三都野生茶树表型性状和生化组分多样性分析
2023-05-23杨春郭灿乔大河郭燕梁思慧李燕陈正武
杨春 郭灿 乔大河 郭燕 梁思慧 李燕 陈正武


摘要:为更好地保护和开发利用三都野生茶树资源,以16份三都野生茶树为研究对象,采用高效液相色谱和紫外分光光度法检测茶树的重要生化组分,并于春季和秋季对表型性状开展观测,结合变异系数和遗传多样性指数分析三都野生茶树资源变异程度和多样性水平。结果表明,20个描述性表型性状变异系数在0~35.68%之间,平均值为9.94%,遗传多样性指数在0~1.095之间,平均值为0.257;9个数量性表型性状变异系数在0~32.60%之间,平均值为12.68%,遗传多样性指数在0~1.927之间,平均值为1.077。17个生化性状变异系数和遗传多样性指数变幅分别为0~400.0%和0~1.927,平均值分别为65.28% 和1.320。三都野生茶树资源表型鉴定性状表现为子房3室、无毛、顶芽有毛。生化特征表现为高茶多酚、高C、高可可碱和低游离氨基酸、无茶氨酸、低儿茶素总量、低EGCG、低或无咖啡碱。三都野生茶树资源表型性状和生化组分较统一,资源内部变异小,C和可可碱为优势特征组分。结合表型和生化特征,推测三都野生茶树资源可能为榕江茶的变种。
关键词:三都;野生茶树;遗传多样性指数;可可碱;表型性状;生化组分
中图分类号:S571.101 文献标志码:A
文章编号:1002-1302(2023)08-0111-08
基金项目 贵州省科技计划(编号:黔科合支撑[2019]2254号、黔科合平台人才[2020]2103号);贵州省农业科学院特色动植物资源收集、保存与创制利用项目(编号:黔农科院种质资源[2022]03号)。
作者简介:杨 春(1988—),女,湖南益阳人,硕士,助理研究员,主要从事茶树资源育种与品种抗性研究。E-mail:610681998@qq.com。
通信作者:陈正武,研究员,主要从事茶树资源育种工作。E-mail:zwchentea@163.com。
我国是茶树[Camellia sinensis (L.) O.Kuntze]的原产地,云南、贵州等省分布着数量繁多、类型多样的野生茶树资源[1-4],这些野生茶树资源对于茶树的遗传演化、品种改良及分子育种等研究有着重要意义[5-7]。因此,加大力度保护和深入研究珍稀野生茶树资源,是现今茶树种质资源研究的重点[8]。
三都水族自治县,位于贵州省黔南布依族苗族自治州,地处月亮山、雷公山腹地,是贵州野生茶树集中分布地之一[9]。前期已针对三都野生茶树亲缘关系、遗传多样性等展开了一定研究[10-11],但尚未见三都野生茶树表型特征及生化性状等方面的相关报道。本研究选取16份代表性三都野生茶树为研究对象,通过测定主要生化成分和观测树、花、叶等表型性状,结合统计分析方法,明晰三都野生茶树资源表型及生化的变异程度和多样性水平,归纳出三都野生茶树资源整体的表型特征和生化特征,以期为进一步保护和开发利用三都野生茶树资源打下基础。
1 材料与方法
1.1 仪器与试剂
高效液相色谱(HPLC)仪:Agilent 1260 Infinity Ⅱ(美国安捷伦);
紫外分光光度计:TU-1800(北京普析);
色谱柱:大连依利特Hypersil ODS2(4.6 mm×250 mm,5 μm)。
咖啡碱、没食子酸、儿茶素类[儿茶素(C)、表儿茶素(EC)、表没食子儿茶素(EGC)、表儿茶素没食子酸酯(ECG)、没食子儿茶素没食子酸酯(GCG)、表没食子儿茶素没食子酸酯(EGCG) ]和茶氨酸标准品购自合肥博美生物科技有限公司,N,N-二甲基甲酰胺、乙腈、甲醇、冰醋酸等有机试剂为市售HPLC级。
1.2 试验材料
试验茶树取自贵州省黔南州三都水族自治县原始森林中,地理范围为25°55′48″~25°59′40″N,108°9′32″~108°20′46″E,海拔范围为1 300~1 500 m。选取基径大于10 cm的野生茶树进行取样和挂牌(图1)。福鼎大白茶(对照)取自贵州省茶叶研究所湄潭种质资源圃。
1.3 试验方法
1.3.1 样品制备 于2020年5月,采各标记单株1芽2叶鲜叶,用微波杀青的方法[12],制备生化检测样。对照取样时间为2020年4月,取样标准1芽2叶。
1.3.2 检测方法
表型性状:分别于2020年、2021年春季和秋季调查记录树、叶、花、芽的性状,表型调查及描述代码参考《茶树种质资源数据质量控制规范》,采用基恩士超景深显微镜(VHX-5000)观测顶芽、子房和腋芽茸毛,每份材料调查10个叶、芽、花,数量性表型性状取平均值,描述性表型性状以出现概率最大的描述代码为准。
生化成分:游离氨基酸含量采用GB/T 8314—2013 《茶 游离氨基酸总量的测定》中的方法进行测定;茶多酚含量采用GB/T 8313—2018 《茶叶中茶多酚和儿茶素类的检测方法》进行测定;茶氨酸含量参考岳婕等的方法[13]进行测定;儿茶素组分参考周顺珍等的方法[14]进行测定。每个样品重复检测3次,数据取3次平均值。
1.3.3 数据处理
评价遗传多样性指数采用香农-维纳指数(Shannon-Wiener index)。性状分布频率及多样性分析采用Excel 2010软件。聚类分析和Pearson相关性分析采用SPSS 21.0。
2 结果与分析
2.1 表型性状遗传多样性分析
本研究共统计分析了16份三都野生茶树树型、叶片大小、叶形等20个描述性表型性状和叶长、叶宽等9个数量性表型性状(表1、表2)。20个描述性表型性状在描述代码上有不同的分布表现,其中叶片大小和雌雄蕊相对高度在描述代码上均有分布但不均匀,具体叶片大小以中叶居多,雌雄蕊相对高度以等高居多。树型、叶形、叶身、叶齿密度、叶齿锐度、叶尖及叶色不均匀分布于描述代码上的2~3项,具体来看,树型表现为小乔木和乔木,以小乔木为主;叶形主要为椭圆形、长椭圆形和披针形,三者数量相差不大;叶身表现为内折和平,以内折居多;葉齿以密度密、锐度中居多;叶尖表现为急尖和渐尖,渐尖居多;叶色有绿色和深绿色,深绿色占绝大多数。叶面、叶质、叶基、萼片茸毛、萼片颜色、花瓣颜色、花瓣质地、花柱裂位、子房茸毛、腋芽茸毛及顶芽茸毛全部集中分布于1个描述代码,具体表现为叶面微隆起、叶质中、叶基楔形、萼片茸毛无、萼片颜色绿色、花瓣颜色白色、花瓣质地薄、花柱裂位高、子房无茸毛、腋芽和顶芽有茸毛。
9个数量性表型性状中萼片数和柱头开裂数并无变化,稳定表现出萼片5瓣和柱头3裂的性状;叶脉对数和花瓣数同样变幅较小,叶脉对数表现出8对或9对,花瓣7瓣居多;叶长、叶宽、叶面积、花冠直径及花柱长度的变幅相对较大,遗传多样性指数同样高于其他4个性状。
20个描述性表型性状变异系数在0~35.68%之间,平均值为9.94%;遗传多样性指数在0~1.095之间,平均值为0.257。9个数量性表型性状变异系数在0~32.60%之间,平均值为12.68%;遗传多样性指数在0~1.927之间,平均值为1.077。29个表型性状的平均变异系数和遗传多样性指数分别为10.79%和0.512,表明三都野生资源表型性状的变异系数和遗传多样性指数处于较低水平,且有11个描述性表型性状和2个数量性表型性状集中分布,变异系数和遗传多样性指数都为0,这说明三都野生茶树在表型上有多个性状表现一致,资源内部变异较小,存在一定的封闭性。
2.2 生化性状遗传多样性分析
参照茶树中常规生化成分和特征性次生代谢产物的标准对16份三都野生茶树的17个生化性状进行分析,以福鼎大白茶为对照比对三都资源与代表性栽培种在生化性状上的典型差异。
由表3可知,17个生化性状变异系数和遗传多样性指数变幅分别为0~400.0%和0~1.927,平均值分别为65.28% 和1.320。三都资源在生化成分上与对照福鼎大白茶差异较大。福鼎大白茶中游离氨基酸和茶氨酸含量分别是3.94%和2.02%,而三都资源中游离氨基酸含量较低,最高仅1.63%,低于对照福鼎大白茶的一半,且16份三都野生茶树中均未检测到茶氨酸。福鼎大白茶中茶多酚含量为16.78%,儿茶素总量为12.49%,儿茶素占比为74.43%,符合儿茶素占茶多酚含量70.0%~80.0%的规律[15],而三都资源表现出典型的高茶多酚、 低儿茶素的特征,茶多酚含量、儿茶素含量和儿茶素占比平均值分别为22.99%、6.29%和27.46%。对比三都野生茶树和福鼎大白茶的儿茶素单体可知,福鼎大白茶中主要儿茶素单体为表没食子儿茶素没食子酸酯(EGCG),含量为7.02%,占儿茶素总量的56.2%。三都资源中占主导的儿茶素单体为儿茶素(C),含量在3.59%~5.56%之间,其次是表儿茶素没食子酸酯(ECG),含量在0.71%~1.69%之间,表没食子儿茶素(EGC)、没食子儿茶素没食子酸酯(GCG)、EGCG和表儿茶素(EC)含量均较低。对比生物碱组成可知,福鼎大白茶中主要生物碱是咖啡碱,含量为3.79%;三都资源中的主要生物碱为可可碱,含量范围在3.31%~5.76%之间,几乎不含咖啡碱。
2.3 聚类分析和相关性分析
2.3.1 聚类分析
以资源间存在变异的22个数量指标(叶长、叶宽、叶面积、叶脉对数、花瓣数、花冠直径、花柱长度、氨基酸含量、茶多酚含量、没食子酸含量、EGC含量、C含量、EC含量、EGCG含量、ECG含量、非酯型儿茶素含量、酯型儿茶素含量、儿茶素总量、儿茶素占比、咖啡碱含量、可可碱含量和茶碱含量)为依据,对16份三都野生茶树进行聚类分析。
由图2、表4可知,16份三都野生茶树在欧式距离为10时,可分为3类。第Ⅰ类包括SD-26、SD-29 等7份野生茶树,主要特点为叶长、叶宽和叶面积均较小,茶多酚和酯型儿茶素含量较高,氨基酸含量较低;第Ⅱ类包括SD-2、SD-20-1等8份野生茶树,主要特点为氨基酸含量较高,C含量、非酯型儿茶素含量和儿茶素总量较低,儿茶素占比较低,可可碱含量较高;SD-20-2单独构成第Ⅲ类,主要特点为叶长、叶宽和叶面积较大,茶多酚、酯型儿茶素和可可碱含量较低,C、非酯型儿茶素含量和儿茶素总量较高,儿茶素占比较高。
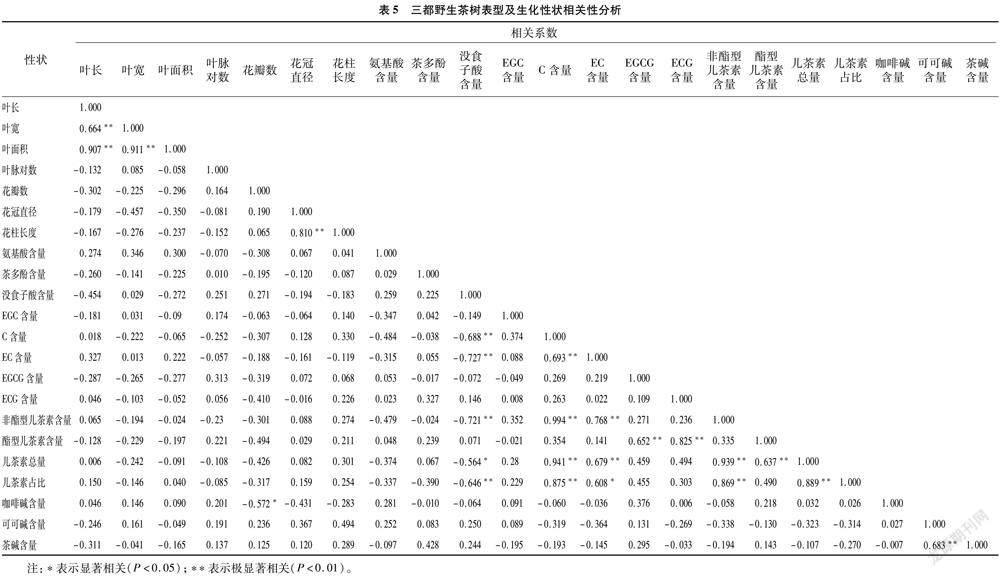
2.3.2 相关性分析
同样以资源间存在变异的22个数量指标进行Pearson相关性分析,结果(表5)显示,三都野生茶树表型性状与生化性状间相关性不大,仅咖啡碱与花瓣数存在显著负相关。表型性状中叶片性状相关性较强,叶长、叶宽和叶面积三者间存在极显著正相关,花性状仅中花冠直径与花柱长度间存在极显著正相关,叶片性状与花性状间不存在显著相关性。生化性状间的相关性要强于表型性状,有19对生化性状间存在显著或极显著相关性,没食子酸含量与C、EC、 非酯型儿茶素含量及儿茶素总量和儿茶素占比间存在显著或极显著负相关。C含量和EC含量不仅相互间存在极显著正相关,同时与非酯型儿茶素含量、儿茶素总量和儿茶素占比间存在显著或极显著正相关关系。EGCG含量和ECG含量与酯型儿茶素含量存在极显著正相关。3个生物碱成分中,可可碱含量与茶碱含量间存在极显著正相关。
3 讨论
3.1 三都野生茶树资源多样性分析
三都野生茶树表型性状和生化成分的遗传多样性指数平均值分别为0.512和1.320,低于陕西[16]、江西[17]、重庆[18]及贵州多地的地方茶树种质[19-20],也低于纳雍[21]、镇宁[22]、雷波[23]等地的野生茶树。说明三都野生茶树生长环境较封闭,表型及生化遗传背景单一,资源内部变异类型不丰富。
3.2 三都野生茶树资源初鉴
子房有无茸毛、子房室数和顶芽(腋芽)是否具毛是茶种植物分类鉴定的主要依据[24]。茶种植物(C. sinensis)的鉴定表型特征为子房3室、有毛、顶芽有毛[25],而三都野生茶树则是子房3室、无毛、顶芽(腋芽)有毛的表型特征。参考陈涛林关于榕江茶、突肋茶和元宝山茶的研究[26]发现,三都野生茶树在形态上与榕江茶相似度较高,除同样是子房3室、无毛,顶芽有毛外,在叶长、叶宽、叶面积、叶尖、叶形、花柱長度等性状上同样表现相近,但在树型、萼片茸毛等性状上存在明显差异,榕江茶为灌木,萼片外侧有疏毛,三都野生茶树不存在灌木类群,且萼片外侧光滑无毛。生化上,三都野生茶树与榕江茶均表现出以可可碱为主要生物碱、C为主要儿茶素单体及茶氨酸含量甚微的生化特征。因此,结合表型性状和生化特性,推测三都野生茶树与元宝山野生茶树一样,可能为榕江茶变种。
3.3 三都野生茶树资源生化特性与利用
咖啡碱、EGCG和茶氨酸是茶树中的优势组分[15]。大量研究发现,野生茶树有着区别于常规栽培种茶树的生化特性。滕杰等研究发现,秃房茶(C. gymnogyna)中生物碱组成明显有别于其他茶种,以可可碱为主要生物碱[27]。Jin等研究发现,福建野生红芽茶中主要儿茶素单体为GCG,主要生物碱组分是可可碱[28]。陈潇敏等研究发现,福建野生茶树中存在以苦茶碱为主要生物碱的特异种质[29]。三都野生茶树资源中C和可可碱为优势组分,可可碱对癌症、心血管疾病、呼吸疾病等方面有积极干预与治疗作用[30],后续可作为特异茶树种质资源加以开发利用。
4 结论
三都野生茶树资源表型和生化的遗传多样性指数均处于较低水平,在表型和生化层面分化程度不高。主要鉴定表型性状表现为子房3室、无毛、顶芽(腋芽)有毛。生化性状整体表现出高茶多酚、低儿茶素、低氨基酸和无茶氨酸的特点,C和可可碱是三都野生茶树的特征性优势组分。结合表型性状和生化特性,推测三都野生茶树可能为榕江茶变种。本研究仅对三都野生茶树资源表型及茶树常规生化开展研究,后续可结合非靶向代谢组分析三都野生茶树与栽培种的差异代谢物,同时结合全基因组测序等分子手段,分析三都野生茶树在茶树起源及进化中的地位。
参考文献:
[1]张文驹,戎 俊,韦朝领,等. 栽培茶树的驯化起源与传播[J]. 生物多样性,2018,26(4):357-372.
[2]刘亚兵,赵华富,乔大河,等. 野生大树茶种质资源的品质指标多样性分析评鉴[J]. 食品工业,2020,41(9):168-172.
[3]牛素贞,安红卫,宋勤飞,等. 贵州野生茶树立地土壤养分状况分析及综合评价[J]. 浙江农业学报,2020,32(6):1039-1048.
[4]王 菲,程小毛,肖云龙,等. 千家寨野生古茶树叶片解剖结构和化学组分计量特征对海拔梯度的适应[J]. 生态学杂志,2021,40(7):1958-1968.
[5]Meegahakumbura M K,Wambulwa M C,Li M M,et al. Domestication origin and breeding history of the tea plant (Camellia sinensis) in China and India based on nuclear microsatellites and cpDNA sequence data[J]. Frontiers in Plant Science,2018,8:2270.
[6]Jiang C K,Zhao W F,Zeng Z,et al. A treasure reservoir of genetic resource of tea plant (Camellia sinensis) in Dayao Mountain[J]. Genetic Resources and Crop Evolution,2018,65(1):217-227.
[7]Du J K,He X L,Zhou Y M,et al. Gene coexpression network reveals insights into the origin and evolution of a theanine-associated regulatory module in non-Camellia and Camellia species[J]. Journal of Agricultural and Food Chemistry,2021,69(1):615-626.
[8]金基强,张晨禹,马建强,等. 茶树种质资源研究“十三五”进展及“十四五”发展方向[J]. 中国茶叶,2021,43(9):42-49,76.
[9]陈湖芳. 基于基因组重测序揭示贵州古茶树起源与进化的研究[D]. 贵阳:贵州大学,2021:63-64.
[10]刘青,赵德刚,赵懿琛. 古茶树种质资源遗传多样性ISSR分析[J]. 种子,2021,40(5):7-14.
[11]郭 灿,皮发娟,吴昌敏,等. 基于GBS测序的全基因组SNP揭示贵州地方茶组植物资源的亲缘关系[J]. 南方农业学报,2021,52(3):660-670.
[12]靖翠翠,杨秀芳,谭 蓉,等. 微波制样对茶叶内质成分的影响[J]. 食品安全质量检测学报,2015,6(4):1265-1270.
[13]岳 婕,李 丹,杨 春,等. 不同茶树品种氨基酸组分及含量分析[J]. 湖南农业科学,2010(23):141-143.
[14]周顺珍,龚 雪,周国兰,等. HPLC法测定茶叶中儿茶素及咖啡碱[J]. 化学分析计量,2013,22(5):27-29.
[15]宛晓春,夏 涛. 茶树次生代谢[M]. 北京:科学出版社,2015:1-3.
[16]丁帅涛,程晓梅,张 亚,等. 基于表型性状和生化成分的陕西茶树种质资源遗传多样性研究[J]. 西北农业学报,2019,28(4):607-619.
[17]王治会,彭 华,江新凤,等. 江西茶树种质资源芽叶表型多样性分析[J]. 江苏农业科学,2020,48(1):134-138,142.
[18]翟秀明,李 解,唐 敏,等. 重庆30份茶树种质资源农艺性状与生化成分多样性[J]. 浙江农业学报,2021,33(7):1244-1255.
[19]张小琴,周富裕,杨 春,等. 贵定鸟王种茶树资源农艺性状和品质性状多样性分析[J]. 分子植物育种,2015,13(2):415-423.
[20]杨 春,吴昌敏,石伟昌,等. 黎平地方茶树资源生化成分多样性分析及优异单株鉴选[J]. 种子,2020,39(10):63-66,72.
[21]葛立雯,郭 维,潘正康,等. 贵州姑菁野生茶树形态多样性及相关性研究[J]. 植物遗传资源學报,2015,16(3):497-502.
[22]曹 雨,乔大河,赵华富,等. 25份贵州镇宁野生茶树种质资源的表型及生化组分多样性分析[J]. 中国农学通报,2018,34(14):81-88.
[23]周 斌,王留彬,韩浩蕾,等. 雷波野生茶树资源春季新梢主要生化成分分析[J]. 西北农业学报,2020,29(3):390-397.
[24]陈 亮,虞富莲,童启庆. 关于茶组植物分类与演化的讨论[J]. 茶叶科学,2000,20(2):89-94.
[25]闵天禄. 世界山茶属的研究[M]. 昆明:云南科技出版社,2000:130.
[26]陈涛林. 广西元宝山一种特异茶饮植物的系统学鉴定与综合评价研究[D]. 长沙:湖南农业大学,2019:16-72.
[27]滕 杰,曾 贞,黄亚辉. 秃房茶嘌呤生物碱组成特点及生化品质成分的研究[J]. 广西植物,2018,38(5):568-576.
[28]Jin J Q,Chai Y F,Liu Y F,et al. Hongyacha,a naturally caffeine-free tea plant from Fujian,China[J]. Journal of Agricultural and Food Chemistry,2018,66(43):11311-11319.
[29]陈潇敏,赵 峰,王淑燕,等. 福建野生茶树资源嘌呤生物碱构成评价及特异资源筛选[J]. 茶叶科学,2022,42(1):18-28.
[30]吴文亮,童 彤,胡 瑶,等. 可可茶及其优势化学成分的健康功效研究进展[J]. 茶叶科学,2021,41(5):593-607.
