棉蚜对吡虫啉和氟啶虫胺腈抗药性监测及抗性机理研究进展
2023-05-30李越,朱鹤,单莹,王子胜
李越,朱鹤,单莹,王子胜

摘要:棉蚜Aphis gossypii的发生会直接影响棉花的正常生长发育甚至威胁产量。吡虫啉和氟啶虫胺腈是防治棉蚜的常用杀虫剂。但因长期使用,我国各地棉蚜已对其产生不同程度的抗药性。就棉蚜在我国三大棉区对吡虫啉和氟啶虫胺腈的抗药性水平以及抗药性机理进行归纳,为后续防治棉蚜中合理用药和抗性治理提供理论依据。
关键词:棉蚜;吡虫啉;氟啶虫胺腈;抗药性监测;抗性机理
棉花是世界重要的经济作物之一。我国是世界棉花生产第二大国、棉花消费第一大国,因此,保障棉花的安全生产十分重要。20世纪50年代我国共划分有五大棉区即为西北内陆棉区、长江流域棉区、黄河流域棉区、辽河流域棉区和华南棉区[1]。随时间推移,经多年发展,西北内陆棉区、长江流域棉区和黄河流域棉区成为三大主产棉区,并且近年长江流域和黄河流域棉区棉花产量大幅缩减,重心逐渐向西北内陆棉区偏移(主要集中在新疆地区)[2-3]。棉花病虫害的发生直接关系到棉花的产量及品质。棉花上常见的蚜虫主要有棉蚜Aphis gossypii和棉长管蚜Acyrthosiphon gossypii[4-6]。棉蚜属半翅目同翅亚目蚜科, 为世界性刺吸式口器害虫,适应能力强,以越冬卵在黄金树等其他寄主上越冬[5],有两性生殖及孤雌生殖方式。棉蚜种群数量达到峰值的时间可随温度升高而缩短,有翅蚜可迁飞扩散至附近植株为害[6]。棉蚜取食棉花汁液,尤其在现蕾前后对棉花生长发育影响较大,在取食过程中分泌的蜜露会诱发产生霉菌,影响棉花产量以及品质[7-8]。
现阶段我国防治棉蚜主要还是依靠化学杀虫剂,常用的杀虫剂有吡虫啉、噻虫嗪、啶虫脒、氟啶虫胺腈、高效氯氟氰菊酯等。吡虫啉为新烟碱类杀虫剂,氟啶虫胺腈为砜亚胺类杀虫剂,二者皆为烟碱型乙酰胆碱受体(nicotinic acetylcholine receptor, nAChR)竞争性调节剂,但氟啶虫胺腈具有独特的结构活性[9-10]。自问世以来,吡虫啉的使用量持续增长。然而需要注意的是长期使用化学杀虫剂,容易导致害虫产生抗药性[11]。我國科技工作者已注意到棉蚜产生抗药性的问题,并积极展开了抗药性监测。拟总结国内3个主产棉区(以西北内陆棉区为主)棉蚜对吡虫啉及氟啶虫胺腈的抗药性情况、抗性机理研究进展以及现阶段针对棉蚜的综合防治技术,旨在为棉花生产中科学合理用药、综合防治棉蚜提供参考。
1 国内各棉区棉蚜对吡虫啉和氟啶虫胺腈抗药性监测进展
1.1 西北内陆棉区
西北内陆棉区主要分布在新疆、甘肃河西走廊一带[1]。新疆是我国棉花主产区,占我国总体棉花生产的绝大部分。国家统计局统计数据显示:2021年新疆棉花种植面积为250.61万hm2,占全国棉花播种面积的82.76%;皮棉产量为512.9万t,占全国棉花总产量的89.50%。要保障新疆地区棉花生产安全,一定要时刻关注棉区的气候、病虫害等可能会威胁产量的因素。
在新疆南疆棉田为害的蚜虫有棉蚜、棉长管蚜、桃蚜Myzus persicae、拐枣蚜Xerophilaphis plothikovi和棉黑蚜A. atrata[12],主要为棉蚜和棉长管蚜[6]。在北疆棉田为害的蚜虫种类主要为棉蚜、棉长管蚜和棉黑蚜[13]。近几年对新疆棉蚜抗药性的监测发现,新疆多数地区的棉蚜已对吡虫啉产生了极高水平的抗药性,大部分地区的棉蚜对氟啶虫胺腈的抗药性也处于低抗或中抗水平(表1)。2020年石河子、吐鲁番、沙湾、乌苏棉蚜对吡虫啉的抗性倍数(resistance ratio, RR)已在2.5万以上,库尔勒、伊犁、奎屯、阿拉尔、昌吉棉蚜对吡虫啉的抗性倍数也在1 000以上[14],这表明吡虫啉已经几乎无法在上述地区的防治棉蚜工作中起到作用。此外,奎屯、库尔勒以及石河子的棉蚜对吡虫啉的抗药性已由2017年的低抗水平(抗性倍数分别为8.85、8.39、5.03)飙升至2020年的高抗水平[14-15]。2021年通过监测发现,新疆库尔勒、阿拉尔、博乐、昌吉、库车、奎屯、沙湾、沙雅、石河子、图木舒克、乌苏等地的棉蚜对吡虫啉已达到高水平抗性,抗性倍数均在1 000以上[16]。而对氟啶虫胺腈,仅有哈密、五家渠、昌吉、喀什地区的棉蚜处于低抗水平,其他地区棉蚜的抗性均处于中等水平至高水平(表1)。奎屯、库尔勒、石河子棉蚜由2017年对氟啶虫胺腈敏感(抗性倍数分别为3.65、2.51、1.43)发展为2020―2021年的中等至高等抗性水平[14-16]。可见,短短几年时间,棉蚜对吡虫啉和氟啶虫胺腈的抗药性发展迅速。建议表1中田间种群对吡虫啉抗性倍数高于100的地区在防治棉蚜过程中避免使用该药剂,其他地区也要注意轮换使用其他作用机理不同的药剂。
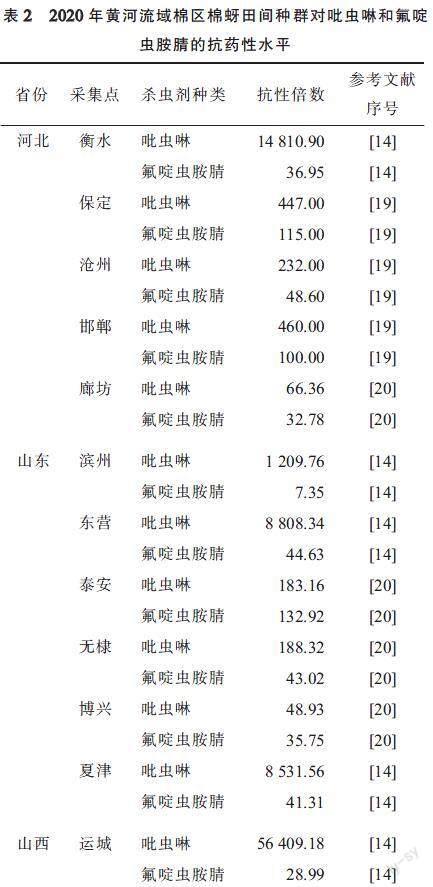
1.2 黄河流域棉区
黄河流域棉区早期主要包括河北和山东两省、山西汾河下游、陕西关中、河南除南阳外、安徽、江苏两省淮河以北地区[1],现在主要包括天津、河北、山西、山东、河南、陕西[18]。从表2可以看出,除山东滨州棉蚜对氟啶虫胺腈为低抗外,黄河流域棉区其他采集点的棉蚜对吡虫啉和氟啶虫胺腈的抗药性达到了中等至高抗水平。对吡虫啉的抗药性,除河北廊坊、山东博兴棉蚜在中等水平外,其余地区棉蚜均为高抗水平。其中,河北衡水,山东滨州、东营、夏津,山西运城棉蚜对吡虫啉的抗性倍数均在1 000以上,因此上述地区在防治棉蚜时应避免使用吡虫啉。2020年棉蚜对氟啶虫胺腈抗性的监测显示,仅河北保定、山东泰安地区棉蚜处于高抗水平,其余地区均处于低至中等水平抗性。但2021年河北衡水棉蚜种群对氟啶虫胺腈的抗药性也达到高抗水平(抗性倍数为140~151)[16]。此外,安静杰等[19]通过皮尔逊相关分析6种杀虫剂LC50对数值的相关性发现,河北保定棉蚜种群对氟啶虫胺腈与噻虫嗪、吡虫啉2种新烟碱类杀虫剂产生交互抗性,且河北沧州和邯郸的田间棉蚜种群对拟除虫菊酯类、氨基甲酸酯类和新烟碱类杀虫剂存在交互抗性。
1.3 长江流域棉区
长江流域棉区早期主要包括陕西汉中、河南南阳、四川、湖北、湖南、江西、浙江、安徽和江苏两省淮河以南、贵州和福建北部,现在主要包括江苏、安徽、江西、湖北、湖南[18]。据报道,2014年湖北襄阳、安徽萧县棉蚜对吡虫啉的抗药性处于中抗水平,抗性倍数分别为26.72、13.34,江苏东台棉蚜对吡虫啉则处于敏感水平(抗性倍数为1.48)[21]。但因近年来缺乏相关报道,该棉区棉蚜对吡虫啉和氟啶虫胺腈的抗药性水平不得而知。
2 棉蚜对吡虫啉以及氟啶虫胺腈的抗药性机理研究进展
目前关于棉蚜对吡虫啉和氟啶虫胺腈产生抗药性机理的研究报道主要涉及解毒代谢增强和靶点突变引起的靶标敏感度降低2个方面[21-24]。
2.1 棉蚜对吡虫啉和氟啶虫胺腈的解毒代谢研究进展
在棉蚜抗药性机理研究中常见的就是其解毒代谢能力增强,涉及的解毒酶主要为细胞色素P450(cytochromeP450, CYP450)、羧酸酯酶(carboxylesterase, CarE)、谷胱甘肽S-转移酶(glutathione S-transferase, GST)。棉蚜吡虫啉抗性品系与敏感品系相比,CarE、P450活性显著升高,GST活性也有所升高[25];抗性品系棉蚜体内的羧酸酯酶基因、细胞色素P450基因CYPCY3-1和CYPCY3-2也与棉蚜对吡虫啉产生抗药性有密切的关系[25]。在绿桃蚜(Myzus pericae)氟啶虫胺腈抗性品系中,发现尿苷二磷酸葡萄糖醛酸转移酶(uridine diphosphate glucuronic acid transferases, UGT)基因UGT344P2和细胞色素P450基因CYP380C40很可能与其产生抗药性相关[26]。而在氟啶虫胺腈抗性品系中棉蚜体内P450和CarE活性显著升高,而且细胞色素P450基因CYP6CY13和CYP6CY19的过量表达被证实与棉蚜对氟啶虫胺腈产生抗药性有关[27]。三磷酸腺苷结合盒转运体(ATP-binding cassette transporter, ABC transporter)蛋白超家族,在解毒过程中发挥着重要作用。已发现ABCA1和ABCD1基因与棉蚜对氟啶虫胺腈产生抗性相关[28]。此外,在氟啶虫胺腈抗性品系中还发现蜕皮激素受体(ecdysone receptor, EcR)、超气门蛋白(ultraspiracle proteins, USP)和保幼激素结合蛋白(juvenile hormone binding protein, JHBP)表达量升高,但卵黄原蛋白(vitellogenin, Vg)表达水平降低等现象,这与棉蚜产生抗药性后的适应性有关,但与抗药性的具体关系尚不明确[29]。
2.2 棉蚜对吡虫啉和氟啶虫胺腈的靶点突变研究进展
吡虫啉的作用靶点为烟碱型乙酰胆碱受体(nicotinic acetylcholine receptor, nAChR),目前已鉴定出棉蚜nAChR的β1亚基R81T、V62I、K264E点突变和R81T-V62I共同突变等会引起棉蚜对吡虫啉的抗药性[15,21,25,30]。通过分子对接发現,棉蚜nAChR的β1亚基R81T、V62I单突变和R81T-V62I共同突变会改变棉蚜nAChR与吡虫啉的结合构象,导致nAChR与吡虫啉的亲和力降低[14]。我国已在山西运城田间棉蚜品系中发现了V62I、R81T和K264E点突变,在湖北襄阳、安徽萧县和山东滨州发现了R81T点突变(杂合子),在河南郑州发现了R81T相应等位基因的纯合突变,在山东滨州和安徽萧县发现了K264E点突变[21]。除此之外,抗吡虫啉棉蚜品系的nAChR β1亚基基因表达量与敏感品系相比有所下降[21,25]。虽然棉蚜对氟啶虫胺腈已产生抗药性,但暂未发现氟啶虫胺腈与棉蚜结合的亲和力有明显变化[14]。
3 现阶段防治技术及展望
3.1 现阶段主要防治技术
现阶段防治棉蚜的主要方法依然是农业防治、物理防治、生物防治和化学防治等几类。农业防治主要措施有破坏棉蚜越冬环境,以减少越冬虫源;种植抗虫品种;加强田间管理促进棉花生长发育,提高棉株抵抗力;及时清洁棉田以及周边杂草、残枝等。物理防治方法主要为在田间设置黄板或黄皿诱蚜[13,31-32]。生物防治措施主要有保护利用天敌,捕捉瓢虫、草蛉等天敌在棉田释放[33],并在棉田周围种植有利于棉蚜天敌生存的植被。同时,要注意对其他蚜虫适当放宽防治,目的是增加棉田天敌种群数量,利于棉蚜生物防治[12]。
在棉蚜化学防治上,有以下3个抗性治理策略:首先,要对现有棉田进行棉蚜抗药性动态监测以了解本地棉蚜抗药性发展情况。因为,棉蚜种群一旦对吡虫啉产生抗药性,即便停止用药,短时间内(12代)也很难再恢复对吡虫啉的敏感性,且会对同种类(新烟碱类)以及有机磷酸酯类、菊酯类、氨基甲酸酯类、吡啶甲亚胺类等其他种类杀虫剂产生交互抗性[34-35]。其次,在化学防治中可以添加增效剂磷酸三苯酯、增效醚等来提升杀虫剂毒力,从而延缓棉蚜对杀虫剂抗药性的发展[22]。最后,轮用或者混用杀虫剂,避免长期单一使用吡虫啉或其他药剂。
3.2 展望
归纳整理了三大棉区棉蚜对吡虫啉以及氟啶虫胺腈的抗药性发展情况。总体上看,西北内陆棉区新疆和黄河流域棉区大多数地区的棉蚜对吡虫啉产生了极高水平的抗药性,在防治过程中应注意避免使用该杀虫剂。虽然对氟啶虫胺腈的抗药性多处于低到中等水平,但仍要注意与其他杀虫剂的混用或是轮换使用,防止其抗药性进一步发展。同时,各棉区应加强做好对本地棉蚜抗药性的监测工作,不限于吡虫啉和氟啶虫胺腈这2种杀虫剂。此外,应加强对棉蚜抗药性机理的研究,验证棉蚜对吡虫啉产生抗药性相关基因的功能,找到棉蚜对氟啶虫胺腈产生抗药性的关键途径等,以便于今后更好地防治棉蚜。
參考文献:
[1] 黄滋康,崔读昌. 中国棉花生态区划[J]. 棉花学报,2002,14(3):185-190.
[2] 王素云. 中国棉花集中生产带划分探讨[J]. 中国棉花,1994,21(4):5-6.
[3] 马春玥,买买提·沙吾提,姚杰,等. 1950-2015年中国棉花生产时空动态变化[J]. 地理学报,2020,75(8):1699-1710.
[4] 朱弘复. 棉虫发生趋势和预测预报[J]. 昆虫知识,1960(2):57-62.
[5] 张强,李贤超,余璐,等. 2020年新疆兵团棉花主要病虫害发生情况及原因分析[J]. 中国棉花,2020,47(12):38-40.
[6] 冯丽凯,高桂珍,吕昭智,等. 不同温度条件对棉蚜、棉长管蚜种间竞争关系的影响[J]. 应用昆虫学报,2015,52(3):557-565.
[7] 汪飞,李号宾,马祁,等. 新疆棉花现蕾前后棉蚜危害对棉花生长发育及产量影响的研究[J]. 新疆农业大学学报,1997,20(4):82-84.
[8] 王小丽,吴娜,张玉栋,等. 取食蚜害后的棉花对棉蚜营养代谢的影响[J]. 应用昆虫学报,2022,59(4):854-861.
[9] Insecticide Resistance Action Committee. IRAC mode of action classification scheme[EB/OL]. [2022-05-31]. https://irac-online.org/mode-of-action/.
[10] Sparks T,Watson G,Loso M,et al. Sulfoxaflor and the sulfoximine insecticides: chemistry, mode of action and basis for efficacy on resistant insects[J]. Pesticide Biochemistry and Physiology,2013,107(1):1-7.
[11] 安静杰,党志红,高占林,等. 河北省棉蚜对新烟碱类杀虫剂敏感基线及抗药性水平[J]. 河北农业大学学报,2018,41(4):112-116.
[12] 李号宾,吴孔明,徐遥,等. 南疆地区棉田蚜虫种群数量动态研究[J]. 新疆农业科学,2008,45(4):670-675.
[13] 芦屹,李晶,魏新政,等. 新疆棉花蚜虫综合防治技术规程[J]. 中国棉花,2022,49(1):38-41.
[14] 李仁,梁平卓,程沈航,等. 我国棉蚜对吡虫啉和氟啶虫胺腈抗性水平监测与交互抗性分析[J]. 植物保护学报,2021,48(5):1104-1113.
[15] 赵鹏程,李焱,闫文静,等. 新疆棉蚜不同地理种群对杀虫剂的敏感性[J]. 石河子大学学报(自然科学版),2018,36(2):159-163.
[16] 任宗杰,秦萌,郭永旺,等. 2021年全国农业有害生物抗药性监测报告与治理对策(小麦、棉花、蔬菜部分)[J]. 中国植保导刊,2022,42(4):68-73.
[17] 邵振润,高聪芬,张帅,等. 水稻二化螟抗药性监测技术规程:NY/T 2058-2014 [M]. 北京:中国农业出版社,2016.
[18] 徐爱武,尚斌,李翼然. 我国三大棉区近二十年棉花单产水平比较与建议[J]. 棉花科学,2022,44(5):11-14.
[19] 安静杰,高占林,党志红,等. 河北省不同地区棉蚜种群对6种杀虫剂的抗性动态[J]. 农药学学报,2021,23(6):1123-1131.
[20] 崔丽,吕宝顺,芮昌辉,等. 华北地区棉蚜对5种杀虫剂的敏感性监测[J]. 农药科学与管理,2022,43(1):29-35.
[21] Chen X W,Li F,Chen A Q,et al. Both point mutations and low expression levels of the nicotinic acetylcholine receptor β1 subunit are associated with imidacloprid resistance in an Aphis gossypii (Glover) population from a Bt cotton field in China[J/OL]. Pesticide Biochemistry and Physiology,2017,141:1-8[2022-11-27]. https://doi.org/10.1016/j.pestbp.2016.11.004.
[22] 马康生,王静慧,解晓平,等. 棉蚜对新烟碱类杀虫剂的抗性现状及其治理策略[J]. 植物保护学报,2021,48(5):947-957.
[23] Wang Z J,Liang C R,Shang Z Y,et al. Insecticide resistance and resistance mechanisms in the melon aphid, Aphis gossypii, in Shandong, China[J/OL]. Pesticide Biochemistry and Physiology,2021,172:104768[2022-11-27]. https://doi.org/10.1016/j.pestbp.2020.104768.
[24] 梁彦,张帅,邵振润,等. 棉蚜抗药性及其化学防治[J]. 植物保护,2013,39(5):70-80.
[25] 郭天凤. 新疆棉区棉蚜对新烟碱类杀虫药剂抗性监测、风险评估及抗性机制的研究[D]. 北京:中国农业大学,2014.
[26] Pym A,Umina P,Reidy-Crofts J,et al. Overexpression of UDP-glucuronosyltransferase and cytochrome P450 enzymes confers resistance to sulfoxaflor in field populations of the aphid, Myzus persicae[J/OL]. Insect Biochemistry and Molecular Biology,2022,143:103743[2022-11-27]. https://doi.org/10.1016/j.ibmb.2022.103743.
[27] Ma K S,Tang Q L,Zhang B Z,et al. Overexpression of multiple cytochrome P450 genes associated with sulfoxaflor resistance in Aphis gossypii Glover[J]. Pesticide Biochemistry and Physiology,2019,157:204-210.
[28] Wang L,Zhu J S,Cui L,et al. Overexpression of ATP-
binding cassette transporters associated with sulfoxaflor resistance in Aphis gossypii Glover[J/OL]. Pest Management Science,2021,7(9):4064-4072[2022-11-27]. https://doi.org/10.1002/ps.6431.
[29] Ma K S,Tang Q L,Xia J,et al. Fitness costs of sulfoxaflor resistance in the cotton aphid, Aphis gossypii Glover[J]. Pesticide Biochemistry and Physiology,2019,158:40-46.
[30] Wang L,Wang Q Q,Wang Q Y,et al. The feeding beha-vior and life history changes in imidacloprid-resistant Aphis gossypii Glover(Homoptera: Aphididae)[J]. Pest Management Science,2020,76(4):1402-1412.
[31] 迟宝杰,董合忠. 间(套)作对棉田病虫害的防控效应及其风险控制[J]. 棉花学报,2019,31(4): 341-351.
[32] 李小飞,韩迎春,王国平,等. 棉田间套复合体系提升生态系统服务功能研究进展[J]. 棉花学报,2020,32(5):472-482.
[33] 高立伟. 棉花蚜虫综合防治技术[J]. 现代农村科技,2011(5):18.
[34] 陈小坤. 抗吡虫啉棉蚜的交互抗藥性及机理的研究[D]. 泰安:山东农业大学,2014.
[35] 杨媛雪,李卓,段爱玲,等. 棉花花生间作田中花生蚜对吡虫啉代谢抗性机制初步研究[J]. 棉花学报,2020,32(2):151-157.
(责任编辑:杨子山 责任校对:王小璐)
