褪黑素调控辣椒幼苗响应盐胁迫下的叶片生理特性
2023-05-27陈楠张维张晓明
陈楠 张维 张晓明
摘 要:為探究褪黑素对盐胁迫下辣椒叶片生长的调控效应,以天线1712和航椒4号2个辣椒品种幼苗为试验材料,设置不同浓度(10、50、100 μmol·L-1)褪黑素喷施处理,对盐胁迫下(200 mmol·L-1NaCl)辣椒苗叶片光合参数,叶绿素含量,PSⅡ光能转换效率,钠、钾离子含量等进行了测定,结合主成分分析研究了褪黑素处理对盐胁迫下辣椒幼苗叶片生理特性的影响。结果表明,3个浓度褪黑素处理显著缓解了盐胁迫对辣椒幼苗的不利影响,提高了盐胁迫下辣椒幼苗的净光合速率、气孔导度、胞间二氧化碳浓度、蒸腾速率、叶绿素含量和PSⅡ光能转换效率;降低了盐胁迫下幼苗的钠离子含量、丙二醛含量、电解质渗透率、过氧化氢含量,提高了钾离子含量和脯氨酸含量,50 μmol·L-1褪黑素处理缓解效果最为显著。由此可以得出,外源褪黑素可以减缓盐胁迫对辣椒幼苗产生的伤害,提高辣椒幼苗的抗盐性,50 μmol·L-1褪黑素处理为调控盐胁迫下辣椒幼苗叶片生长的最适浓度。
关键词:辣椒幼苗;褪黑素;盐胁迫;光合特性
中图分类号:S641.3 文献标志码:A 文章编号:1673-2871(2023)05-084-07
Melatonin regulates physiological characteristics of pepper seedlings in response to salt stress
CHEN Nan ZHANG Wei ZHANG Xiaoming
(1.Wushan County Productivity Promotion Center, Wushan 741300, Gansu, China; 2.Wushan Agricultural Technology Extension Center, Wushan 741300, Gansu, China; 3.Wushan County Agricultural and Rural Bureau, Wushan 741300, Gansu, China)
Abstract: In order to explore the regulatory effect of melatonin on pepper leaf growth under salt stress, pepper seedlings of Tian Xian 1712 and Hang Jiao 4 were used as experimental materials, and different concentrations(10, 50, 100 μmol·L-1)of melatonin spraying treatment were set, the photosynthetic parameters, chlorophyll content, PSⅡ light energy conversion efficiency, sodium and potassium ion content of pepper seedlings under salt stress(200 mmol·L-1NaCl)were measured. Combined with principal component analysis, the effects of melatonin treatment on physiological characteristics of pepper seedling leaves under salt stress were studied. The results showed that the three concentrations of melatonin significantly alleviated the adverse effects of salt stress on pepper seedlings, and increased the net photosynthetic rate, stomatal conductance, intercellular carbon dioxide concentration, transpiration rate, chlorophyll content and PSⅡ light energy conversion efficiency of pepper seedlings under salt stress;It decreased the content of sodium ion and increased the content of potassium ion under salt stress: The malondialdehyde content, electrolyte leakage rate, hydrogen peroxide content and proline content were reduced. 50 μmol·L-1melatonin treatment had the most significant alleviating effect. It can be concluded that exogenous melatonin can slow down the damage of salt stress on pepper seedlings and improve the salt resistance of pepper seedlings, 50 μmol·L-1melatonin treatment was the optimal concentration to regulate the leaf growth of pepper seedlings under salt stress.
Key words: Pepper seedlings; Melatonin; Salt stress; Photosynthetic characteristics
园艺作物是人体营养中碳水化合物、矿物质、微量营养素、蛋白质、维生素、脂肪、纤维、有机酸、色素和抗氧化剂的主要来源[1]。盐胁迫抑制植物生长发育,是对园艺作物最具破坏性的非生物威胁之一[2]。据联合国粮农组织统计,全球范围内,4500万hm2耕种土地(占世界总灌溉面积的19.5%)已经受到盐胁迫的破坏。盐胁迫下植物体内离子稳态被破坏,引起钠离子含量升高,钾离子含量降低[3-4];植物体内活性氧累积,造成氧化胁迫,导致细胞膜受损,表现为丙二醛含量和电解质渗透率升高[5-6]。此外,盐胁迫引起的渗透势改变也会导致脯氨酸含量升高[7]。光合作用是盐胁迫下受影响最严重的过程之一[8]。盐分通过降低叶绿素和类胡萝卜素含量,扭曲叶绿素、质体超微结构和PSII系统,降低气孔导度,从而阻碍光合作用、蒸腾作用和气体交换[9]。
褪黑素(N-乙酰-5-甲氧基色胺,MT)于1958年首次被鉴定,是一种具有吲哚结构的低分子激素,存在于从原核生物到真核生物,从动物到植物的所有领域[10-11]。在植物中,褪黑素与生长、生根和光合作用过程有关,在植物对干旱、高温、盐碱等胁迫的反应中起着重要作用[12]。褪黑素可以通过两种途径提高植物对盐胁迫的耐受性:一是通过直接清除活性氧等过程;另一种是通过间接加工,通过提高抗氧化酶活性、光合效率、代谢物含量和调节应激相关转录因子[13]。前人研究表明,褪黑素在盐胁迫下维持Na+/K+稳态方面也起着重要作用。外源MT可提高盐胁迫下水稻或玉米的K+/Na+比值,并可促进苹果中MdNHX1/MdAKT1和油菜中SOS2/NHX1的表达[14-16]。
据前人报道,在一些园艺作物中,褪黑素处理改善了盐胁迫下植物光合作用和其他相关机制。在西瓜中,褪黑素(50~500 μmol·L-1)的预处理提高了净光合速率、叶绿素含量和气孔功能[17];黄瓜的光系统II活性和叶绿素含量也有所增加[18];在番茄中也有类似的观察结果[19]。辣椒(Capsicum annuum L.)作为园艺作物,在中国蔬菜设施栽培中占有重要地位[20],它富含维生素B、胡萝卜素以及钙、铁等矿物质,是人们日常生活中不可或缺的蔬菜[21]。辣椒的生长发育对高温、盐分和干旱胁迫十分敏感[22],土壤盐渍化将导致辣椒根系不发达、植株矮小、病害加重,進而降低果实的产量,严重影响了辣椒的经济效益[23]。褪黑素可以缓解盐胁迫对大多数作物的不利影响,然而目前关于褪黑素提高辣椒盐胁迫耐受能力的相关研究还未见报道。因此,笔者以天线1712和航椒4号两个品种辣椒幼苗为材料,进行了褪黑素对盐胁迫下辣椒幼苗生理特性影响的探究试验,以期为利用褪黑素提高辣椒等园艺作物耐盐性提供理论依据。
1 材料与方法
1.1 材料
供试辣椒品种是天线1712和航椒4号,种子由天水绿鹏农业科技有限公司提供。试验于2021年9月在天水市农业科学研究院进行。挑选籽粒饱满、大小均匀和无杂质的种子进行催芽,种子催芽后播种在小花盆中(直径×高=15 cm×12 cm),每盆播种1粒,每个品种播种32盆,育苗基质为V蛭石∶V草炭=1∶1,定植后60 d进行处理,每个处理设置3个生物学重复。
1.2 方法
参照张俊峰[24]的方法进行褪黑素喷施,参照刘微等[25]的结果进行盐胁迫浓度设置。试验共设计5个处理,分别为CK:清水对照;T1:200 mmol·L-1NaCl;T2:10 μmol·L-1MT+200 mmol·L-1NaCl;T3:50 μmol·L-1MT+200 mmol·L-1NaCl;T4:100 μmol·L-1MT+200 mmol·L-1NaCl,各处理随机选择已播种的5盆辣椒进行试验。处理时首先在夜晚避光条件下对辣椒幼苗进行不同浓度褪黑素喷洒,直至叶片表面有小水珠落下,为避免褪黑素液体挥发,用袋子罩住营养钵,连续喷施5 d,用清水喷施对照组辣椒幼苗。第6天开始对T1~T4进行盐胁迫处理,每次浇灌100 mL的NaCl(200 mmol·L-1)溶液,每3 d浇灌1次处理液,连续处理15 d,最后1次处理完成后第5天测定光合指标,取功能叶片(从上向下完全展开的第3~4片叶)置于液氮中,贮藏于低温冰箱,后续进行其他指标测定。
1.3 测定项目与方法
采用TARGAS-1(美国PP systems)便携式光合仪测定光合参数,主要测定参数为净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)。叶绿素含量测定方法参考刘微等[25]的报道,采用95%提取辣椒幼苗叶绿素,利用分光光度法测定665、649 nm处吸光值,计算叶绿素a、叶绿素b含量,叶绿素总含量=叶绿素(a+b)。采用叶绿素荧光仪(北京Yaxin-1161G)测定PSⅡ光能转换效率,测定前利用暗适应夹子将叶片暗处理15 min,然后进行测定。叶片钠钾离子含量测定参考偶春等[26]的报道,测定前将叶片置于120 ℃烘箱中处理30 min,研磨为粉末采用硝酸-高氯酸混合液(V浓硝酸∶V浓高氯酸=5∶1)消煮处理,利用原子吸收分光光度计测定钠钾离子含量。采用硫代巴比妥酸法[27]测定丙二醛含量,采用电导法[27]测定电解质渗透率,采用酸性茚三酮法[27]测定脯氨酸含量。过氧化氢含量测定参考张俊峰[24]和Fya等[28]方法,新鲜样品用2 mL冷丙酮均质,并在6000 r·min-1和4 ℃下离心20 min。1 mL上清液与0.1 mL 5% TiSO4(溶解于98% H2SO4)和0.2 mL NH4OH溶液混合,然后在3000 r·min-1下离心,并丢弃上清液。沉淀物用5 mL 2 mol·L-1H2SO4,在415 nm处读取吸光度值。
1.4 数据分析
采用Excel 2016软件进行数据处理及作图,采用SPSS 19.0软件进行方差分析。
2 结果与分析
2.1 褪黑素对盐胁迫下辣椒幼苗光合参数的影响
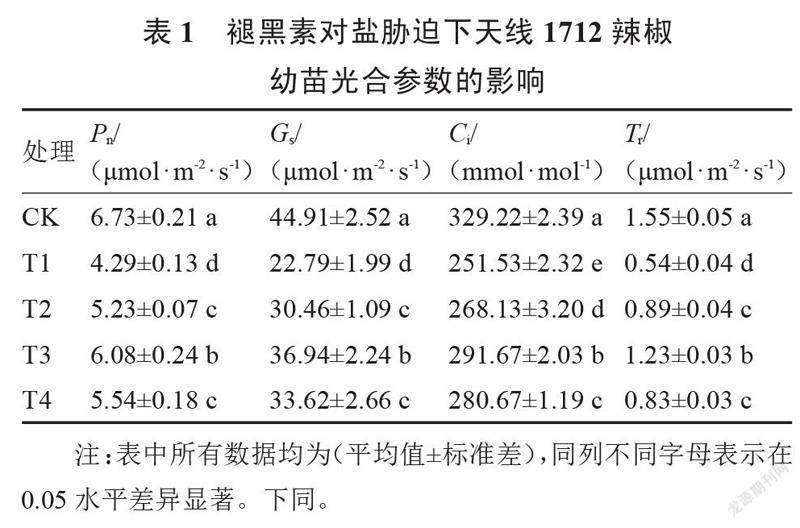
从表1和表2可以看出,盐胁迫显著降低了2个品种辣椒幼苗的光合参数,3个浓度的褪黑素处理均显著提高了盐胁迫下4个光合参数指标。盐胁迫下(T1处理)天线1712的Pn、Gs、Ci和Tr,分别较CK降低了36.26%、49.25%、23.60%和65.16%;与T1处理相比,T3处理下4个光合参数指标分别显著提高了41.72%、62.09%、15.96%和127.78%,且显著高于T2和T4处理。盐胁迫下(T1处理)航椒4号辣椒幼苗的Pn、Gs、Ci和Tr,分别较CK降低了33.49%、40.92%、32.77%和49.25%;与T1处理相比,T3处理下4个光合参数指标分别显著提高了36.32%、54.54%、31.29%和62.09%,且显著高于T2和T4处理。
2.2 褪黑素对盐胁迫下辣椒幼苗叶绿素含量的影响
从表3和表4可以看出,盐胁迫显著降低了2个辣椒品种幼苗的叶绿素含量,3个浓度褪黑素处理明显抑制了盐胁迫引起的叶绿素含量的降低。盐胁迫下(T1处理)天线1712幼苗叶绿素a、叶绿素b和叶绿素总含量分别较CK显著降低了30.98%、41.04%和32.31%;与T1处理相比,T3处理下叶绿素a、叶绿素b和叶绿素总含量分别显著提高了25.45%、58.82%和29.30%,且显著高于T2和T4处理。盐胁迫下(T1处理)航椒4号幼苗叶绿素a、b和总含量分别较CK显著降低了35.00%、45.54%和36.82%。与T1处理相比,T3处理叶绿素a、b和叶绿素总含量分别显著提高了27.26%,66.38%和32.44%,且显著高于T2和T4处理。
2.3 褪黑素对盐胁迫下辣椒幼苗PSⅡ光能转换效率的影响
由表5可知,盐胁迫处理显著降低了2个辣椒品种幼苗的PSⅡ光能转换效率,盐胁迫下(T1处理)天线1712和航椒4号分别较CK显著降低了17.50%和16.67%。3个浓度褪黑素处理显著缓解了盐胁迫引起的Fv/Fm值的降低,T3处理下幼苗Fv/Fm值显著高于T2和T4处理。与T1处理相比,T3处理下两个品种幼苗Fv/Fm分别显著提高了15.15%和18.46%。
2.4 褪黑素对盐胁迫下辣椒幼苗细胞膜的影响
从图1可以看出,盐胁迫处理下2个辣椒品种幼苗丙二醛含量和电解质渗透率显著升高,3个浓度褪黑素处理显著抑制了盐胁迫诱导的丙二醛含量和电解质渗透率升高。盐胁迫下(T1处理)天线1712和航椒4号幼苗丙二醛含量分别较CK显著提高了66.72%和59.77%。与T1处理相比,T3处理下2个品种丙二醛含量分别显著降低了37.57%和36.91%,且均显著低于T2和T4处理。
盐胁迫下(T1处理)天线1712和航椒4号幼苗的电解质渗透率分别较CK显著提高了143.84%和166.83%。与T1处理相比,T3处理下2个品种电解质渗透率分别显著降低了42.26%和47.01%,且显著低于T2和T4处理。
2.5 褪黑素對盐胁迫下辣椒幼苗钠钾离子含量的影响
由表6和7可知,盐胁迫处理提高了2个辣椒品种幼苗叶片钠离子含量,降低了钾离子含量,导致钾钠比降低;3个浓度褪黑素处理显著抑制了盐胁迫引起的钠离子的升高和钾离子的降低。与CK相比,T1处理天线1712幼苗钠离子含量显著提高261.29%,钾离子含量显著降低了36.82%。与T1处理相比,天线1712幼苗T3处理下钠离子含量显著降低了34.82%,钾离子含量显著提高了46.65%。与CK相比,航椒4号幼苗(T1处理)钠离子含量显著提高288.41%,钾离子含量显著降低了41.05%;与T1处理相比,T3处理下航椒4号幼苗钠离子含量显著降低了42.54%,钾离子含量显著提高了60.72%。
2.6 褪黑素对盐胁迫下辣椒幼苗过氧化氢和脯氨酸含量的影响
由图2所示,盐胁迫下2个辣椒品种幼苗过氧化氢和脯氨酸含量均显著升高。天线1712和航椒4号幼苗(T1处理)过氧化氢含量分别较CK显著提高了103.16%和100.89%。与T1处理相比,T3处理2个品种过氧化氢含量分别显著降低了38.43%和40.92%,且低于T2和T4处理。
天线1712和航椒4号幼苗脯氨酸含量(T1处理)分别较CK显著提高了62.02%和57.81%。与T1处理相比,T3处理2个品种脯氨酸含量分别显著提高了98.26%和87.17%,且显著高于T2和T4处理。
2.7 褪黑素对盐胁迫下辣椒幼苗叶片生理特性的主成分分析
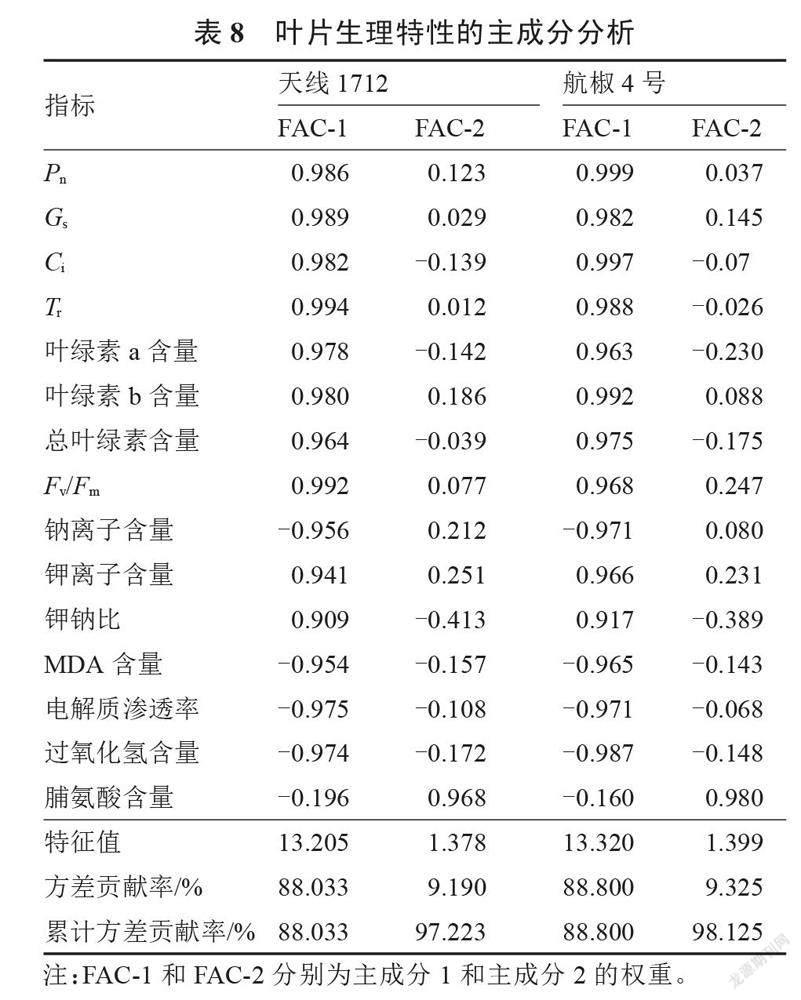
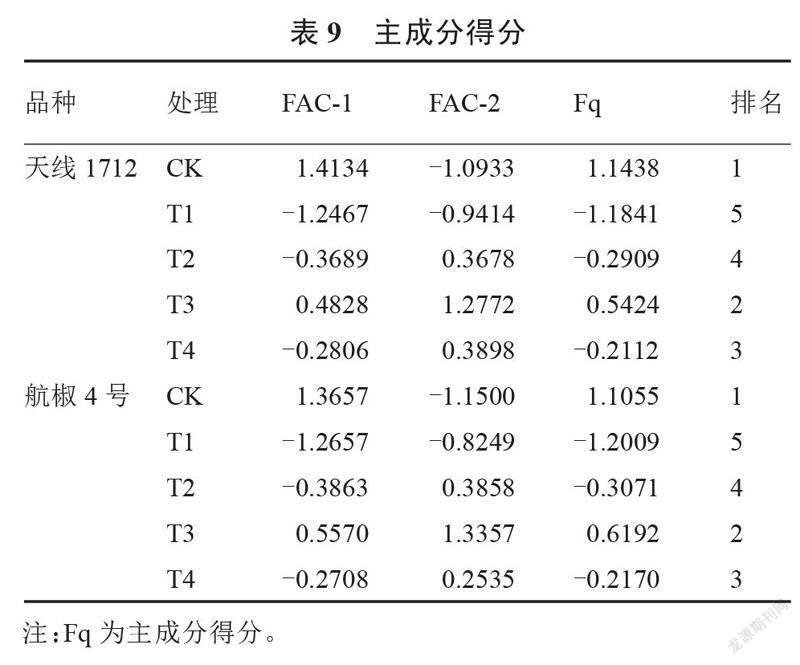
对褪黑素处理的2种盐胁迫辣椒幼苗的叶片生理特性进行主成分分析。由表8所示,两种辣椒中均提取出特征值大于1的主成分2个,其中天线1712中2个主成分的特征值分别为13.205和1.378,累计方差贡献率为97.223%;航椒4号中2个主成分的特征值分别为13.320和1.339,累计方差贡献率为98.125%。两者累计贡献率均高于85%,说明提取出的主成分可以较好地解释原有品质指标的所有信息。其中,Pn、Gs、Ci和Tr在2个品种的主成分1中均占有较高的正向量值,钠离子含量、MDA含量、电解质渗透率和过氧化氢含量在2个品种的主成分1中均占有较高的负向量值;脯氨酸含量在2个品种的主成分2中占有较高的正向量值,钾钠比在2个品种的主成分2中占有较高的负向量值。对2个品种的主成分得分进行计算,结果见表9。由表9所示,2个品种的排名均表现为CK>T3>T4>T2>T1,CK处理下2个品种的得分最高,T1处理下2个品种的得分最低,T3处理排名第二,说明正常生长过程中叶片的生理特性优于盐胁迫处理,进行褪黑素处理后(T2、T3和T4处理)的辣椒苗生理特性均优于T1,T3处理效果最为显著。由此可以得出,褪黑素处理可以增强辣椒苗的抗盐性,喷施50 μmol·L-1褪黑素处理效果最佳。
3 讨论与结论
光合作用是高等植物产生能量的最重要的物理和化学过程,对盐胁迫非常敏感[8]。盐胁迫下中椒4号辣椒幼苗的光合参数(Pn、Gs、Tr、Ci)与对照相比都有较大幅度下降[29]。张玲等[30]发现盐胁迫下耐盐品种(强丰7301)和盐敏感品种(秋艳)辣椒幼苗的Fv/Fm和叶绿素含量降低。笔者的研究结果表明,与对照相比,T1处理的Fv/Fm值显著下降,说明盐胁迫能够降低辣椒幼苗的Fv/Fm值。本试验研究表明,与对照相比,盐胁迫下天线1712和航椒4号幼苗的光合参数(Pn、Gs、Tr、Ci)显著降低,叶绿素含量显著下降,出现该现象的原因可能是褪黑素在应对植物非生物胁迫时,它重要的作用机制是通过减少叶绿素降解来提高光合活性的[31]。褪黑素预处理提高了低温胁迫下甜瓜植株的叶绿素含量和光合速率[32],施用50~150 μmol·L-1褪黑素缓解了盐分胁迫下黄瓜净光合速率的下降,增加了总叶绿素含量,显著提高了黄瓜的光合能力[33]。笔者试验发现,3个浓度褪黑素(10、50、150 μmol·L-1)褪黑素预处理缓解了盐胁迫对天线1712和航椒4号幼苗的抑制作用,显著提高了盐胁迫下光合参数(Pn、Gs、Tr、Ci)、Fv/Fm值和叶绿素含量。盐胁迫下植物体内水势升高、气孔导度下降可以减少水分流失,褪黑素可以改善气孔的功能,使植物在盐胁迫等渗透胁迫下重新开放气孔[34],适宜浓度的褪黑素可以改善气孔的功能,使植物在盐胁迫等渗透胁迫下重新开放气孔,提高Gs和Ci,这与笔者试验所得结果相一致。
细胞膜对物质具有选择透过性,是维持细胞内环境和代谢正常进行的基础。在逆境胁迫下,植物细胞内产生过量的自由基会使膜脂发生过氧化作用从而破坏膜系统[35]。此外,在极端盐胁迫下,黄瓜[36]、番茄[37]和柑橘[38]经褪黑素处理后,H2O2和O2·浓度急剧降低,活性氧清除能力增强。MDA作为膜脂过氧化的代谢产物,反映了细胞的受损程度。本试验中T1处理中天线1712和航椒4号幼苗的H2O2含量较对照显著增加,MDA含量和电解质渗透率显著提高。T3处理降低了盐胁迫下2个品种辣椒幼苗H2O2含量、MDA含量和电解质渗透率,这表明褪黑素预处理可以通过降低辣椒幼苗H2O2含量缓解细胞膜氧化损伤。
盐胁迫下,Na+进入胞质溶胶引起质膜去極化,导致K+持续流出。Na+的积累和K+的外流导致细胞质中Na+/K+的升高,进而对植物造成严重损害[39-40]。盐胁迫下天线1712和航椒4号幼苗的Na+含量升高,K+含量降低。褪黑素具有维持植物系统内稳定的离子浓度的作用,在玉米[14]、水稻[16,28]、香椿[26]和甜瓜[41]中都发现褪黑素处理能减缓植株对Na+的吸收积累,调节植物体内离子稳态,缓解因钠离子积累导致的离子毒害。深入研究表明,这是由于褪黑素能够调控离子转运相关基因的表达[14-16]。笔者的研究结果表明,适宜浓度的褪黑素处理能够降低盐胁迫下天线1712和航椒4号幼苗的Na+含量,并提高K+含量。
综上,褪黑素处理显著缓解了盐胁迫对辣椒幼苗的不利影响,提高了盐胁迫下辣椒幼苗的净光合速率、气孔导度、胞间二氧化碳浓度、蒸腾速率、叶绿素含量、PSⅡ光能转换效率和脯氨酸含量;降低了盐胁迫下幼苗的钠离子含量、丙二醛含量、电解质渗透率和过氧化氢含量,可以减缓盐胁迫对辣椒幼苗产生的伤害,提高辣椒幼苗的抗盐性,其中,50 μmol·L-1褪黑素处理为调控盐胁迫下辣椒幼苗叶片生长的最适浓度。
参考文献
[1] IMAHORi Y.Role of ascorbate peroxidase in postharvest treatments of horticultural crops[J].Oxdiative Damage to Plants,2014:425-451.
[2] CHANG B W,YANG L,CONG W W,et al.The improved resistance to high salinity induced by trehalose is associated with ionic regulation and osmotic adjustment in catharanthus roseus[J].Plant Physiology and Biochemistry,2014,77:140-148.
[3] ISAYENKOV S V,MAATHUIS F J M.Plant salinity stress:many unanswered questions remain[J].Frontiers In Plant Science,2019,10:80.
[4] CAMBRIDGE M L,ZAVALA-Perez A,CAWTHRAY G R,et al.Effects of high salinity from desalination brine on growth,photosynthesis,water relations and osmolyte concentrations of seagrass Posidonia australis[J].Marine Pollution Bulletin,2017,115(1/2):252-260.
[5] WANI A S,AHMAD A,HAYAT S,et al.Epibrassinolide and proline alleviate the photosynthetic and yield inhibition under salt stress by acting on antioxidant system in mustard[J].Plant Physiology and Biochemistry,2019,135:385-394.
[6] KAYA C,AKRAM N A,ASHRAF M,et al.Exogenous application of humic acid mitigates salinity stress in maize(Zea mays L.)plants by improving some key physico-biochemical attributes[J].Cereal Research Communications,2018,46(1):67-78.
[7] ARIF Y,SINGH P,SIDDIQUI H,et al.Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance[J].Plant Physiology and Biochemistry,2020,156:64-67.
[8] YANG Z,LI J L,LIU L N,et al.Photosynthetic regulation under salt stress and salt-tolerance mechanism of sweet sorghum[J].Frontiers in Plant Science,2020,10:1722-1733.
[9] PAN T,LIU M,KRESLAVSKI V D,et al.Non-stomatal limitation of photosynthesis by soil salinity[J].Critical Reviews in Environmental Science and Technology,2020,51(7):1-35.
[10] ARNAO M B,HERNANDEZ R J.Melatonin:plant growth regulator and/or biostimulator during stress? [J] Trends in Plant Science,2014,19(12):741-808.
[11] LERNER A B,CASE J D,TAKAHASHI Y,et al.Isolation of melatonin,the pineal gland factor that lightens melanocytes[J].Journal of the American Chemical Society,1958,80:2587-2587.
[12] ARNAO M B,HERNANDEZ R J.Functions of melatonin in plants:A review[J].Journal of Pineal Research,2015(2):133-150.
[13] LI J,LIU J,ZHU T,et al.The role of melatonin in salt stress responses[J].International Journal of Molecular Sciences,2019,8,20(7):1735-1749.
[14] JIANG C,CUI Q,FENG K,et al.Melatonin improves antioxidant capacity and ion homeostasis and enhances salt tolerance in maize seedlings[J].Acta Physiologiae Plantarum,2016,38:82-90.
[15] LI C,WANG P,WEI Z,et al.The mitigation effects of exogenous melatonin on salinity-induced stress in Malus hupehensis[J].Journal of Pineal Research,2012(3):298-306.
[16] LI X J,YU B J,CUI Y Q,et al.Melatonin application confers enhanced salt tolerance by regulating Na+ and Cl? accumulation in rice[J].Plant Growth Regulation,2017,83:441-454.
[17] MANDAL M K,SUREN H,WARD B,et al.Differential roles of melatonin in plant-host resistance and pathogen suppression in cucurbits[J].Journal of Pineal Research,2018,65(3):e12505.
[18] WANG L,LIU J L,WANG W,et al.Exogenous melatonin improves growth and photosynthetic capacity of cucumber under salinity-induced stress[J].Photosynthetica,2016,54(1):19-27.
[19] YIN Z,LU J Z,MENG S,et al.Exogenous melatonin improves salt tolerance in tomato by regulating photosynthetic electron flux and the ascorbate-glutathione cycle[J].Journal of Plant Interactions,2019,14(1):453-463.
[20] 牛彩霞,郁繼华,张韵,等.钠盐对辣椒种子萌发和幼苗生长的影响[J].甘肃农业大学学报,2006,41(1):34-38.
[21] GUO M,LIU J H,MA X,et al.Genome-wide analysis of the Hsp70 family genes in pepper(Capsicum annuum L.)and functional identification of CaHsp70-2 involvement in heat stress[J].Plant Science.2016,252:246-256.
[22] GUO W L,CHEN R G,DU X H,et al.Reduced tolerance to abiotic stress in transgenic arabidopsis overexpressing a Capsicum annuum multiprotein bridging factor 1[J].BMC Plant Biology,2014,20(14):138-150.
[23] 郑佳秋,郭军,梅燚,等.辣椒种子萌发和幼苗生理特性对盐胁迫的响应[J].江苏农业科学,2016,44(11):182-186.
[24] 张俊峰.基于外源褪黑素作用的辣椒耐低温弱光生理及分子响应机理研究[D].兰州:甘肃农业大学,2020.
[25] 刘微,李佳薇,徐若瑄,等.NaCl胁迫对辣椒种子萌发及幼苗生长的影响[J/OL].分子植物育种,2021:1-14.https: //kns.cnki.net/kcms/detail/46.1068.s.20211029.1404.004.html.
[26] 偶春,张敏,姚侠妹,等.褪黑素对盐胁迫下香椿幼苗生长及离子吸收和光合作用的影响[J].西北植物学报,2019,39(12):2226-2234.
[27] 陈刚,李胜.作物生理学实验[M].北京:高等教育出版社,1999.
[28] FYA B,HWA B,YDAB C,et al.Melatonin enhances Na+/K+ homeostasis in rice seedlings under salt stress through increasing the root H+-pump activity and Na+/K+ transporters sensitivity to ROS/RNS[J].Environmental and Experimental Botany,2021,182:104328-104261.
[29] 张志刚,尚庆茂.辣椒幼苗叶片光合特性对低温、弱光及盐胁迫3重逆境的响应[J].中国生态农业学报,2010,18(1):77-82.
[30] 张玲,王华,周静,等.NaCl胁迫对两个辣椒品种幼苗叶绿素荧光参数等生理特性的影响[J].浙江农业学报,2017,29(4):597-604.
[31] SANTOSH K B,PRIANKA H.Melatonin plays multifunctional role in horticultural crops against environmental stresses:A review[J].Environmental and Experimental Botany,2020,176:104063-104123.
[32] ZHANG Y,YANG S,CHEN Y.Effects of melatonin on photosynthetic performance and antioxidants in melon during cold and recovery[J].Biologia Plantarum,2017,61(3):571-578.
[33] WANG L,LIU J,WANG W,et al.Exogenous melatonin improves growth and photosynthetic capacity of cucumber under salinity-induced stress[J].Photosynthetica,2016,54(1):19-27.
[34] YE J,WANG S,DENG X,et al.Melatonin increased maize(Zea mays L.)seedling drought tolerance by alleviating drought-induced photosynthetic inhibition and oxidative damage[J].Acta Physiologiae Plantarum,2016,38(2):1-13.
[35] 國海燕.褪黑素引发种子处理对冬小麦光合特性及生长发育的影响[D].陕西杨凌:西北农林科技大学,2017.
[36] ZHANG N,ZHANG H J,SUN Q Q,et al.Proteomic analysis reveals a role of melatonin in promoting cucumber seed germination under high salinity by regulating energy production[J].Science Reports,2017,29,7(1):503.
[37] YIN Z,LU J Z,MENG S,et al.Exogenous melatonin improves salt tolerance in tomato by regulating photosynthetic electron flux and the ascorbate-glutathione cycle[J].Journal of Plant Interactions,2019,14(1):453-463.
[38] KOSTOPOULOU Z,THERIOS I,ROUMELIOTIS E,et al.Melatonin combined with ascorbic acid provides salt adaptation in Citrus aurantium L.seedlings[J].Plant Physiology and Biochemistry,2015,86:155-165.
[39] MUNNS R,TESTER M.Mechanisms of salinity tolerance[J].Annual Review Plant Biology,2008,59:651-681.
[40] SHABALA S,DEMIDCHIK V,SHABALA L,et al.Extracellular Ca2+ ameliorates NaCl-induced K+ loss from arabidopsis root and leaf cells by controlling plasma membrane K+-permeable channels[J].Plant Physiology,2006,141(4):1653-1665.
[41] 高青海,郭远远,吴燕,等.盐胁迫下外源褪黑素和Ca2+对甜瓜幼苗的缓解效应[J].应用生态学报,2017,28(6):1925-1931.
