长期结实和不结实红松针叶光合生理参数的差异
2023-05-29陆天宇沈海龙王元兴
林 强,陆天宇,2,沈海龙*,王元兴,张 鹏
(1.林木遗传育种国家重点实验室,东北林业大学林学院,黑龙江 哈尔滨 150040;2.辽宁省农业科学院,辽宁 沈阳 110161;3. 吉林省林业科学研究院,吉林 长春 130031)
红松(Pinuskoraiensis)是东北温带湿润地区地带性顶极群落——阔叶红松林的建群种,是优质用材树种,也是世界四大食用松树种之一[1-2]。优质用材林是保障国家木材供给安全、尤其是大宗珍贵用材供给安全的重要保障;生产食用松籽则是充分发挥红松生产潜能、“以短养长”促进林区经济发展和保障林农长期经济收益的有效途径。因此,培育同时满足优质大径材生产和食用松籽需求的红松林是在适宜立地条件下优选的培育策略[2-5]。红松种内变异丰富,利用生长、结实性状等可以筛选出生长与结实性状遗传增益均较高的品系[4,6-7]。解析生长与结实性状上产生差异的个体、家系和无性系等的生物学机理,对果材兼用林品种选育和培育均具有重要意义。
已有研究表明,不同植物之间光合生理特性存在差别,针对光合参数的分析已经成为研究遗传资源、新品种培育和作物栽培技术改进的基础[8-13],如木棉(Bombaxceiba)株高与种源的净光合速率(Pn)呈显著正相关,Pn可作为选取种源的参考指标[8];红松光合作用相关指标在不同无性系之间差异显著,且与环境因子间显著相关,可作为无性系选择和生长发育的环境调控依据[10];不同种源山桐子(Idesiapolycarpa)在不同月份的光合特性差异可为种源选择和栽培管理方式制定提供参考[11];辣椒(Capsicumannuum)氮素的供应影响花和果蔗糖供给进而影响具有不同源库关系品种的产量性状[12]。利用模式植物研究光合作用机理非常重要,而利用作物研究光合作用机理对其在实践中的应用更为重要[13]。
本研究以吉林省露水河林业局红松种子园中生长环境、营养生长状态相似,但结实量差异巨大的30年生红松个体为对象,研究生长季内其针叶光合生理参数差异,分析其结实状态与针叶光合能力的关系,探索红松果材兼顾培育相关的光合作用机理,以期为果材兼用良种选育、培育措施的制定提供理论参考,也为深入研究红松营养生长和生殖生长关系及调控生物学技术奠定基础。
1 材料与方法
1.1 研究区概况
试验地点设置于吉林省露水河林业局(127°29′~128°02′E,42°20′~43°29′N)红松种子园实生区,红松栽植密度为3 m×3 m。样地海拔850 m,东北坡,平均坡度为8°,土壤为暗棕壤。属温带大陆性季风气候,年平均气温2.7 ℃,7月平均气温-19.0 ℃,绝对高温为34.2 ℃,1月平均气温-19.5 ℃,绝对低温-44.4 ℃,≥10 ℃有效积温2 590 ℃;年平均日照2 117 h;年平均降水量871 mm,最大降水量1 170 mm,多集中在6—8月;初霜期多为9月中旬,晚霜一般出现在5月下旬,全年无霜期107~122 d。
1.2 观测样木选择
根据在露水河红松种子园实生区内红松母树结实情况多年观察结果,并结合2017年的采种数据,在林缘(个体生长空间不受密度控制,营养生长和生殖生长均处于自由树状态)和林内(个体生长空间受密度控制,营养生长和生殖生长均处于林内树状态)各选择3对结实状况差异明显的红松母树(表1)。每对母树中1株结实量大且几乎每年结实,另1株几乎每年都不结实,但二者树干和枝条等营养器官的生长状态相似,所处立地条件一致。针对母树搭建分层观测架,对各个冠层进行光合和生长指标测定。
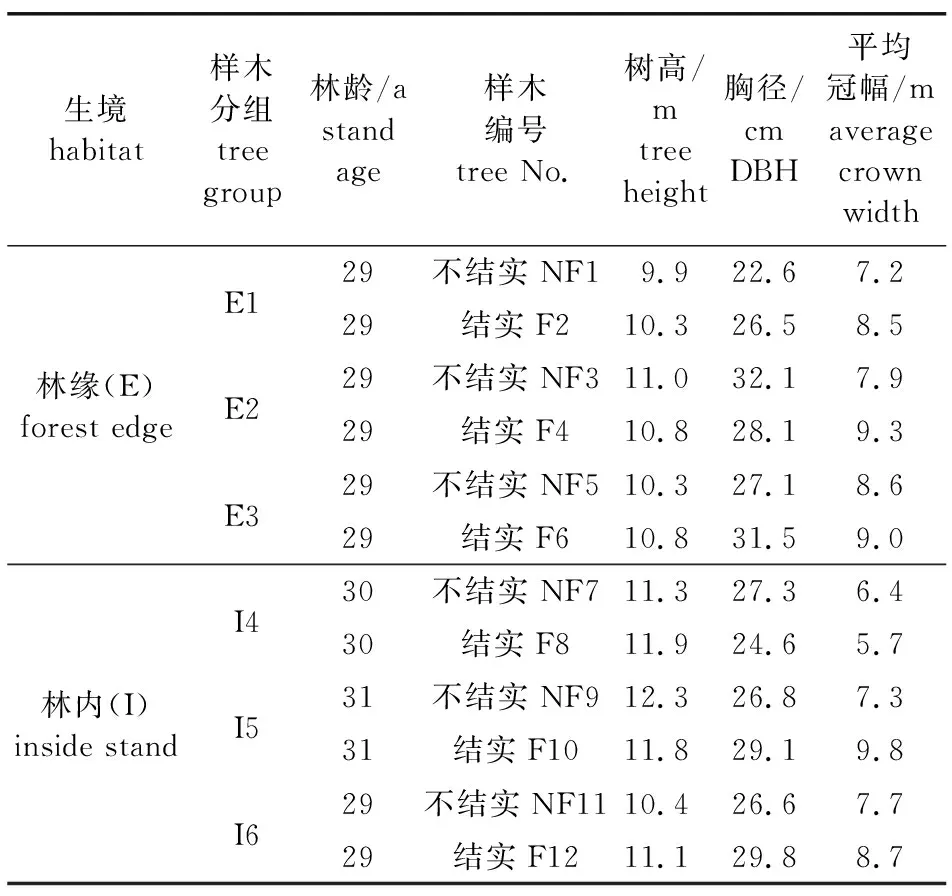
表1 露水河红松种子园样木概况
1.3 指标测定
1.3.1 光合特性观测
使用便携式CO2/H2O红外气体分析仪(Li-6400,Li-Cor. Lincoln,NE,USA)于生长季(5—9月)每月中旬测定光合特性,选择在晴朗无风的天气于9:00—11:00进行测定。测定时选取各冠层南向中部枝条。在测量过程中,叶室温度与外界一致,利用缓冲瓶保持叶室CO2含量与大气一致,为保证叶室内测定针叶的一致性,对其进行叶面积的测定,并按照比例进行换算。测量前对针叶进行20~30 min的诱导,有效光合辐射(PAR)为1 000 μmol/(m2·s),随后以红蓝光源(Li-6400-02B,Li-Cor. Lincoln,NE,USA)设置光量子通量密度梯度为2 000、1 500、1 000、800、600、400、200、100、50、20、0 μmol/(m2·s)。使用仪器的光响应曲线测定光合强度,仪器自动完成光响应曲线绘制。根据文献[14-15]计算出表观量子效率(AQY)。
1.3.2 比叶面积及叶绿素含量测定
比叶面积(SLA)即叶片光合面积和烘干质量的比值。在上中下3个冠层南向选取1年生健康针叶,使用Epson V700扫描仪(Seiko Epson Corporation,JPN)扫描叶面积后,将针叶装于信封中65 ℃烘干至质量恒定。比叶面积为叶面积(cm2)与叶片干质量(g)之比。
取部分针叶擦净表面的称取0.2 g剪碎后样品,置于50 mL离心管中并用40 mL体积分数99.5%的丙酮、95%的乙醇体积比1∶1的混合液于10~25 ℃避光条件下浸提至材料脱色变白。参照文献[3]的方法测定和计算光合色素的含量。
1.3.3 非结构性碳水化合物(NSC)含量测定
取中冠层南向1年生正常生长轮枝枝条上1年生针叶作为针叶样品。将样品放置于微波炉中中火加热5 min进行杀青处理,随后放置于65 ℃烘箱中烘干48 h至质量恒定。利用球磨仪对样品进行粉碎处理,过孔径0.15 mm筛,用于NSC的测定。使用蒽酮比色法[16]对可溶性糖和淀粉的含量进行测定,NSC含量为可溶性糖和淀粉含量之和。
1.4 数据处理
使用SPSS 25.0对不同结实特性红松的针叶叶绿素含量、光响应参数、NSC含量进行双因素方差分析(MANOVA),检验结实、季节(月份)和生境及其交互作用对指标影响是否显著,若显著则进行邓肯检验(Duncan),交互作用显著时进行简单效应分析。图表数据均为红松各指标平均值。
2 结果与分析
2.1 不同结实特性红松针叶比叶面积和叶绿素含量
对不同结实特性红松在不同生境下、不同月份间的叶绿素和比叶面积(SLA)进行双因素方差分析(表2),结果表明两类结实特性红松叶绿素a、b、类胡萝卜素和总叶绿素含量差异极显著(P<0.001),SLA和叶绿素a/b(Chl a/b)差异不显著;不同结实特性红松针叶在不同月份间光合色素及SLA差异极显著(P<0.001),不同生境间针叶类胡萝卜素含量差异显著(P<0.05),结实和月份的交互作用、结实和生境的交互作用对叶绿素和SLA的影响不显著(表2)。

表2 结实、月份、生境及交互作用对红松针叶叶绿素含量、SLA、NSC含量的影响
在相同月份,林缘和林内的结实和不结实红松间叶绿素和比叶面积间存在差异(表3)。6月林缘红松叶绿素a/b在结实和不结实之间存在显著差异(P<0.05);8月林缘红松叶绿素a、类胡萝卜素和总叶绿素含量在结实和不结实之间存在显著差异(P<0.05),林内红松比叶面积在结实和不结实之间存在显著差异(P<0.05);9月林缘红松除叶绿素a/b与比叶面积外,结实和不结实红松其余指标均存在显著差异(P<0.05),林内红松类胡萝卜素含量和叶绿素a/b在结实和不结实之间存在显著差异(P<0.05)。无论是林缘还是林内,叶绿素a、叶绿素b、类胡萝卜素和总叶绿素含量在各月份均表现为不结实大于结实。叶绿素a/b在6—8月时,林缘红松结实大于不结实,林内则相反。5月林内结实红松叶绿素a/b更高;林缘则是不结实更高,9月林缘和林内叶绿素a/b均表现为结实大于不结实;比叶面积除7月林内,8、9月林缘外均表现为结实大于不结实(表3)。

表3 相同月份不同生境结实和不结实红松叶绿素和比叶面积差异
两种结实特性红松叶绿素含量和比叶面积随月份变化趋势基本一致(图1),Chl a、Chl b、类胡萝卜素(Car)和总叶绿素(Chl T)含量在生长季期间表现为下降趋势,而SLA在5—9月则表现为上升,Chl a/b在5—8月下降,8—9月上升。在整个生长季期间,红松的Chl a、Chl b、Car和Chl T含量均表现为不结实大于结实,而Chl a/b则相反,SLA除7月外均表现为结实大于不结实。在9月时,结实和不结实红松针叶Chl a、Chl b、Chl T和Car含量达到显著差异,其他月份结实和不结实红松差异不显著(图1)。

不同小写字母表示相同月份下结实和不结实红松之间差异显著。Different lowercase letters indicate the difference between fruiting and non fruiting P. koraiensis in the same month.
2.2 不同结实特性红松针叶光响应参数差异
对不同结实特性红松不同冠层在生长季间的光响应参数进行双因素方差分析(表4)表明,不同结实特性红松针叶除光饱和点(LSP)差异达到显著水平外(P<0.05),其余指标均无显著差异;不同月份间也仅LSP达到极显著差异(P<0.01);冠层间光补偿点(LCP)和LSP则表现为极显著差异(P<0.001);结实与月份以及结实与冠层间交互作用影响差异不显著。
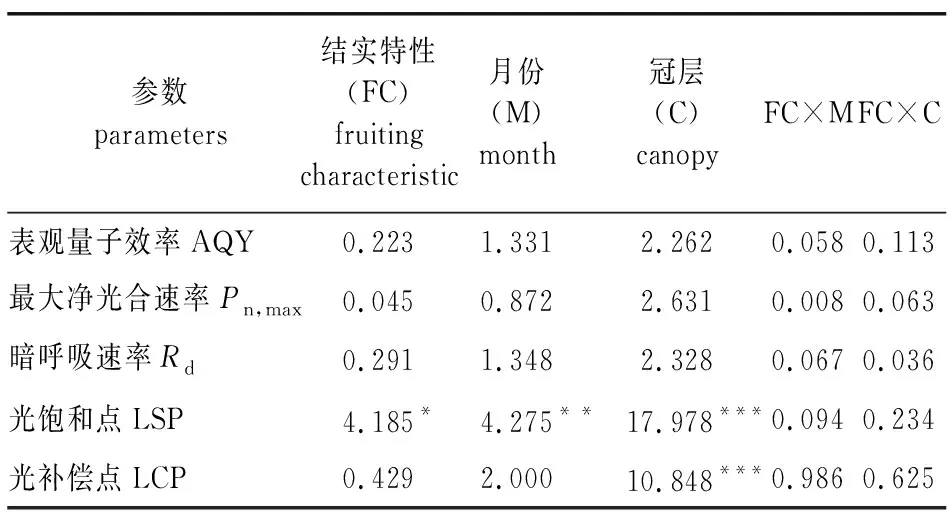
表4 结实、月份、冠层及交互作用对红松针叶光合响应参数的影响
对不同结实特性红松在不同月份间的光响应参数指标进行双因素分析(表5)发现,不结实红松的LSP和LCP均随冠层升高而增大。从整体上来看,表观量子效率(AQY)在整个生长季表现为上升趋势,最大净光合速率(Pn,max)和LSP则表现为下降趋势,暗呼吸速率(Rd)的趋势为先上升后不变,不结实红松LCP表现为下降趋势,但结实红松LCP则为先上升后下降最终保持平稳的趋势。5—9月间不结实红松AQY一直大于结实红松,而Pn,max和LSP则相反,表现为结实大于不结实,Rd除5月外也表现为结实大于不结实,LCP除5月和7月外均表现为结实大于不结实。除6月林缘下冠层AQY、林内中冠层LSP、7月林缘上冠层Pn,max在结实和不结实之间存在显著差异外(P<0.05),相同生境条件下相同冠层红松针叶光响应参数在结实和不结实之间没有显著差异(表5)。

表5 不同结实特性红松光合指标间差异
在整个生长季,红松上、中、下3个冠层间的LSP达到极显著差异(P<0.01),LCP在上冠层与中下两个冠层间存在极显著差异(P<0.01)(图2)。

相同颜色柱状图间不同小写字母表示不同冠层间的差异极显著(P<0.01)。Different lowercase letters in the same color of column indicate the difference between different canopy (P <0.01).
2.3 不同结实特性红松针叶非结构性碳水化合物差异性
对不同结实特性的红松可溶性糖、淀粉和非结构性碳水化合物(NSC)含量进行双因素方差分析,结果表明3个指标月份之间差异显著,不同生境下可溶性糖和淀粉含量差异极显著(P<0.001),但不同结实特性红松的月份和生境条件下差异均不显著(表2)。在整个生长季,不同结实特性红松针叶的可溶性糖、淀粉和NSC含量均表现为下降趋势,其中6—7月淀粉和NSC含量下降幅度较大(图3)。
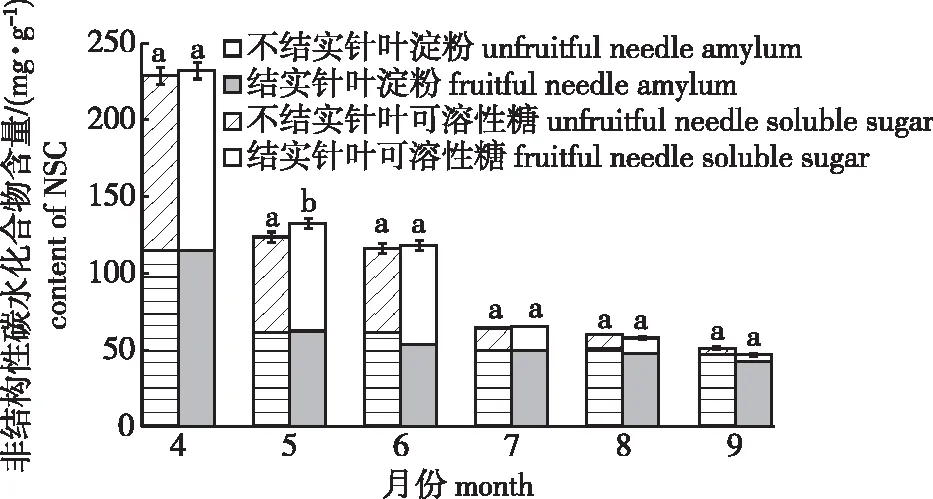
不同小写字母表示相同月份下各指标结实和不结实之间的差异显著。Different lowercase letters indicate the difference between fruiting and non fruiting in the same month.
除5月外,其他月份红松针叶可溶性糖含量均表现为不结实大于结实,除9月外其他月份淀粉含量均表现为结实大于不结实。4、7月NSC含量表现为结实大于不结实,而8、9月则相反。红松针叶可溶性糖含量在5月时,结实和不结实红松之间存在显著差异(P<0.05),其他情况(其他月份的可溶性糖、淀粉、NSC含量在结实和不结实红松之间)均不显著(图3)。
3 讨 论
光合作用不仅是物质生产更是产量形成的重要因素,光合产物为林木生长与果实发育提供物质基础,研究不同结实特性红松光合生理特点,可以为促进光合作用、提高果实产量服务,同时为果材兼用的生物学机理解析提供基础。
3.1 长期结实和不结实红松个体光合色素差异特点
光合色素是光合作用过程的主要指标,它能衡量植物光合作用强弱。叶绿素a将光能转化为化学能,叶绿素b负责捕获和传递光能,类胡萝卜素具有光能捕获和光破坏防御两大功能,而Chl a/b则代表叶片在强光条件下获取碳的能力[17]。两类结实特性红松Chl a、Chl b、Chl T和Car含量之间差异显著,表现为不结实红松大于结实红松。这说明不结实红松能够更高效的捕获光能并将其转化为化学能作为物质存储的基础[18],而结实红松则具有更高的Chl a/b,表明结实红松在光照条件下能够同化更多的碳[19]。
在整个生长季,不结实红松的叶绿素含量要高于结实红松且差异显著,这表明不结实红松进行了更强的光合,因此需要更多的叶绿素;但叶绿素a/b却刚好相反,这说明结实红松可以更好地利用光照,更高效地转化能量。不同结实特性红松的叶绿素含量在生长季期间均表现为降低,这有可能是光合作用的增加,加剧了叶绿素的消耗[20]。结实和不结实红松在相同月份或相同生境下的光合色素含量差异不显著,这意味着生长季节和环境不会影响光合作用这个过程,限制红松果实发育的因素可能与水分和温度等有关[17]。
3.2 长期结实和不结实红松个体比叶面积差异特点
SLA是表征植物对资源吸收能力的指标,能够体现针叶形态差异,且和单位生物量上的光合速率呈正相关。SLA还与叶面积指数有关,后者可以量化森林生态系统功能[21-22]。在整个生长季,结实红松SLA除7月外均大于不结实红松,这有可能是因为红松在结实过程中需要截获更多的光,增强了在强光条件下的自我保护能力[23],不同结实特性红松SLA在生长季期间均表现为上升趋势,两种特性的红松SLA在相同时间段、相同生境下的差异不显著,这表明SLA不是限制红松生殖生长的首要因素。
3.3 长期结实和不结实红松光响应参数及光合效率参数的差异特点
光响应参数作为光合指标的一部分,能够反应植物光合作用的强弱,对结实也有一定的影响,LSP反映了植物对强光照的利用能力,LSP值越高,说明植物利用强光照的能力越强,而AQY和LCP反映了植物在弱光条件下的光合性能,其值越高,说明植物对弱光的利用能力越强[24]。CO2饱和点和CO2补偿点是光合作用中植物对CO2利用能力的表现,CO2饱和点是反映植物利用CO2能力的重要指标,CO2补偿点越低表明植物利用低含量CO2进行光合作用积累干物质能力越强[25]。
在本研究中,生长季内AQY和LCP表现为不结实红松大于结实红松,而LSP、Pn,max和Rd则相反,其中只有LSP差异显著,其余指标均不显著,这表明不结实红松对弱光的利用能力要高于结实红松,而结实红松可以更高效地利用强光,且这种能力的差异会影响红松的正常结实。Pn,max揭示植物利用光能的最大能力,即正常生长的光合潜力[26]。Rd代表植物在无光条件下的呼吸速率,能够反映出植物光合积累物质的消耗能力,这说明结实红松具有更高的光合潜力和物质消耗能力,在其他研究中亦存在着佐证[27]。在相同月份或相同生境条件下不同结实特性的红松之间光响应参数差异不显著,这同样表明了光合效率的高低不会影响红松结实的过程。
两种结实特性红松的AQY、LSP和Pn,max变化趋势基本一致,这表明红松的光合效率参数季节变化不受结实过程的影响,而LCP在5月和7月时表现为不结实红松大于结实红松,这意味着LCP和果实的生长发育所关联[25]。通过研究发现,除LSP外,两种结实特性红松的其余指标在不同月份间差异不显著,这意味着利用强光的效率会影响红松的繁殖发育过程而其他条件对红松的结实过程不产生影响,同时LCP和LSP在冠层间存在显著差异,但不同结实特性红松指标在冠层间差异不显著,LCP和LSP表现为随冠层增高而逐渐增大,如日本落叶松与红松类似,LCP和LSP也随冠层的升高而增加,且冠层结构对光响应参数均有显著影响[28],这是因为光照强度影响叶片形态功能等多种指标,所以叶片的光合指标在冠层间存在垂直变化[25,29-30]。
3.4 长期结实和不结实红松非结构性碳水化合物的差异特点
NSC及其组分是植物光合作用的主要产物,也是植物合成各种物质的基础。NSC主要包括可溶性糖和淀粉,二者含量占90%以上且可以互相转化,这些指标在植物的生长发育过程中必不可少,且随外界环境的改变而改变[31],植物NSC含量与作为碳源的净光合作用和作为碳汇的呼吸作用有者不可分割的关系,能够维系源汇之间的平衡[32]。
两类结实特性红松针叶NSC含量在不同月份间存在着显著差异,这说明红松的繁殖发育和碳的存储和转变有着密不可分的关系[33]。其中可溶性糖表现为不结实红松大于结实红松,淀粉含量则相反,这说明不结实红松在生长发育期间具有更强的光合能力[27],而结实红松则形成了为应对生长季后期的低温渗透调节存储了较多的淀粉,在冬季需要时转化为可溶性糖的生长机制。相同生境下或相同月份间,两类结实特性红松NSC含量差异不显著,这与之前的研究有所不同,这有可能是测定指标时枝条选择差异所造成的[33]。
NSC含量在5—7月表现为结实红松>不结实红松,而8—9月则不结实红松>结实红松,但不同结实特性红松的NSC含量差异均不显著,这有可能是因为在结实期间,结实红松的更多的可溶性糖转化为淀粉进行物质的存储,NSC含量的变化趋势也证明了这点。在本研究中两种结实特性红松NSC含量在生长季均表现为下降趋势,这是因为红松在生长季需要消耗更多的碳储备,保证其他代谢活动的正常进行[34],在整个生长季,可溶性糖含量在不结实红松中更高,但淀粉和NSC含量则恰好相反,针叶初期生长依靠着生枝条存储的碳水化合物,而新生枝条则消耗已着生枝条上的1年生针叶[35],因此在生长季的结实红松需要存储更多的淀粉和NSC。
3.5 长期结实和不结实红松光合能力与非结构性碳水化合物的关系
碳的同化和代谢是植物生长发育的能量来源,碳库中的碳利用与碳源中的碳同化及代谢之间有紧密的联系,即叶片NSC含量变化可以指示叶片光合能力变化,但是有关这方面的关系的知识主要来源于草本植物,以树木为材料的研究成果尚不多见[36-37]。树木中关于生殖生长与光合能力关系的研究主要集中于阔叶果树或经济林木[36,38-39],对针叶树的研究鲜见相关报道。低负载量(果少)苹果(Maluspumila)树叶的碳同化水平,即整个生长季虽然高、低负载量树体叶碳同化水平变化趋势相似,但低负载量个体的同化水平始终低于高负载量个体的同化水平低,而果实收获后这种差异逐渐消失;低负载量个体积累高水平的NSC,即1天中低负载量个体NSC水平降低幅度始终小于高负载量个体的NSC水平[36]。油橄榄(Oleaeuropaea)库源比下降后作为源的叶净光合速率显著下降,叶片中可溶性糖和淀粉含量显著升高,与桃(Prunuspersica)和樱桃(Pinusspp.)的研究结果类似;油橄榄源叶净光合速率在库源比下降处理90 d后仍处于降低状态,但可溶性糖和淀粉含量的差异在处理30 d后就消失[38]。大久保桃(P.persica)去果处理后源叶的净光合速率显著降低,但对源叶的可溶性糖含量和淀粉酶活性无影响[39]。牡丹(Paeonia×suffruticosa)在花衰败前有花枝净光合速率明显低于无花枝,但可溶性糖含量却高于无花枝,有花枝可溶性糖昼夜无变化而无花枝翌日早较低;花衰败后,有花枝和无花枝净光合速率和可溶性糖含量的差异逐渐消失,翌日早晨有花枝和无花枝可溶性糖含量均变低[40]。红松生殖枝针叶净同化速率低于营养枝,NSC含量和比叶重高于营养枝,即生殖枝针叶净同化速率受NSC积累的限制[5]。这些研究表明,树木生殖生长的库碳和光合能力之间有着比较复杂的关系,生殖生长对NSC的利用可能对光合速率产生反馈[5,36,38-40],但也存在NSC的其他来源[5,36,38]。本研究选择了营养生长相似而生殖生长差异很大的2组红松个体,探讨光合参数和NSC的变化格局,研究结果展示了2组红松个体光合能力和NSC的一些差异格局;但是由于没有直接测定光合速率,也未与当时的结实状态建立直接联系,因此,还不能如这些研究那样探讨其光合能力与NSC变化的直接关系,下一步应该开展针对性的研究。
总体上看,吉林红松林区的结实红松具有更高的叶绿素a/b,这表明更高的光合潜力和物质消耗能力可以促进其结实。与不结实红松相比,结实红松在光照条件下会吸收更多的碳,并提高了能量转换效率,表明红松在结实过程的碳汇储备需要较好的光照条件支持,对非结构性碳水化合物的分析发现,结实红松具有更高含量的淀粉和NSC,这可能是为了在生长季能够存储更多养分为果实发育提供营养支撑。研究结果对解析红松果材兼用生物学机理提供了一定的科学依据,可为制定果材兼用培育措施提供理论支撑。
