辣椒TIFY基因家族的鉴定与分析
2023-05-26苏世贤尤春生张永平韦金江江苏燕祝洪江王娅珠邓代信
吴 媛, 苏世贤, 尤春生, 张永平, 汪 淘,2, 龙 茶,韦金江,2, 江苏燕, 祝洪江, 王娅珠, 邓代信, 李 伟
(1.贵州大学农学院, 贵阳 550025; 2.贵州大学辣椒产业技术研究院, 贵阳 550025;3.贵定县农业农村局, 贵州 贵定 551300; 4.贵州省果树蔬菜工作站, 贵阳 550001;5.贵州省高等学校设施蔬菜工程研究中心, 贵阳 550025)
TIFY家族基因是植物中的一类特异性转录因子,首次在拟南芥中进行鉴定,属于GATA转录因子家族,所有TIFY基因均含有高度保守结构域(TIF[F/Y]XG),所以将其命名为TIFY基因家族[1-2]。根据TIFY中保守结构域的不同,将其分为4个亚族:TIFY、JAZ、PPD和ZML。TIFY亚族中仅含有TIFY保守结构域,该结构域含有28个氨基酸和一个保守基序TIF[F/Y]XG,该结构域二级结构预测形成α-α-β折叠[3]。JAZ亚家族中除TIFY结构域之外,在序列C端还存在一个Jas保守结构域,可以与bHLH转录因子MYC 2和MYC 3相互作用,抑制茉莉酸反应[4-5]。PPD亚家族含有TIFY结构域、PDD结构域以及PY基序缺失的Jas结构域,在拟南芥中PPD 1和PPD 2可以协调组织生长,调节叶片大小和叶片边缘的弯曲程度[6],ZML 蛋白除含有TIFY结构域外,还包含有CCT基序和GATA锌指结构域[1]。
TIFY基因家族已经在多个物种进行鉴定,如在拟南芥(Arabidopsisthaliana(L.) heynh.)中有18个TIFY成员[7],水稻(OryzasativaL.)中20个[8],番茄(SolanumlycopersicumL.)中有21个[9],马铃薯(SolanumtuberosumL.)中有21个[9],核桃(JuglansregiaL.)中有21个[10],茶树(Camelliasinensis)中有16个[11],毛果杨(Populustrichocarpa)中有25个[12]。较多研究表明,植物中TIFY成员对维持植物正常生长发育有重要作用。拟南芥中AtTIFY1可促进叶柄和下胚轴的生长[2]。在水稻中过表达OsTIFY11b增加了株高和籽粒重,与未转化植株相比,植株在抽穗期长叶,并且叶鞘和茎穗中的蔗糖含量更高[13]。研究发现,PpTIFY基因与花青素积累和JA信号传导有关,时空表达模式表明,其在叶片中表达水平较高,而在成熟期间表达水平显著降低[14]。部分研究表明,TIFY在植物对非生物胁迫的响应过程中发挥重要作用。多数苹果(MaluspumilaMill.)TIFY基因和葡萄(VitisviniferaL.)中多个TIFY基因对脱落酸和茉莉酸甲酯等有响应,但对水杨酸和乙烯没有响应[15-16]。荧光定量PCR显示,CsTIFYs在盐胁迫、冷胁迫、干旱胁迫和茉莉酸甲酯处理后反应强烈[17]。水稻中几乎所有的OsTIFY基因都对一种或多种非生物胁迫有反应,包括干旱、盐和低温等胁迫,过表达OsTIFY11a导致对盐和干旱胁迫的耐受性显著增加[8]。据报道,TaJAZ1的C末端区域与TaABI5相互作用,可以调节小麦(TriticumaestivumL.)的种子萌发和ABA反应[18]。
近年来,TIFY的鉴定分析已经在多个物种中进行,但是目前很少有关于辣椒(CapsicumannuumL.)TIFY基因家族成员的相关信息,并且辣椒中TIFY家族成员进行鉴定及基因表达模式的研究较少。辣椒是主要的蔬菜之一,富含辣椒素和维生素C等物质,广泛应用于农业、制药和食品加工等多个领域[19]。因此阐明辣椒TIFY基因对辣椒生长发育的调控具有重要意义。基于辣椒全基因组信息,鉴定辣椒TIFY基因,利用生物信息学方法对CaTIFYs的理化性质、保守基序、保守结构域、基因结构、顺式作用元件和系统发育以及基因表达模式进行可视化分析,为解析TIFY基因家族在辣椒生长发育和逆境胁迫中的生物学功能提供参考。
1 材料与方法
1.1 辣椒TIFY基因家族成员的鉴定
于茄科植物全基因组数据库(https://solgenomics.net/)下载辣椒基因组文件,拟南芥全基因组数据库TAIR(https://www.arabidopsis.org/)下载已报道的拟南芥18个TIFY转录因子序列。在TBtools软件中Blast Compare Two Seq进行Blastp比对,设定E值为10-5,进行第一次比对[20]。使用NCBI中BlastP功能进行第二次比对。在Pfam数据库[21](https://pfam.xfam.org/)下载原始隐马尔可夫模型(HMM)数据TIFY(PF 06200)文件,采用HMMER3.0查询含有TIFY保守结构域的蛋白序列,设定E值为10-5。使用NCBI中CDD数据库和SMART在线网站(http://smart.embl.de/)进行验证是否含有该结构域,去除冗余的基因,含有该结构域的基因最终确定为辣椒TIFY基因家族成员。
1.2 辣椒TIFY染色体定位、蛋白理化性质和结构特征分析
采用TBtools软件进行染色体定位分析;在Expasy (https://www.expasy.org/)中对理化性质分析[22];使用在线软件PSORT Ⅱ prediction (https://psort.hgc.jp/form2.html)进行亚细胞定位预测分析;利用SOMPA (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)预测二级结构[23];SWISS-MODEL (https://swissmodel.expasy.org/interactive)预测三级结构[24];TMHMM(https://services.healthtech.dtu.dk/service.php?TMHMM-2.0)预测跨膜结构[25]。
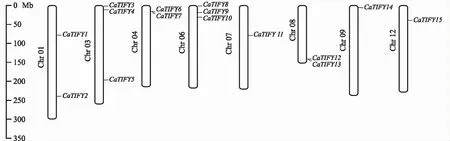
图1 CaTIFYs基因的染色体定位Fig.1 Location of CaTIFYs gene on chromosome
1.3 辣椒TIFY家族蛋白保守基序、保守结构域、基因结构和顺势作用元件分析
利用MEME在线软件(https://meme-suite.org/meme/index.html)挖掘CaTIFYs蛋白的保守基序[26];运用CDD数据库(http://www.ncbi.nlm.nih.gov/cdd/)对保守结构域进行分析;采用DSDS(http://gsds.gao-lab.org/)进行基因结构分析;采用TBtools软件提取辣椒TIFY家族成员起始密码子上游2 kb的序列,然后在PlantCare网站(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)进行顺势作用元件预测[27],最后用TBtools软件进行可视化分析。
1.4 辣椒TIFY基因家族的系统进化树构建
对拟南芥、番茄、水稻和辣椒中TIFY蛋白序列使用MEGA X软件进行多序列比对,采用邻接法构建系统发育树,bootstrap值设置为1 000[28]。
1.5 辣椒TIFY基因在不同组织、非生物胁迫和激素处理后表达模式分析
使用PepperHub[29](http://pepperhub.hzau.edu.cn/index.php)下载‘6421’辣椒CaTIFYs基因在不同组织发育时期的表达量数据,以及在多种非生物胁迫如冷胁迫、热胁迫、盐胁迫和干旱胁迫,多种激素处理如茉莉酸、脱落酸、水杨酸和赤霉素处理后,根和叶0~24 h(在0,0.5,1,3,6,12,24 h取样)内CaTIFYs基因的表达量数据。将CaTIFYs的表达量值取对数log2(FPKM)进行转换,采用TBtools软件绘制热图并进行聚类分析。
2 结果与分析
2.1 CaTIFYs家族成员鉴定、染色体定位、蛋白的理化性质和结构特征分析
基于辣椒基因组信息中共鉴定出15个TIFY成员,CaTIFYs位于8条染色体上,依据其在染色体的位置,分别命名为CaTIFY1~CaTIFY15(图 1)。对CaTIFYs成员的理化性质分析显示(表1),氨基酸数目在153~411个之间,分子量在17 108.52~45 694.75 kD之间,等电点在4.92~10.07个之间,平均亲水性介于-0.147~-0.744之间,脂肪系数介于55.50~80.92之间,不稳定系数在33.34~62.70之间。亚细胞定位预测显示,CaTIFYs均定位于细胞核中。预测所有成员均无跨膜螺旋结构。蛋白二级结构显示,辣椒TIFY成员蛋白二级结构相似,各二级结构所占比例排序为:无规则卷曲>α-螺旋>延伸链>β-转角。无规则卷曲占比最高,为46.63%~73.37%,β-转角含量最少,为2.68%~7.14%。三级结构预测(图 2)显示,无规则卷曲及α-螺旋在三级结构中占主要部分,与预测的二级结构相同。

表1 CaTIFYs基因理化性质和二级结构分析Table 1 Physical and chemical properties and secondary structure analysis of CaTIFYs gene
2.2 辣椒TIFY基因家族保守基序、保守结构域和基因结构分析
对CaTIFYs的保守基序和保守结构域进行可视化分析(图 3),显示Motif 2存在于所有TIFY成员中,Motif 3和Motif 5仅存在于CaTIFY6、CaTIFY12、CaTIFY13中,CaTIFY1成员仅有Motif 2成员,Motif 6仅存在于CaTIFY15和CaTIFY8中。所有成员均含有TIFY保守结构域,属于TIFY superfamily,CaTIFY6、CaTIFY7、CaTIFY12、CaTIFY13含有CCT保守结构域,其他成员含有CCT-2保守结构域,分别属于CCT superfamily和CCT-2 superfamily。ZnF-GATA结构域仅存在于CaTIFY12和CaTIFY13,GATA仅存在于CaTIFY6中。

图2 辣椒TIFY蛋白的三级结构预测Fig.2 Prediction of the tertiary structure of pepper TIFY protein
对CaTIFYs基因结构进行分析(图 4),CaTIFY14仅含有2个外显子1个内含子,CaTIFY7和CaTIFY11含有3个外显子两个内含子,CaTIFY3和CaTIFY15含有5个外显子,CaTIFY1、CaTIFY9、CaTIFY10含有6个外显子,CaTIFY2、CaTIFY4、CaTIFY6、CaTIFY12含有7个外显子,CaTIFY5和CaTIFY8含有9个外显子,CaTIFY13含有10个外显子。
2.3 不同物种TIFY家族基因的系统发育分析
对拟南芥、水稻、番茄和辣椒74个TIFY蛋白序列进行多重比较,构建进化树(图 5),结果表明,TIFY蛋白分为Ⅰ~Ⅷ共八个大类群,第Ⅱ、Ⅲ、Ⅳ类群中不含CaTIFY成员,第Ⅱ类群中均为单子叶植物水稻中的TIFY成员。第Ⅰ类群中含有15个TIFY成员,其中CaTIFY15、CaTIFY3、CaTIFY11、CaTIFY1分别于SlTIFY9、SlTIFY15、SlTIFY13、SlTIFY12聚集在一起,处于一个分支中。第Ⅴ类群含有6个成员,其中CaTIFY8和CaTIFY5分别于SlTIFY3、SlTIFY10聚集在一个分支。第Ⅵ类群含有19个成员,其中CaTIFY10、CaTIFY13、CaTIFY7、CaTIFY12分别于SlTIFY16、SlTIFY8、SlTIFY18、SlTIFY4聚集在一个分支,而CaTIFY6与AtTIFY1聚为一个分支。第Ⅶ类群含有7个成员,CaTIFY1与SlTIFY20、SlTIFY5聚集为一分支。第Ⅷ类群含有11个TIYF成员,其中CaTIFY4、CaTIFY9、CaTIFY2分别与SlTIFY14、SlTIFY21、SlTIFY6聚集一个分支。

图3 辣椒TIFY基因家族的系统进化树(A)、保守基序(B)和保守结构域(C)分析Fig.3 Phylogenetic tree (A), conservative motif (B) and conservative domain (C) analysis of pepper TIFY gene family

图4 辣椒TIFYs基因结构分析Fig.4 Analysis of TIFYs gene structure in pepper

图5 不同物种TIFY家族基因系统发育树Fig.5 Phylogenetic tree of TIFY family genes in different species
2.4 辣椒TIFY基因家族成员顺式作用元件分析
使用TBtools软件进行可视化分析显示(图6),CaTIFYs除了含有大量的CAAT-box和TATA-box等基因元件外,还含有大量的调控作用元件。除CaTIFY8外,其余成员均含有较多的光响应元件(GT 1-motif、G-box、ACE、G-Box、Sp 1和3-AF 1 binding site),仅有CaTIFY5具有参与类黄酮生物合成基因调控的MYB结合位点(MBSI)。仅有CaTIFY5和CaTIFY6含有参与栅栏叶肉细胞分化的元件(HD-Zip 1)和参与胚乳表达的顺式调控元件(GCN4-motif)。CaTIFYs含有大量激素响应元件,如茉莉酸响应元件(CGTCA-motif和TGACG-motif)和水杨酸响应元件(TCA-element),所有CaTIFYs均含有赤霉素响应元件(ABRE),CaTIFY1、CaTIFY3、CaTIFY12、CaTIFY13含有生长素响应元件(TGA-element)。还有大量的响应逆境胁迫相关元件,除CaTIFY2、CaTIFY3、CaTIFY4、CaTIFY13外,其他成员均含有无氧诱导相关元件(ARE),CaTIFY1和CaTIFY8含有1个创伤响应元件(WUN-motif),还具有响应干旱胁迫(MBS)元件、低温响应元件(LTR)和参与防御和应激反应的顺式作用元件(TC-rich repeats)等。

注:L 1~L 9表示生长2~50 d的叶片;F 1~F 9表示花蕾出现后的2~50 d;P 10、O 10和STA 10分别为花蕾出现后60 d采集的花瓣;子房和雄蕊;FST 0与FST 1为第3天和第7天采集的整个果实;G 1~G 11为10~60 d的果皮;S 3~S 11为20~60 d的种子;ST 1和ST 2为10~15 d的胎座和种子混合;T 3~T 11分别代表花后20~60 d的胎座。图7 CaTIFY基因在果实(A)、叶片(B)和花(C)中不同发育时期的表达量变化Fig.7 Expression of CaTIFY gene in fruits(A), leaves(B) and flowers(C) at different development stages
2.5 辣椒TIFY基因家族成员组织表达特征分析
由图7可知,在叶片、花朵、果皮、种子和胎座发育过程中CaTIFY2表达量一直处于较高的表达水平,在叶片发育过程中CaTIFY3、CaTIFY11和CaTIFY14表达水平呈先升高后降低的趋势。在花朵发育过程中CaTIFY3、CaTIFY4、CaTIFY11、CaTIFY14在花朵发育20~40 d的表达量较高,CaTIFY4在雄蕊中表达量较高。在果实发育过程中,CaTIFY11和CaTIFY3在果皮发育的前期上调,在35~45 d表达量明显下调,然后逐渐上调。CaTIFY14表达量果皮发育前15 d较高,CaTIFY4和CaTIFY7在果皮发育的后期表达量逐渐上调。种子发育过程中,CaTIFY11的表达量较高,在种子发育55~60 d明显下调。CaTIFY12在种子发育的前15 d表达量较高,然后下调。在胎座发育过程中,CaTIFY11基因表达量较高,CaTIFY3、CaTIFY4和CaTIFY7基因表达水平随着胎座的发育逐渐上调,在发育60 d达到最大值,CaTIFY10和CaTIFY14在胎座发育的前35 d表达量较高,然后明显下调。

注:A、B、C、D分别为高温、低温、干旱和盐胁迫处理;E、F、G、H分别为赤霉素、脱落酸、茉莉酸甲酯和水杨酸处理;C为对照;L为叶片;R为根系;0~6分别表示处理0~24 h。图8 CaTIFYs基因在非生物胁迫和激素处理下的表达情况Fig.8 Expression of CaTIFYs genes in response to abiotic stress and hormone treatment
2.6 辣椒TIFY基因家族成员非生物胁迫和激素处理后表达特征分析
研究非生物胁迫和激素处理后CaTIFYs家族成员基因的表达模式(图 8)。总的来说,通过多种非生物胁迫和激素处理后,辣椒叶片和根系中CaTIFY2基因的相对表达量一直处于较高的表达水平,没有明显地下调,而CaTIFY6、CaTIFY8、CaTIFY15、CaTIFY1、CaTIFY13基因的表达水平一直较低,处理后没有明显地上调。叶片中CaTIFY9基因的表达量处理明显下调,而CaTIFY3的表达量上调,CaTIFY11的表达量呈先上调后下调的趋势。根系中CaTIFY4、CaTIFY9、CaTIFY11、CaTIFY14基因的表达量明显上调。
高温处理后,叶片中CaTIFY14处理前1 h上调然后下调,CaTIFY5前期下调,处理6~24 h逐渐上调。根系中CaTIFY10在处理0.5 h表达量上调,而CaTIFY5在处理6~24 h表达量明显上调。通过低温胁迫处理,叶片中CaTIFY14处理后0.5~24 h表达量逐渐上调;根系中CaTIFY3在处理1 h和24 h表达量明显上调;CaTIFY10基因的相对表达量在处理后上调。干旱处理结果表明,叶片中CaTIFY4和CaTIFY14在处理3 h表达量上调,CaTIFY3在处理1~6 h表达量上调;根系中CaTIFY10在处理0.5 h和6 h表达量明显上调,CaTIFY12在处理后表达量明显上调。使用NaCl进行盐胁迫处理,叶片中CaTIFY3和CaTIFY14在处理1~3 h表达量上调。根系中,CaTIFY14在处理前1 h表达量明显上调,然后逐渐下调。
赤霉素处理后,根系中CaTIFY10和CaTIFY12的表达量明显上调,CaTIFY3呈先上调后下调的趋势。通过脱落酸处理后,根系中CaTIFY3在处理1 h后明显上调,CaTIFY12在处理1.5 h~24 h表达量明显上调。茉莉酸甲酯处理后,叶片中CaTIFY3的表达量呈先上调后下调的趋势。水杨酸处理后,叶片中CaTIFY9的表达量明显下调,处理6 h明显上调,CaTIFY4、CaTIFY3、CaTIFY11、CaTIFY14基因的相对表达量在处理12~24 h明显上调,处理24 h达到最大值。根系中CaTIFY3的表达量在处理3 h和处理24 h明显上调。CaTIFY12和CaTIFY10处理前1 h表达量明显上调。
3 讨论与结论
TIFY基因家族是植物特有的转录因子,在植物生长发育和抗逆胁迫中发挥重要作用[30]。在模式植物中TIFY基因的鉴定和功能方面取得了重要进展,但是在辣椒中研究较少。本研究从辣椒中共鉴定到15个TIFY基因家族成员,与拟南芥中18个、大麦中15个[31]相似。15个TIFY成员分布于8条染色体上,不同CaTIFYs长度差异很大,意味着TIFY基因的复杂性较高。等电点位于4.92~10.07之间,CaTIFY1、CaTIFY6、CaTIFY7、CaTIFY12、CaTIFY13等电点均小于7,均翻译为酸性蛋白,其余成员均为碱性蛋白,与甘蓝(Brassicaoleraceavar. )TIFY研究结果相似,既有酸性蛋白又有碱性蛋白且碱性蛋白偏多[32]。辣椒TIFYs平均亲水性均小于0,且不稳定系数均小于40,说明CaTIFY均为不稳定亲水性蛋白,与毛果杨和西瓜(Citrulluslanatus)中TIFYs研究结果相似[12,33]。所有CaTIFYs亚细胞定位预测均位于细胞核中,前人研究黄瓜(CucumissativusL.)中TIFY均定位于细胞核[34],而在毛果杨[12]和核桃[10]等物种中TIFYs大部分定位于细胞核,也有部分TIFY定位于叶绿体、线粒体和内质网中。CaTIFYs蛋白保守基序数目较少,只有Motif 2存在于所有CaTIFY蛋白中,不同TIFYs基序呈现较高的多样性,推测在进化过程中存在较大分化。通过对TIFY基因结构分析显示外显子数目在3~10个之间,基因结构复杂,同一分组基因的结构相似,而不同分组中差异较大,与前人在水稻[8]和西瓜[33]中研究结果相似,根据其TIFY基因结构的多样性推测CaTIFY基因功能的多样性。
系统发育分析显示,CaTIFY家族蛋白分为八大类群,其中第Ⅱ类群中仅含有OsTIFYs成员,说明第Ⅱ类群起源于双子叶植物和单子叶植物分化之后。其中第Ⅱ、Ⅲ和Ⅳ类群中不含CaTIFYs成员,其他类群中,与单子叶植物水稻相比,辣椒与双子叶植物番茄和拟南芥的亲缘关系更近,多数CaTIFYs成员均与SlTIFYs聚集为一分支,进而推测辣椒与拟南芥和番茄这些双子叶植物具有相同的祖先。在甘蓝的研究中,将91个TIFY蛋白分为10个分支[32]。拟南芥中TIFY1不仅与花序的发育有关,而且还促进叶柄和下胚轴的延伸[1-2],其中CaTIFY6和AtTIFY1聚集同一分支,推测其与AtTIFY1具有相同的功能。
启动子顺式作用元件对调节基因表达至关重要,研究启动子中的顺式作用元件有助于基因转录和调控的研究[35]。CaTIFYs启动子中存在4种激素响应元件(生长素、茉莉酸甲酯、赤霉素和水杨酸),不同CaTIFYs中含有的元件类型不同且数量不等,表明CaTIFY基因对激素响应的程度不同,所以推测CaTIFYs间存在功能差异。多数CaTIFYs均含有非生物胁迫响应元件,在CaTIFYs中鉴定出多个非生物胁迫响应元件如厌氧诱导元件、防御和应激反应的顺式作用元件等,表明CaTIFYs在厌氧诱导、防御和胁迫反应中发挥重要作用。CaTIFY12含有两个低温响应元件,表明该基因可能在低温胁迫中起关键作用。值得注意的是,CaTIFY4、CaTIFY9、CaTIFY12、CaTIFY14并不含有响应干旱的诱导元件,但是转录组数据分析表明,通过干旱胁迫后这4个基因在根中强烈诱导,因此推测,它们可能与其他基因相互作用,来应对干旱胁迫。在研究中,CaTIFYs至少含有一个胁迫或激素响应元件,表明其对外源植物激素和环境胁迫有反应。
在植物生命周期中,不可避免地受到多种生物和非生物胁迫,对其生长发育造成影响。非生物胁迫如干旱、高温、低温和盐胁迫以及重要的植物激素等,在植物的生命周期中对其生长和发育造成一定伤害[36]。一些研究表明,TIFY基因家族在细胞信号转导和植物胁迫反应中具有重要的调节作用[37]。在本研究中,CaTIFY2、CaTIFY4、CaTIFY9、CaTIFY11、CaTIFY14在多种非生物胁迫以及激素处理后,表现出较高的表达水平,表明CaTIFYs在这些胁迫中具有重要功能。CaTIFY9在不同处理后,在叶片中表达量明显下调,而在根系中表达量明显上调,表明CaTIFY9在不同组织中具有不同的生理生化功能来适应各种环境胁迫。研究表明,JAs在植物生长发育中有作用,包括根系的生长发育以及叶片的衰老[38]。OsJAZ1可以通过水稻中的JA信号与OsMYC2相互作用来调节根系发育[39]。本研究中,通过MeJA处理后,根系中CaTIFY4、CaTIFY3、CaTIFY7、CaTIFY9、CaTIFY11、CaTIFY14基因的相对表达量显著上调,表明CaTIFYs成员可能通过JA信号通路参与辣椒根系生长和发育。
为研究CaTIFYs在辣椒生长发育中的作用,对其不同组织及发育时期的基因表达量进行测定,结果显示,在果实发育过程中,CaTIFY2、CaTIFY4、CaTIFY11在果皮、胎座和种子中表达量较高,而CaTIFY3、CaTIFY7、CaTIFY12仅在果皮和胎座中有一定的表达,在种子中表达量很低。CaTIFY2、CaTIFY3、CaTIFY11、CaTIFY14在叶片发育过程中表达量较高,CaTIFY2、CaTIFY3、CaTIFY4、CaTIFY11、CaTIFY14、CaTIFY15在花发育过程中表达量较高。表明CaTIFYs对辣椒生长过程、花芽分化及果实成熟过程中均发挥一定作用。通过芯片数据显示,玉米中TIFYs基因参与玉米不同发育阶段,具有组织特异性表达模式[40]。TIFY基因家族在植物生长发育和抗逆性中具有多重调控作用,该家族中含有大量与植物生命活动和抗逆反应相关的基因,因此对这些基因的挖掘及鉴定具有重要意义。
