 水稻黑条矮缩病毒
1.1.1 分类地位及寄主范围、分布范围 水稻黑条矮缩病毒(RBSDV)属于呼肠孤病毒科斐济病毒属1组[7],可侵染多种禾本科植物,RBSDV的自然寄主有水稻、小麦、玉米等作物[8]。在我国,RBSDV是玉米粗缩病的主要病原[9-11],主要分布于我国北方的黄淮海夏玉米主产区[12-14],可引发玉米粗缩病。
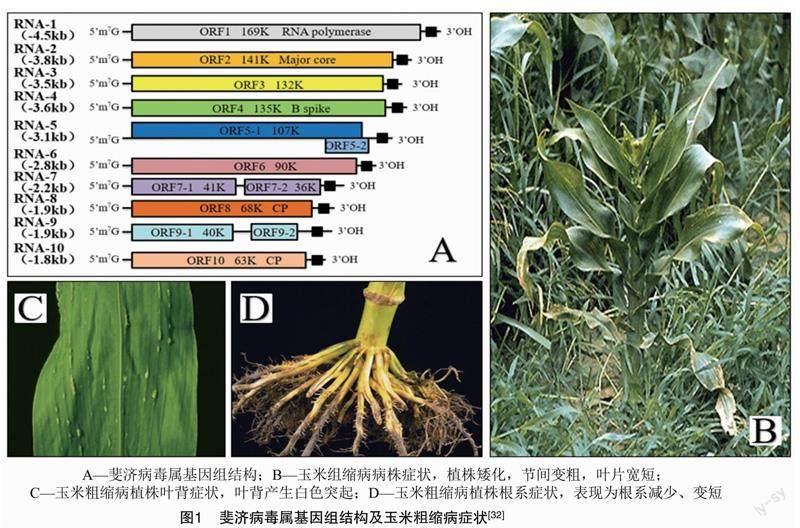
1.1.2 病毒特性 RBSDV的病毒粒子为双层外壳的正二十面体,直径为70~80 nm[15],其基因组包括10条RNA(S1~S10),大小为29 142 nts。RBSDV基因组大多只编码1个蛋白,少数基因组片段编码2个蛋白,如S5、S7和S9[16]。S1编码RNA聚合酶;S2编码毒粒子的核心蛋白;S3编码结构蛋白;S4编码B刺突结构蛋白;S5编码2个蛋白,功能尚不明晰;S6编码的蛋白参与病毒复制;S7编码2个蛋白,前者参与形成管状结构,后者功能未知;S8编码外壳蛋白;S9编码2个蛋白,40 ku蛋白参与病毒的包装形成,24 ku蛋白尚无检测报道[17];S10编码病毒粒子外层外壳蛋白。该病毒基因组结构见图1-A。
1.1.3 侵染途径与症状 RBSDV在田间的主要传毒介体是灰飞虱,带毒的灰飞虱在生命期内均能传毒,但不能通过卵传播[18]。受病毒侵染的植株严重矮化,植株节间变短、变粗,病株分蘖多、根系减少,在土壤中扎根变浅且易拔出;叶片的典型症状是顶叶簇生,心叶卷曲变小;影响抽穗结实,产量严重下降;叶背有白色突起,手感粗糙[19],植株表现如图1所示。
1.2 南方水稻黑条矮缩病毒
1.2.1 分类地位及寄主范围 南方水稻黑条矮缩病毒(SRBSDV)属于呼肠孤病毒科斐济病毒属2组,与玉米粗缩病毒(MRDV)亲缘关系最近,都是斐济病毒属2组成员[20];该病原最早被认作RBSDV的新株系[21],后续发现该病毒的媒介昆虫不是灰飞虱,而是白背飞虱,该病毒基因组与RBSDV基因组存在明显差异,最终被确定为SRBSDV[20-23];国际病毒分类命名委员会在2016年认定该病毒为确定种[24]。该病毒自然寄主为玉米、水稻、小麦等禾本科作物[25],主要分布于我国中部及南部[26],最早于2008年在山东济宁玉米上被发现,并引发玉米粗缩病[27]。
1.2.2 病毒特性 SRBSDV的基因组由10条dsRNA组成,含29 124个碱基,球状病毒粒体(直径70~75 nm),仅在感病植株韧皮部聚集,在寄主细胞内成晶格状结构[24]。病毒基因组共有10个片段,命名为S1~S10[22-23],共编码13个蛋白,其中S1~S4、S6、S8、S10各编码1个蛋白,S5、S7、S9分别编码2个蛋白。结构蛋白分别为 P1、P2、P3、P4、P8和P10[28],可参考图1。
1.2.3 侵染途径及症状 自然条件下白背飞虱高效传播SRBSDV[29],该病毒在白背飞虱体内可增殖,携带该病毒的白背飞虱终生带毒,但无法卵传毒。所有龄期均能传毒,与成虫相比,若虫传毒效率更高,病毒在玉米和白背飞虱中的循环主要发生于玉米苗期[30-31]。植株受SRBSDV侵染后的典型症状是严重矮化、节间缩短,叶片变宽变短,根系发育迟缓,雌雄穗畸形或不发育,玉米整个生育期均可被该病毒侵染,苗期对病毒最敏感,植株后期抗病性增强,发病减轻。
2 马铃薯Y病毒科
2.1 玉米矮花叶病毒
2.1.1 分类地位及寄主范围 玉米矮花叶病毒 (MDMV)属于马铃薯Y病毒科(Potyviridae)马铃薯Y病毒属(Potyvirus)。最早于1968年在我国河南省新乡市发现,并于1972年确定病原物为MDMV。MDMV的寄主范围较窄,仅侵染部分禾本科作物,如玉米、甘蔗、高粱等[33]。
2.1.2 病毒特性 MDMV為正义单链RNA病毒,其基因组结构为典型的马铃薯Y病毒属结构(图 2-A),无膜包被的弯曲线状病毒粒子[34],分子大为(430~750) nm×(12~15) nm[35];分子量为(3.0~3.5)×103 ku;病毒粒子中核酸占5%左右,其他部分由蛋白组成。该病毒的致死温度为55~60 ℃,在室温下能保持毒力1~2 d,在低温条件下数年后仍具有侵染力。该病毒基因组编码的多聚蛋白在水解酶的切割下分为10个功能不同的蛋白;P1-Pro蛋白、HC-Pro(辅助成分蛋白酶)、P3蛋白、6K1蛋白、CI蛋白(柱状内含体蛋白)、6K2蛋白、VPg蛋白(基因组连接功能)、NIa蛋白(核内含体A)、NIB蛋白(核内含体B)、CP蛋白(外壳蛋白)。不同株系中P1蛋白差异较大,NIB蛋白较为保守[36]。
2.1.3 侵染途径及症状 玉米矮花叶病毒通过侵染在越冬性禾本科植物上成为次年的初侵染源(图2)。病毒侵染植株后,前期表现为叶片黄化坏死,后期出现植株矮化、无法抽穗、果穗结实不良等症状[37]。病害具备暴发性、迁移性、间歇性的特点,是玉米的主要病害之一[38]。自然环境下病毒主要通过蚜虫传毒[39],也可通过种子传毒。
2.2 甘蔗花叶病毒
2.2.1 分类地位及寄主范围 甘蔗花叶病毒 (SCMV)属于马铃薯Y病毒科Y病毒属。病毒寄主为禾本科植物,主要侵染甘蔗、玉米、高粱[40]。目前已报道证实,我国玉米矮花叶病主要病原是SCMV[41-42],该病毒在全国玉米主产区内均有分布。
2.2.2 病毒特性 SCMV为正义单链RNA(大小为9 000~12 000个核苷酸),线性病毒粒子,长 680~900 nm[43]。病毒基因组编码的多聚蛋白经蛋白酶切割加工成10个蛋白,分别为P1-Pro蛋白、HC-Pro蛋白(辅助成分蛋白酶)、CI蛋白、P3蛋白、VPg蛋白、NIa蛋白(核内含体蛋白a)、NIb蛋白(核内含体蛋白b)、 CP蛋白、6K1蛋白、6K2蛋白[44],基因组结构见图2。
2.2.3 侵染途径及症状 SCMV侵染玉米后,叶片出现浅黄色或浅绿色,有黄绿相间的不规则的嵌纹或条纹(图2)。长短不一、布满叶片,新叶症状表现最为明显,同时伴随植株矮化。该病毒通过蚜虫传播或人工摩擦传播[45]。除此之外,也有相关报道指出SCMV通过种子传播[46]。
2.3 高粱花叶病毒
高粱花叶病毒(SrMV)属于马铃薯Y病毒科Y病毒属。自然寄主局限于禾本科植物,田间主要通过蚜虫传播,其经常与SCMV复合侵染[47]。自然条件下未见该病毒侵染玉米的情况,但人工接种条件下可侵染玉米[48]。基因组结构与马铃薯Y病毒属内其他病毒基因组结构相似(图2)。
2.4 白草花叶病毒
2.4.1 分类地位及寄主范围 白草花叶病毒(PenMV)属于马铃薯Y病毒科马铃薯Y病毒属SCMV亚组。早期被认为是MDMV的不同株系,于1986年被分离鉴定,2003年被证实为该属内新病毒,并将之命名为PenMV[49]。病毒主要侵染玉米、高粱、白草等禾本科作物和杂草,侵染玉米可引发矮花叶病[50]。我国首次于2003年在山西省忻州市发现该病毒自然侵染玉米的情况[51]。
2.4.2 病毒特性 PenMV为线性病毒粒子,植物组织内可见病毒形成的风轮状、柱状、卷筒状、板状等内含体;该病毒钝化温度为53 ℃,体外可存活 48 h,具备病毒紫外吸收特性曲线[52],病毒基因组结构为马铃薯Y病毒属的经典结构(图2)。
2.4.3 侵染途径及症状 该病毒主要通过蚜虫传播[50],受PenMV侵染的植株表现为系统性的花叶,叶脉间有花叶条纹,且深浅相同,幼叶上症状尤为明显,病原物可扩展到整棵植株。
3 黄症病毒科
3.1 大麦黄矮病毒
3.1.1 分类地位及寄主范围 大麦黄矮病毒(BYDV)是引发玉米红叶病的病原,属于黄症病毒
科(Luteovirdae)的病毒。在我国,引发玉米红叶病的BYDV分为GAV、GPV、PAV和RMV等4种株系[55],其中GAV株系和PAV株系属于黄症病毒属(Luteovirus),而PAV和RMV株系尚无明确分类地位[56],经过相关学者对其生物学信息的分析,推测PAV和RMV株系属于马铃薯卷叶病毒属(Polerovirus)[57],中国最早于1984年在河南地区发现该病害[58],该病毒主要侵染禾本科植物。
3.1.2 病毒特性 黄症病毒科病毒均为线性的正义单链RNA。BYDV病毒粒体形态呈正十二面体,无膜包被的对称球形,立体直径为25~30 nm。病毒各个株系之间基因组大小都约为5.7 kB,没有帽子结构和Poly(A)结构[59]。基因组结构如图3-A所示。
基因组内共编码6个蛋白,分别为P1(功能尚不明确)、P2(RNA聚合酶,P2蛋白不可单独表达,需与P1蛋白融合表达)、P3(外壳蛋白CP)[60]、P4(病毒运动相关蛋白MP)、P5(与蚜虫传播密切相关)[61]、P6(功能未知)。
3.1.3 传播途径及症状 玉米红叶病有效的传毒介体为蚜虫(多种蚜虫如玉米蚜、麦二叉蚜等)。植株感病后主要的症状为叶片褪绿黄化或红化,病叶质地硬,无卷曲,叶片发亮,严重时叶片干枯(图3)。发病越早症状越明显,病后期叶片出现红色条斑,也会产生细条症状[62]。
4 黄瓜花叶病毒
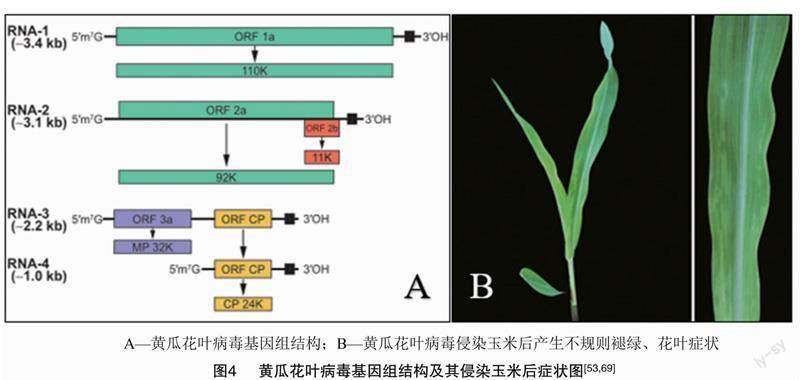
黄瓜花叶病毒(CMV)属于雀麦花叶病毒科(Bromoviridae) 黄瓜花叶病毒属(Cucumovirus),可侵染100多个科1 200多种单、双子叶植物,是寄主植物最多、分布最广、经济重要性最高的植物病毒之一[64]。我国已有报道显示该病毒可在自然条件下侵染玉米并造成损失[65],侵染玉米后导致植株出现不规则条纹花叶、矮化、斑驳等症状[66](图4)。自然界内通过蚜虫持续传毒。
CMV病毒粒子呈二十面体类球体。基因组包含3条正单链RNA,每个RNA片段的5′端为甲基化帽子结构,RNA 片段的3′端均有约200个核苷酸的高度保守区域,3′端无Poly(A)結构,有tRNA状结构,能结合酪氨酸[67],基因组结构如图4所示。
RNA1编码la蛋白(解旋酶和甲基转移酶);RNA2编码2a蛋白(RNA聚合酶),与la蛋白协同作用形成复制复合体[67]。此外,RNA2还编码产生2b蛋白:一种多功能蛋白,与病毒症状的产生、RNA沉默的抑制和病毒的长距离移动有关[68]。RNA3编码3a蛋白(运动蛋白MP);RNA3的亚基因组RNA4,编码产生CMV的外壳蛋白(CP)(参与病毒粒子的包装、病毒的蚜虫传播和长距离移动)[68]。
5 玉米鼠耳病毒
玉米鼠耳病由玉米鼠耳病毒(MWEV)侵染所致,最早于1988年出现在四川省南充市[70],之后主要集中发生于重庆、四川和贵州等地[71]。该病毒目前尚无明确的分类地位,但根据之前的相关研究,认为其有可能是斐济病毒属的一个成员。MWEV的主要寄主为玉米、水稻、大麦等禾本科作物及杂草。
MWEV病毒粒子是一种具有双层衣壳的dsRNA球状病毒,衣壳形态与斐济病毒(FDV)形态极其相似。基因组由10条dsRNA片段组成,关于其基因组其他研究尚处于空白状态[72]。
玉米鼠耳病毒可通过叶蝉传播,无法通过种子、土壤、机械方法或其他昆虫传播,在我国主要传毒昆虫为二点叶蝉和斑翅二室叶蝉[73],有报道称MWEV可通过叶蝉的卵进行传播[74-75]。
MWEV侵染玉米后,植株矮缩,节间缩短,叶片缩小加厚,叶缘内卷,心叶卷缩不能展开,呈“火炬”状,轻病株有康复的能力[73]。玉米鼠耳病症状类似于玉米粗缩病,但玉米粗缩病株矮缩不如鼠耳病明显,叶背瘿瘤沿叶脉纵向排列,叶肉无横向皱缩,叶缘无内卷,心叶不卷曲,轻病株极少恢复[71]。
6 其他病毒
伴随着分子生物学的飞速发展,对玉米病毒病病原的检测及鉴定也越来越精确,除上述病毒以外,一些病毒在国内虽然没有大面积暴发过,但也曾被检测到,如番茄丛矮病毒科(Tombusviridae)玉米褪绿斑驳病毒属(Machlomovirus)成员玉米褪绿斑驳病毒(MCMV)是正义单链RNA病毒,基因组结构见图5-A所示,p50蛋白及其通读产物p111蛋白,即RNA聚合酶。p32蛋白与症状的形成有关。外壳蛋白 (CP) 和2种运动相关蛋白 p7a 和 p7b 是细胞间运动所必需的。p7a编码p31蛋白是有效全身感染所必需的[76]。p31是MCMV的主要致病性决定因素,它可以劫持玉米过氧化氢酶 (ZmCATs) 以减弱水杨酸 (SA) 介导的防御反应[77]。但MCMV感染的分子机制尚不明确。MCMV是我国重要的对外检疫性病毒[78]。MCMV通过叶甲传播,玉米受侵染后产生的症状主要包括叶片褪绿、叶色斑驳坏死、植株矮化、果穗畸形等症状(图5-B)。MCMV能够与其他病毒复合侵染玉米,引发玉米致死性坏死病并造成严重危害[79-80]。
玉米线条病毒(Maize streak reunion virus,MSRV)属于双生病毒科(Geminiviridae)玉米线条病毒属(Mastrevirus),为单链环状DNA病毒(图5-C),于2014年在我国云南首次被检测到[81],导致植物叶片产生条斑等症状(图5-D)。除此之外,还有部分玉米上检测到我国未发现的病毒,随着科技的进步,越来越多未被发现的病毒会逐渐暴露于人们的视野下。
7 小结
玉米病毒病在全世界广泛发生,造成全球的玉米生产损失,严重危害经济效益和农产品安全。人们必须对其进行细致的研究并用理论来指导实践生产,全面防控玉米病毒病害。病毒病一旦发生便无法根治,故对于玉米病毒病,防大于治,通过了解不同病毒的介体昆虫及传播途径;结合害虫发生动态和实际生产活动情况,对可能发生的玉米病毒病害有所准备,从根源上预防病毒病的发生。
同时本文阐述了玉米病毒病相关病原物的研究进展,包括相关病毒的首次报道、分类地位、分布范围、病毒特性等,总结常见的玉米病毒病病原,展示相关病毒的基因组结构,可方便识别和研究玉米病害,在生产中如有玉米病毒病的发生,可通过植株症状表现,缩小检索范围,快速地识别相关病原,有利于后续进一步的鉴定操作和生产处理,尽可能降低经济损失,为研究建立高效、安全的病毒防控技术提供理论指导。除此之外,现在对农产品的关注点并不单单是产量,还包括农产品品质,对玉米病毒病的深入了解能够更好地保证生产需求。根据上述国内关于玉米病毒病的总结,希望为后续相关病毒的深入研究提供一定的参考。
参考文献:
[1]王琦琪,陈印军. 我国玉米种植的优势分析[J]. 中国农业科技导报,2018,20(3):1-9.
[2]崔爱民,张久刚,张 虎,等. 我国玉米生产现状及发展变革[J]. 中国农业科技导报,2020,22(7):10-19.
[3]马先红,李 峰,宋荣琦. 玉米的品质特性及综合利用研究进展[J]. 粮食与油脂,2019,32(1):1-3.
[4]王晓鸣,段灿星. 玉米病害和病原名称整理及其汉译名称规范化探讨[J]. 中国农业科学,2020,53(2):288-316.
[5]王晓鸣. 六种重要玉米病害病原名称的厘定[J]. 中国农业科学,2018,51(18):3497-3507.
[6]杨俊伟,王建军,李彦良,等. 玉米病毒病的发生及综合防治[J]. 农业灾害研究,2019,9(6):6-7,32.
[7]方守国,王朝辉,韩成贵,等. 水稻黑条矮缩病毒基因组片段6编码一种非结构蛋白[J]. 华北农学报,2007,22(6):5-8.
[8]Zhou T,Wu L J,Wang Y,et al. Transmission of rice black-streaked dwarf virus from frozen infected leaves to healthy rice plants by small brown planthopper (Laodelphax striatellus)[J]. Rice Science,2011,18(2):152-156.
[9]張恒木,雷娟利,陈剑平,等. 浙江和河北发生的一种水稻、小麦、玉米矮缩病是水稻黑条矮缩病毒引起的[J]. 中国病毒学,2001,16(3):246-251.
[10]杨 青,郝俊杰,王新涛. 玉米粗缩病抗病基因定位和抗病育种研究进展[J].分子植物育种,2020,18(12):4021-4028.
[11]Zhang H M,Chen J P,Lei J L,et al. Sequence analysis shows that a dwarfing disease on rice,wheat and maize in China is caused by rice black-streaked dwarf virus[J]. European Journal of Plant Pathology,2001,107(5):563-567.
[12]Bai F W,Yan J,Qu Z C,et al. Phylogenetic analysis reveals that a dwarfing disease on different cereal crops in China is due to rice black streaked dwarf virus (RBSDV)[J]. Virus Genes,2002,25(2):201-206.
[13]Fang S,Yu J,Feng J,et al. Identification of rice black-streaked dwarf fijivirus in maize with rough dwarf disease in China[J]. Archives of Virology,2001,146(1):167-170.
[14]Li Y Q,Jia M G,Jiang Z D,et al. Molecular variation and recombination in RNA segment 10 of rice black-streaked dwarf virus isolated from China during 2007—2010[J]. Archives of Virology,2012,157(7):1351-1356.
[15]郑巧兮,沈菊英,龚祖埙,等. 一种新的纯化玉米粗缩病毒的方法及病毒形态的研究[J]. 生物化学与生物物理学报,1984(6):571-576.
[16]Wang Z H,Fang S G,Xu J L,et al. Sequence analysis of the complete genome of rice black-streaked dwarf virus isolated from maize with rough dwarf disease[J]. Virus Genes,2003,27(2):163-168.
[17]Isogai M,Uyeda I,Lee B C.Detection and assignment of proteins encoded by rice black streaked dwarf fijivirus S7,S8,S9 and S10[J]. The Journal of General Virology,1998,79(6):1487-1494.
[18]周 彤,吴丽娟,王 英,等. 灰飞虱从冷冻病叶获得水稻黑条矮缩病毒方法的研究初报[J]. 中国水稻科学,2010,24(4):425-428.
[19]陈声祥,洪 健,吕永平,等. RBSDV在玉米叶脉细胞内的侵染状态与灰飞虱传毒活力的关系[J]. 中国病毒学,2004,19(2):153-157.
[20]王 康,郑静君,张曙光,等. 室内试验证实南方水稻黑条矮缩病毒不经水稻种子传播[J]. 广东农业科学,2010,37(7):95-96.
[21]周国辉,许东林,李华平. 广东发生水稻黑条矮缩病病原分子鉴定[C]//中国植物病理学会2004年学术年会论文集.2004:223-225.
[22]周国辉,张曙光,邹寿发,等. 水稻新病害南方水稻黑条矮缩病发生特点及危害趋势分析[J]. 植物保护,2010,36(2):144-146.
[23]Zhou G H,Wen J,Cai D J,et al. Southern rice black-streaked dwarf virus:a new proposed Fijivirus species in the family Reoviridae[J]. Chinese Science Bulletin,2008,53(23):3677-3685.
[24]周国辉,温锦君,蔡德江,等. 呼肠孤病毒科斐济病毒属一新种:南方水稻黑条矮缩病毒[J]. 科学通报,2008,53(20):2500-2508.
[25]杨本荣,马巧月. 玉米粗缩病的病毒寄主范围研究[J]. 植物病理学报,1983,13(3):1-8.
[26]Wang Q,Yang J,Zhou G H,et al. The complete genome sequence of two isolates of southern rice black-streaked dwarf virus,a new member of the genus Fijivirus[J]. Journal of Phytopathology,2010,158:733-737.
[27]Yin X,Xu F F,Zheng F Q,et al. Molecular characterization of segments S7 to S10 of a southern rice black-streaked dwarf virus isolate from maize in Northern China[J]. Virologica Sinica,2011,26(1):47-53.
[28]张 彤,周国辉. 南方水稻黑条矮缩病研究进展[J]. 植物保护学报,2017,44(6):896-904.
[29]曹 杨,潘 峰,周 倩,等. 南方水稻黑条矮缩病毒介体昆虫白背飞虱的传毒特性[J]. 应用昆虫学报,2011,48(5):1314-1320.
[30]Pu L L,Xie G H,Ji C Y,et al. Transmission characteristics of Southern rice black-streaked dwarf virus by rice planthoppers[J]. Crop Protection,2012,41:71-76.
[31]Li Y Z,Cao Y,Zhou Q,et al. The efficiency of Southern rice black-streaked dwarf virus transmission by the vector Sogatella furcifera to different host plant species[J]. Journal of Integrative Agriculture,2012,11(4):621-627.
[32]Williams L E,Gordon D T,Nault L.Proceedings:international maize virus disease colloquium and workshop[EB/OL]. (1976-08-16)[2022-04-22]. https://agris.fao.org/agris-search/search.do?recordID=XF2016010266.
[33]史春霖,徐紹华. 北京玉米和高粱上的玉米矮花叶病毒[J]. 植物病理学报,1979,9(1):35-40,67-68.
[34]景 岚,姚学慧. 玉米矮花叶病毒生物学特征研究概述[J]. 榆林学院学报,2004,14(3):65-68.
[35]陈 刚,周广和,陈剑波. 玉米矮花叶病毒提纯及特性的研究[J]. 中国病毒学,1992,7(1):93-98.
[36]沈小娟. 抗矮花叶病RNA干扰表达载体构建及玉米茎尖转化[D]. 雅安:四川农业大学,2011.
[37]周伦理. 玉米矮花叶病研究进展[J]. 基因组学与应用生物学,2010,29(2):396-401.
[38]王海光. 玉米矮花叶病流行原因分析及预测方法[D]. 北京:中国农业大学,2003.
[39]张 虎,景晓雅,孙柳清,等. 玉米矮花叶病毒分离物基因组克隆及多样性分析[J]. 中国农业科技导报,2019,21(9):36-43.
[40]郑艳茹,翟玉山,邓宇晴,等. 甘蔗花叶病毒(SCMV)种群结构分析[J]. 福建农林大学学报(自然科学版),2016,45(2):135-140.
[41]高文臣,魏宁生,郭 明,等. 玉米矮花叶病毒原的生物学鉴定[J]. 北京师范大学学报(自然科学版),2000,36(2):250-255.
[42]程 晔,陈剑平,陈 炯. 一个引起玉米矮花叶病的甘蔗花叶病毒基因组全序列测定及其结构分析[J]. 中國科学(C辑:生命科学),2001,31(6):497-504.
[43]党明青. 玉米褪绿斑驳病毒致病性分析及与甘蔗花叶病毒协同侵染机制研究[D]. 杭州:浙江大学,2017.
[44]Padhi A,Ramu K. Genomic evidence of intraspecific recombination in sugarcane mosaic virus[J]. Virus Genes,2011,42(2):282-285.
[45]吕香玲,张宝石. 玉米矮花叶病研究进展[J]. 作物杂志,2007(3):27-31.
[46]李 莉,王锡锋,郝宏京,等. 甘蔗花叶病毒在玉米种子中的分布及其与种子传毒的关系[J]. 植物病理学报,2004,34(1):37-42.
[47]王卫兵,洪 健,周雪平. SrMV和SCMV侵染玉米的细胞超微病变比较研究[J]. 浙江大学学报(农业与生命科学版),2004,30(2):215-220.
[48]缪 倩,任春梅,吴丽莉,等. 江苏玉米矮花叶病病原鉴定[J]. 江苏农业科学,2013,41(2):118-121.
[49]Fan Z,Chen H,Cai S,et al. Molecular characterization of a distinct potyvirus from whitegrass in China[J]. Archives of Virology,2003,148(6):1219-1224.
[50]姜 辛. 白草花叶病毒变异的初步研究[D]. 北京:中国农业大学,2005.
[51]石秀清,王富荣,石银鹿,等. 玉米种质资源抗矮花叶病鉴定[J]. 植物遗传资源学报,2003,4(4):338-340,364.
[52]崔小雯. 白草花叶病毒种群遗传结构分析[D]. 泰安:山东农业大学,2012.
[53]King A M Q,Adams M J,Carstens E B. Virus taxonomy[EB/OL]. (2011-11-10)[2022-04-22]. https://www.sciencedirect.com/book/9780123846846/virus-taxonomy.
[54]Tosic M. Physical and serological properties of maize dwarf mosaic and sugarcane mosaic viruses[J]. Phytopathology,1974,64(3):312.
[55]李秀坤,刘昌林,周 羽,等. 玉米病毒病的研究进展[J]. 作物杂志,2015(3):13-16.
[56]Jaroová J,Beoni E,Kundu J K. Barley yellow dwarf virus resistance in cereals:approaches,strategies and prospects[J]. Field Crops Research,2016,198:200-214.
[57]刘艳莉,王艳芳,赵彦宏,等. 大麦黄矮病毒的生物信息学分析[J]. 南方农业学报,2018,49(9):1760-1767.
[58]吴尔福,王鸣岐. 玉米红叶病鉴定诊断及其化学防治[J]. 自然杂志,1984,6(2):84-86,160.
[59]李 娜. 大麦黄矮病毒GAV株系和小麦黄花叶病毒单克隆抗体的制备及其应用[D]. 杭州:浙江大学,2015.
[60]乔世英,闫佳会,郭青云.中国大麦黄矮病毒及BYDV-GAV株系研究进展[J]. 广东农业科学,2020,47(10):103-111.
[61]张惠芳,朱福成,杨凤琪,等. 大麦黄矮病毒侵染玉米研究初报[J]. 甘肃农业科技,1993(6):32-33.
[62]毛晓红,于 毅,张秀霞,等. 春季西葫芦日光温室黄瓜花叶病毒病发生规律与防虫网的防控效果[J]. 江苏农业科学,2017,45(16):96-98.
[63]Haber S. Situation review of barley yellow dwarf in Canada[EB/OL]. (1987-07-31)[2022-04-22]. https://www.researchgate.net/publication/236232303_Situation_review_of_barley_yellow_dwarf_in_Canada.
[64]Wang R,Wang N,Ye T,et al. Natural infection of maize by cucumber mosaic virus in China[J]. Journal of Phytopathology,2013,161:880-883.
[65]Mochizuki T,Ohki S T. Cucumber mosaic virus:viral genes as virulence determinants[J]. Molecular Plant Pathology,2012,13(3):217-225.
[66]王 蓉. 用于玉米基因功能研究的黄瓜花叶病毒基因沉默载体的创建[D]. 北京:中国农业大学,2016.
[67]Feng J L,Liu S S,Wang M N,et al. Identification of microRNAs and their targets in tomato infected with Cucumber mosaic virus based on deep sequencing[J]. Planta,2014,240(6):1335-1352.
[68]Callaway A,Giesman-Cookmeyer D,Gillock E T,et al. The multifunctional capsid proteins of plant RNA viruses[J]. Annual Review of Phytopathology,2001,39:419-460.
[69]Wang R,Yang X X,Wang N,et al. An efficient virus-induced gene silencing vector for maize functional genomics research[J]. The Plant Journal,2016,86(1):102-115.
[70]李小珍,劉映红,李雪燕.玉米鼠耳病的发生与防治研究[J]. 玉米科学,2004(4):89-91.
[71]林代福,彭丽娟,李 明,等. 玉米鼠耳病症状识别与发病因素调查[J]. 山地农业生物学报,2000,19(4):262-265.
[72]李雪燕.玉米鼠耳病病原鉴定及检测系统建立[D]. 重庆:西南农业大学,2004.
[73]青 玲,牛颜冰,刘映红,等. 玉米鼠耳病研究进展[J]. 植物保护,2005,31(4):14-18.
[74]李祥坤,邓先明,梁映鸿,等. 玉米鼠耳病传毒叶蝉研究[J]. 西南农业大学学报,1996,18(6):599-601.
[75]李小珍,刘映红,青 玲,等. 玉米鼠耳病及其介体二点叶蝉研究进展[J]. 生态学报,2006,26(4):1270-1279.
[76]Kay S.Analysis of gene functions in maize chlorotic mottle virus[J]. Virus Research,2016,222:71-79.
[77]Jiao Z Y,Tian Y Y,Cao Y Y,et al. A novel pathogenicity determinant hijacks maize catalase 1 to enhance viral multiplication and infection[J]. The New Phytologist,2021,230(3):1126-1141.
[78]赵明富,黄 菁,吴毅歆,等. 玉米褪绿斑驳病毒及传播介体研究进展[J]. 中国农业科技导报,2014,16(5):78-82.
[79]李敬娜,王乃顺,宋 伟,等. 玉米褪绿斑驳病毒研究进展及防治策略[J]. 生物技术通报,2018,34(2):121-127.
[80]李晓静. 三七病毒病病原种类鉴定及其相关病毒分子变异分析[D]. 昆明:云南农业大学,2014.
[81]陈 莎. 深度测序鉴定玉米病毒及感病玉米组织中小RNA分析[D]. 杭州:浙江大学,2015.
[82]Lommel S,Kendall T,Siu N,et al. Characterization of maize chlorotic mottle virus[J]. Phytopathology,1991,81:819-823.
