寒地水稻龙粳31耐冷特性及油菜素内酯的耐冷调节作用
2023-05-26曾维康魏嘉琪过一驰程一珊姜树坤赵明辉耿丽娟郭志富
高 洋, 曾维康, 魏嘉琪, 过一驰, 程一珊, 姜树坤, 赵明辉, 耿丽娟, 郭志富
(1.沈阳农业大学生物科学技术学院, 沈阳 110866;2.黑龙江省农业科学院耕作栽培研究所, 哈尔滨 150086;3.沈阳农业大学水稻研究所, 沈阳 110866; 4.沈阳市食品药品检验所, 沈阳 110000)
低温冷害是常见的非生物逆境之一,对水稻(OryzasativaL.)的生长发育影响巨大,是限制水稻产量稳定和提高的主要环境因素[1]。寒地水稻因特殊的选育和生长环境,具备更好的低温耐受潜力和基础[2],其主要在东北稻区尤其是黑龙江稻区种植,而黑龙江稻区作为我国最大的商品粳稻生产基地,水稻种植面积和产量均为全国第一,近三年水稻种植面积约占全国粳稻面积的50%[3],对保障国家粮食安全具有重要的战略意义。
多年来,针对高纬度低积温以及冷害频发等问题,东北地区水稻育种工作者选育出很多熟期早、抗性强、产量高的寒地水稻品种,为东北稻区水稻产量的稳定和提高奠定了基础,同时为深入开展水稻耐冷基础理论研究提供了理想的研究材料。龙粳、龙稻、绥粳、垦稻等系列品种,均在东北稻区有一定的种植面积[4]。在众多寒地水稻品种中,龙粳31因其高产、优质、抗逆性强等显著特点,多年来一直是全国粳稻种植面积最大的品种,已累计推广超过130万hm2。目前,针对龙粳31的基础理论研究主要集中在栽培措施的优化上,如种植密度调整、肥料配比优化、旱直播条件摸索等方面,最近也出现了龙粳31遗传基础分析以及利用基因编辑技术对其进行品质改良探索的相关报道[5-8]。龙粳31作为种植面积最大的寒地水稻品种,有很强的耐冷性,但目前对于龙粳31低温耐受表型特征及生理生化等方面的研究较为少见,限制了对寒地水稻耐冷机理的深入了解。
油菜素内酯(Brassinolides,BRs)是甾醇类植物激素中最具活性的一种,可有效提高植物的抗冷性。研究表明,外源喷施BRs对于水稻不同时期耐冷性的提高均有显著的促进作用[9-12]。目前,BRs处理与寒地水稻耐冷性关系方面的研究尚未见报道。基于以上分析,本研究以耐冷性较强的寒地水稻龙粳31和冷敏感水稻9311为研究材料,在芽期和苗期分别鉴定其在冷处理以及低温和BRs联合处理下的耐冷表型性状变化,并测定耐冷相关生理生化指标及内源激素含量的变化情况,为寒地水稻耐冷生理基础及BRs途径下水稻耐冷调控机制的明确提供参考。
1 材料与方法
1.1 试验材料
本研究以寒地水稻龙粳31和冷敏感水稻9311为试验材料。龙粳31由黑龙江农科院水稻研究所提供,9311由沈阳农业大学水稻研究所提供。
1.2 试验方法
1.2.1低温胁迫及BRs处理后低温发芽率测定
试验分为三个处理,常温对照为25 ℃(ck)、低温处理温度为17 ℃、联合处理为BRs(0.01 μmol/L)+17 ℃低温处理。选取50粒健康饱满的水稻种子,在45 ℃烘箱处理2 d打破休眠后,分别使用蒸馏水和BRs(0.01 μmol/L)进行24 h浸种处理。将浸种后的种子置于放有湿润滤纸的培养皿中分别放入25 ℃和17 ℃光照培养箱中处理7 d。
1.2.2低温胁迫及BRs处理后苗期表型的测定
试验分为3个处理,常温25 ℃、4 ℃低温处理、联合处理为BRs(0.1 μmol/L)+4 ℃低温处理。选取健康饱满的种子,在45 ℃烘箱里放置3 d打破种子休眠后,放在垫有湿润滤纸的玻璃培养皿里,于35 ℃恒温箱中黑暗催芽72 h,种在混合基质土(V基质土∶V蛭石∶V珍珠岩=2∶2∶1)的栽培盆(7 cm×7 cm×7 cm),在25 ℃培养室内生长21 d,长至三叶期,将4 ℃、4 ℃+BRs(处理前24 h,用0.1 μmol/L BRs进行根际灌溉和叶面喷施)两种处理,置于10 ℃低温培养箱中处理7 d后,常温恢复7 d,将表型与ck进行比较。
1.2.3低温胁迫及BR处理后苗期叶片生理生化指标测定
将前述苗期低温和联合处理的水稻幼苗按0、1、3、5、7 d进行叶片取样,分别进行脯氨酸(Pro)含量、丙二醛(MDA)含量、相对电导率、超氧化物歧化酶(SOD)活性测定[14-15]。
1.2.4低温胁迫及BR处理后苗期叶片生理生化指标测定
苗期低温和联合处理3 d后,对幼苗叶片进行取样,使用预冷却的研钵将0.5 g样品匀浆,然后在4 mL提取溶液中进行超声辅助提取45 min。12 000 r/min离心10 min后,收集上清液,重复抽提一次。用2 mL甲醇和2 mL甲酸(1 mol/L)预活化HLB和MCX柱。然后,将500 μL溶液通过0.22 μm过滤器过滤,并转移到2 mL的LC-MS瓶中进行UPLC-MS/MS分析[16]。
2 结果与分析
2.1 低温胁迫及BRs处理对水稻发芽率的影响
由图1可知,龙粳31的ck、低温处理和BRs+低温处理的发芽率分别为100.00%、93.33%、97.33%,而9311的ck、低温处理和BR+低温处理的发芽率分别为100.00%、82.67%、86.00%,冷处理的龙粳31和9311发芽率分别降低6.67%和17.33%,证明龙粳31的低温发芽率要高于9311,具备更好的芽期耐冷基础。BRs浸泡后进行冷处理的龙粳31和9311发芽率分别降低4.29%和4.03%,证明低温条件下,BRs对水稻种子发芽率有一定的抑制作用。
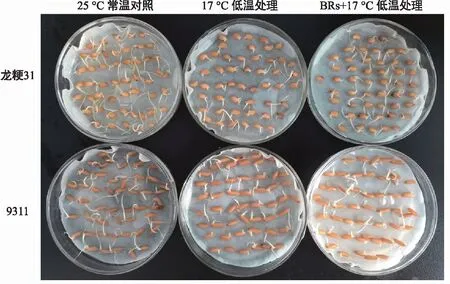
图1 低温发芽率表型Fig.1 Germination rate phenotype under cold temperature

注:每组材料从左到右分别为25 ℃常温对照,4 ℃冷处理,BRs+4 ℃冷处理。A为冷处理3 d;B为冷处理7 d;C为恢复生长3 d;D为恢复生长7 d。图2 苗期耐冷表型Fig.2 Phenotype with cold tolerance at seedling stage
2.2 低温胁迫及BRs处理后苗 期表型变化
苗期耐冷表型鉴定结果表明,冷处理7 d后,龙粳31和9311的生长均受到明显的抑制。当恢复生长3 d时,9311出现枯萎现象,而龙粳31长势明显好于9311,当恢复生长7 d时,9311已经干枯死亡,龙粳31仍然长势良好,说明龙粳31具有更强的苗期耐冷性。与单独冷处理相比,BRs+冷处理后明显缓解了低温对龙粳31的生长限制,而对于9311的缓解作用并不明显(图2)。
2.3 低温胁迫及BRs处理对水稻生理指标的影响
为了进一步探究寒地水稻耐冷生理变化,对低温及BRs处理下的SOD活性、Pro和MDA含量和相对电导率进行鉴定。常温下,龙粳31和9311幼苗的SOD活性相近,随着低温处理时间的延长,两种材料的SOD活性均呈现先升高后降低的趋势,且龙粳31的上升幅度明显高于9311。0.1 μmol/L的BR处理后,龙粳31和9311在低温胁迫后SOD活性同样呈先上升再下降的趋势,而上升幅度大于单独低温处理条件(图3 A);常温下,龙粳31幼苗的Pro含量始终高于9311;在低温处理后两材料的Pro含量均呈上升趋势,相比而言,龙粳31上升幅度更大,说明在龙粳31受低温胁迫后Pro有更多的积累,可能比9311更具耐冷生理基础。与单独低温处理相比,BRs+低温处理下龙粳31的Pro含量整体变化不大,但9311的Pro含量显著提高(图3 B);常温下,龙粳31和9311幼苗的MDA含量相近;随低温处理时间的延长,龙粳31和9311的MDA含量累积逐渐增多,且龙粳31上升程度明显低于9311,说明龙粳31可能具有更强的清除膜脂过氧化物的能力;与单独低温处理相比,BRs+低温处理后MDA含量均呈下降趋势,且9311下降幅度更为明显(图3 C);常温下,龙粳31幼苗的相对电导率和9311近似;随着低温处理时间延长,龙粳31和9311的相对电导率均显著上升,且9311较龙粳31的相对电导率上升幅度更大,说明9311的质膜对低温胁迫可能更敏感。与单独低温处理相比,BRs+低温处理可以降低龙粳31和9311的相对电导率,且对9311的作用更为明显(图3 D)。
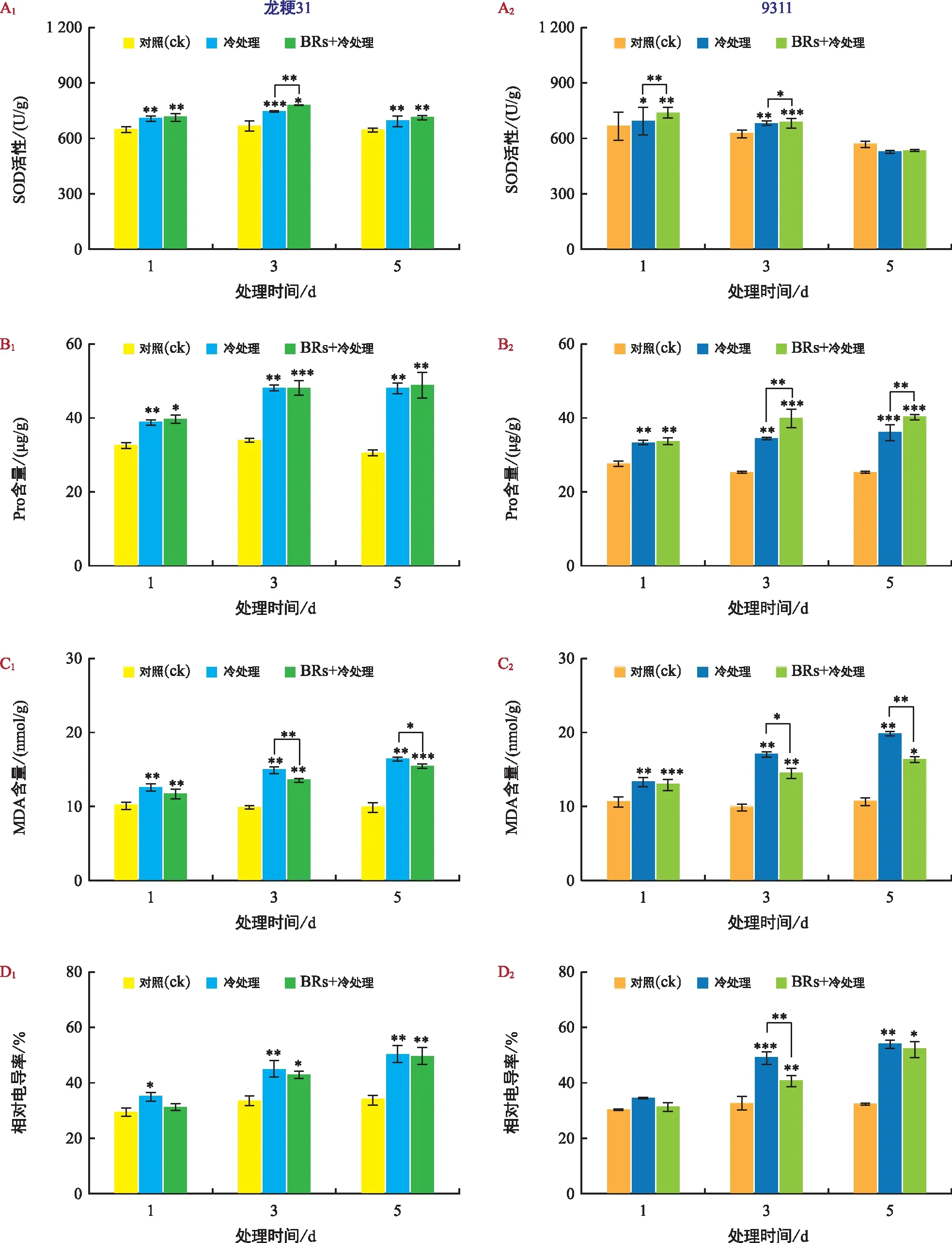
注:A为SOD活性;B为Pro含量;C为MDA含量;D为相对电导率。“*”表示p<0.05,“**”表示p<0.01,“***”表示p<0.001。下同。图3 不同处理下生理指标变化Fig.3 Changes of physiological indicators under different treatments
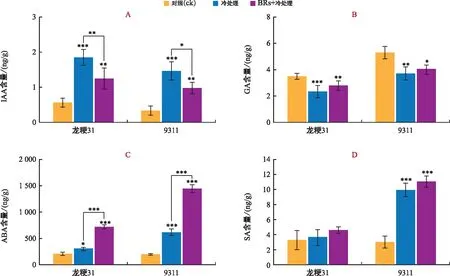
注:A为IAA含量;B为GA含量;C为ABA含量;D为SA含量;“*”表示p<0.05,“**”表示p<0.01,“***”表示p<0.001。图4 不同处理下植物激素含量变化Fig.4 Changes of the plant hormone content under different treatments
2.4 低温胁迫及BRs处理对水稻内源激素的影响
在不同处理下,对植物低温响应相关的激素IAA、GA、ABA和SA的含量分别进行测定。结果表明,常温下龙粳31幼苗IAA含量要高于9311。低温处理后,龙粳31和9311的IAA含量均显著上升,分别提高了228.44%和338.62%,9311的上升幅度更大。与单独低温处理相比,BRs+低温处理下龙粳31和9311的IAA含量分别降低了32.58%、33.56%(图4 A);GA含量的变化情况显示,常温下9311幼苗GA含量高于龙粳31。低温处理后,龙粳31和9311的GA含量均显著下降,分别下降了39.20%和30.23%,龙粳31的下降幅度更大。与单独低温处理相比,BRs+低温处理下龙粳31和9311的GA含量分别升高了20.76%和11.43%,且龙粳31上升幅度更大(图4 B);针对ABA含量而言,常温下龙粳31和9311幼苗ABA含量相近。低温处理后,龙粳31和9311的ABA含量均呈上升趋势,分别提高了44.78%和214.99%,9311上升幅度更大。与单独低温处理相比,BRs+低温处理下龙粳31和9311的ABA含量分别升高了140.40%和135.87%(图4 C);而SA含量在常温下龙粳31和9311幼苗相近。低温处理后,龙粳31和9311中SA含量均有所上升,分别提高了12.61%、229.12%,9311的上升幅度显著高于龙粳31。与单独低温处理相比,BRs+低温处理下龙粳31和9311中SA含量分别升高了10.48%和25.52%,9311上升幅度更大(图4 D)。
综合来看,IAA、ABA和SA含量均在低温处理后有所提升,且9311上升幅度更为显著。相反地,GA含量在低温处理后呈下降趋势,且龙粳31下降幅度更大。与单独低温处理相比,BRs+低温处理使ABA、SA和GA含量有所提升,9311提升的幅度更大,而BRs+低温处理后IAA呈下降趋势。这些结果表明,龙粳31内源激素受低温影响的敏感程度要低于9311,而外源施加BR会对大部分植物激素含量产生影响。
3 讨论与结论
3.1 寒地水稻表型性状与耐冷性的关系
近年来,研究人员多以低温处理后的发芽率、成苗率、发芽系数、芽期根长、苗长和第一叶长等表型指标评价寒地水稻的芽期耐冷性;以死叶率、叶片赤枯度、叶片卷曲度、根长、苗长、侧根数、干重及鲜重等表型指标进行水稻苗期耐冷性鉴定[17-19]。在本研究中,龙粳31与9311的低温发芽率有所降低,但龙粳31的降低幅度要小于9311。对于苗期而言,9311低温处理后恢复,出现了濒临枯死的状态,但龙粳31低温处理恢复后植株依旧长势良好。这些结果表明,寒地水稻龙粳31在芽期和苗期均具有更好的耐冷能力。外源施加BRs能缓解低温对水稻芽期和苗期生长的不利影响,进一步说明BRs处理对于水稻耐冷表型有一定的缓解作用。
3.2 外源BRs及低温胁迫对水稻幼苗生理生化指标的影响
研究表明,低温逆境会减少水稻叶绿素的合成,增加ROS、MDA和可溶性糖含量以及电解质渗漏率及细胞膜透性等,同时也可以显著提高SOD、POD和CAT等抗氧化酶活性,进而赋予植物对低温胁迫的抵抗力[1,14-15]。外源施加BRs可显著改善低温诱导的氧化胁迫,降低MDA含量,提高POD、SOD的活性[9-12]。本研究中,龙粳31和9311在遭遇低温胁迫期间,均可通过增加渗透调节物质含量来提高自身耐冷性。随着低温处理时间的延长,在一定时间内,Pro含量继续呈上升趋势,随着冷胁迫时间不断加长,Pro含量开始逐渐减少。BRs+低温处理条件下,0.1 μmol/L BRs处理可以影响低温下龙粳31和9311的Pro含量,但影响并不显著。低温胁迫下龙粳31和9311的MDA累积逐渐增多,相对电导率显著上升,但相对而言龙粳31的上升幅度更小,说明龙粳31清除膜脂过氧化物的能力可能更强。针对BRs+低温处理条件来分析,0.1 μmol/L的BRs处理初期,龙粳31和9311在低温下MDA的含量和相对电导率均有所降低,这也为BRs提高水稻耐冷性提供了生理层面的证据。低温胁迫初期水稻可以通过增加抗氧化酶活性来提高自身耐冷性,随着低温处理时间延长,龙粳31和9311的SOD活性均呈现先上升再下降趋势,说明植物体内酶活性存在动态平衡,超出植物承受范围,平衡遭到破坏。综合来看,龙粳31与9311相比,更具耐冷生理基础,而外源BRs处理可使水稻生理指标产生利于耐冷性提高的变化趋势。
3.3 外源BRs以及低温胁迫对水稻幼苗内源激素含量的影响
近年来,激素在植物低温胁迫应答中的作用逐渐受到重视。外源施加BRs可显著影响低温胁迫下内源激素的含量变化,有研究表明,植物激素参与植物对低温的响应,包括ABA、JA、IAA、GA、BRs、SA、C2H4等[9-12]。低温胁迫后,冬小麦(TriticumaestivumL.)ABA的含量快速增加,GA水平升高,随着冷处理时间延长,植物适应低温,ABA、CTK和GA的含量降低,SA和JA的含量开始增加[21]。然而,Du等[22]研究发现,水稻经过1 d的低温处理后,IAA含量连续5 d保持在较高水平。本研究结果与之一致。IAA的浓度易受低温胁迫的影响,其差异取决于植物种类、发育背景和生理环境。例如,低温胁迫后冬小麦中IAA含量增加,而春小麦中IAA含量保持不变[23]。低温处理小麦后,在小麦冠状组织中IAA浓度显著增加,而小麦叶片中的IAA浓度却无明显变化[24]。本研究中,龙粳31内源激素受低温影响的敏感程度要低于9311,这与表型和生理指标方面的结论相对应。
综上所述,通过对龙粳31和9311耐冷特性鉴定及BRs调节其耐冷表型和生理生化指标变化的研究,初步证明龙粳31具备更强的芽期和苗期的低温抗性,同时具备更好的耐冷生理基础。BRs处理,可对水稻芽期和苗期的多个表型指标产生倾向于耐冷性提高的正向影响,对龙粳31的影响可能更显著。但是,水稻耐冷性是一个复杂的数量性状,关于寒地水稻耐冷性与激素之间的关系需要进一步研究,外源施加BRs会对大部分植物激素含量产生影响,但BRs与水稻内源激素之间的关系仍需要进一步探讨。
