GRB7 基因在5 个猪群体内的多态性分析
2023-05-18吴华莉张莺莺王洪洋曹建国涂尾龙谈永松
吴华莉,张莺莺,黄 济,王洪洋,曹建国,涂尾龙,范 春,谈永松*
(1 上海市农业科学院畜牧兽医研究所,上海 201106;2 上海种猪工程技术研究中心,上海 201302;3 上海实验动物研究中心,上海 201302)
生长因子受体结合蛋白7(Growth factor receptor-bound protein7,GRB7)是重要的信号衔接蛋白,介导来自FAK 和EphB1 受体的信号通路,在细胞肿瘤迁移、增殖等过程中发挥重要作用[1-3]。 GRB7 蛋白含535 个氨基酸残基,结构上有3 个保守区域,分别为富含脯氨酸的N 末端、GM 结构域和C 末端SH2 结构域。 GRB7 与GRB10、GRB14 序列同源性较高,共同构成GRB7∕10∕14 家族蛋白,其主要是通过C 端的SH2结构域识别胞膜酪氨酸激酶受体,参与级联酶促反应[4]。 人的GRB7 基因位于17q21[5]。 猪GRB7 基因定位于12 号染色体,包含15 个外显子,长度为2 191 个核苷酸[6]。 杨金娥[7]研究发现猪GRB7 存在两个突变位点(理论上可产生9 个合并基因型),突变位点对应基因片段的163 bp 和243 bp 处。 杨述林等[8]研究发现,猪GRB7 基因163 位点和243 位点与背膘厚显著相关(P<0.05)。 本试验通过检测GRB7 基因163 突变位点和243 突变位点在上海地区5 个猪群体中的多态分布,旨在为猪遗传育种研究提供参考。
1 材料与方法
1.1 材料采集
于上海祥欣畜禽有限公司采集92 个杜洛克猪、148 个大白猪和69 个长白猪猪耳组织块,于上海猪状元畜牧综合场采集79 个梅山猪猪耳组织块,于上海富民农场采集74 个申农猪猪耳组织块,共计462 个样品。 将其放入预先加入1.0 mL 75%酒精的1.5 mL 离心管内,储存在冰盒里运回实验室,置于-20 ℃冰箱保存备用。
组织DNA 提取试剂盒采购于上海轩略科技发展有限公司,PCR 引物在生工生物工程(上海)股份有限公司合成,2 ×Taq PCR Master Mix 和DL 2000 DNA Marker 采购于宝生物工程(大连)有限公司;BstUⅠ内切酶采购于上海朝瑞生物科技有限公司。
1.2 试验方法
1.2.1 DNA 提取
利用AXYGEN 组织DNA 提取试剂盒提取基因组DNA,并将其统一稀释至50.0 ng∕mL,分装后将样品保存在-80 ℃冰箱内备用。
1.2.2 PCR 扩增
PCR 反应体系为20 μL:DNA 模板1.0 μL,2 ×Taq PCR Master Mix 11.0 μL,上游引物0.5 μL,下游引物0.5 μL,RNase-Free ddH2O 补至20 μL。 扩增上游引物序列为5’-GTCTCATCTCAGCAGCTCC-3’,下游引物序列为5’-GAGCATCTCGCACACGTAG-3’[7]。 反应条件为:94 ℃5 min;94 ℃30 s,63 ℃30 s,72 ℃30 s,32 个循环;72 ℃10 min。 利用1.2%琼脂糖凝胶电泳检测PCR 产物。
1.2.3 基因型检测
选择只有一条目的条带的样品PCR 产物,进行限制性内切酶试验。 反应体系为:PCR 产物4.0 μL,10 ×Buffer 2.0 μL,BstUⅠ内切酶0.5 μL,ddH2O 9.5 μL,总体积为16.0 μL。 反应温度37 ℃,时间2 h,利用1.8%琼脂糖凝胶电泳检测基因型。
1.3 基因型统计
PCR 扩增序列片段存在两处T∕C 多态,分别位于GRB7 基因第2 内含子和第3 外显子[7]。 理论上PCR 扩增产物经过BstUⅠ酶切后可产生9 种基因型[7](表1)。

表1 GRB7 基因酶切后的基因型Table 1 Genotypes of GRB7 gene after enzyme digestion
1.4 数据统计及分析
根据参考文献[9]中方法计算GRB7 基因163 位点和243 位点突变在5 个猪群体内的基因型频率、等位基因频率以及合并基因型频率。 采用GENEPOP 3.1 软件计算基因杂合度(Heterozygosity,He)、纯合度(Homozygosity,Ho)、有效等位基因数(Effective number of alleles,Ne)和多态信息含量(Polymorphism information content,PIC)。
2 结果与分析
2.1 猪GRB7 基因PCR 扩增
由图1 可知,以各样本DNA 为模板,PCR 产物经电泳检测后,获得的目的条带均为626 bp。 该片段包含2 个突变位点,可用于后续的酶切试验。
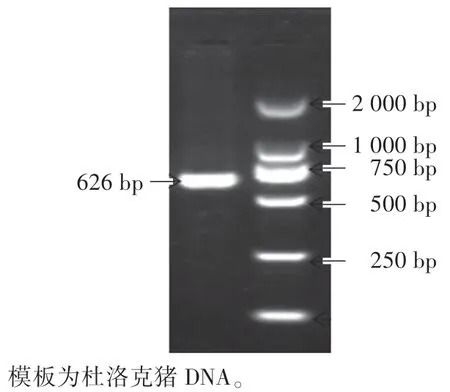
图1 GRB7 基因PCR 产物的琼脂糖凝胶电泳图Fig.1 PCR products of agarose gel electrophoresis of GRB7 gene
2.2 PCR-RFLP 琼脂糖凝胶电泳检测
猪GRB7 基因163 位点和243 位点经过内切酶BstUⅠ消化后,检测出6 种合并基因型(图2,图3):(1)CCTT 型:463 bp、163 bp; (2) CCTC 型: 463 bp、 383 bp、 163 bp、83 bp;(3)CCCC 型:383 bp、163 bp、83 bp;(4)CTTT 型:626 bp、463 bp、163 bp;(5)CTCT 型:626 bp、383 bp、163 bp、83 bp;(6)TTTT 型:626 bp。
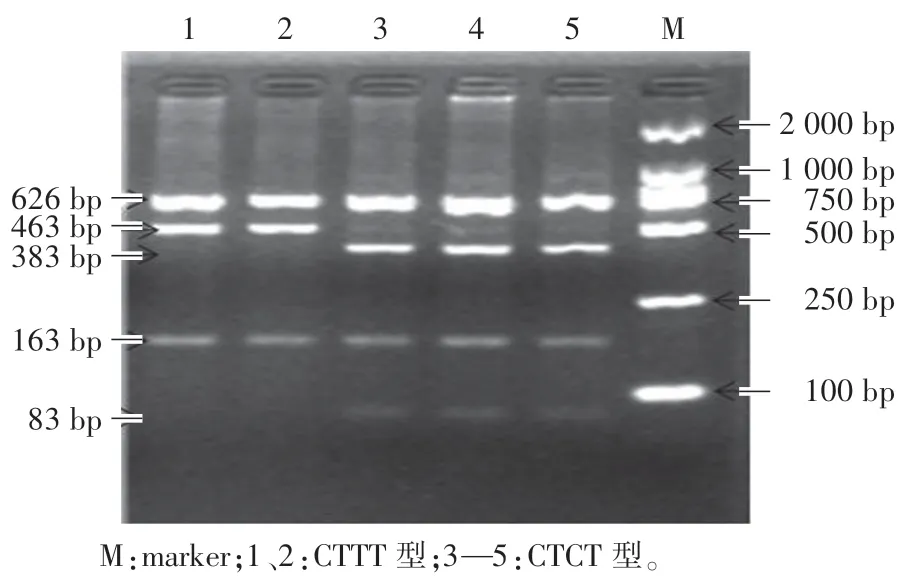
图2 猪GRB7 基因PCR-RFLP 琼脂糖凝胶电泳图Fig.2 PCR-RFLP result of agarose gel electrophoresis of GRB7 gene
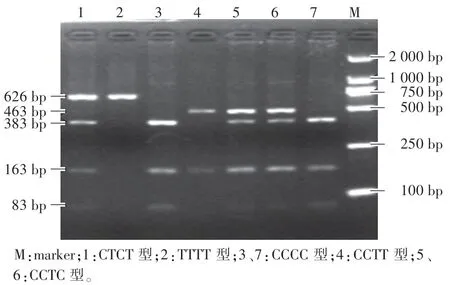
图3 猪GRB7 基因PCR-RFLP 琼脂糖凝胶电泳图Fig.3 PCR-RFLP result of agarose gel electrophoresis of GRB7 gene
2.3 GRB7 基因在5 个猪群体内的基因型频率、等位基因频率及群体遗传结构
由表2 所示,163 位点在杜洛克猪群体内CC 基因型为优势基因型,只有1 个样品为TT 基因型,无CT基因型;在长白猪和大白猪群体内CC 基因型和CT 基因型呈中态分布,只有少量TT 基因型;在申农猪群体内CC 基因型少于CT 基因型和TT 基因型;在梅山猪群体内TT 基因型最多,CT 基因型较少,未检测出CC 基因型。 在等位基因频率方面,163 位点在梅山猪群体内T 等位基因比例较高,在杜洛克猪、长白猪和大白猪群体内C 等位基因比例高,在申农猪群体内C 和T 等位基因呈现均匀分布。 163 位点突变PIC 值在长白猪、大白猪和申农猪群体内为0.25—0.50,表现为中度多态,在杜洛克和梅山猪群体内的PIC 值低于0.25,表现为低度多态。
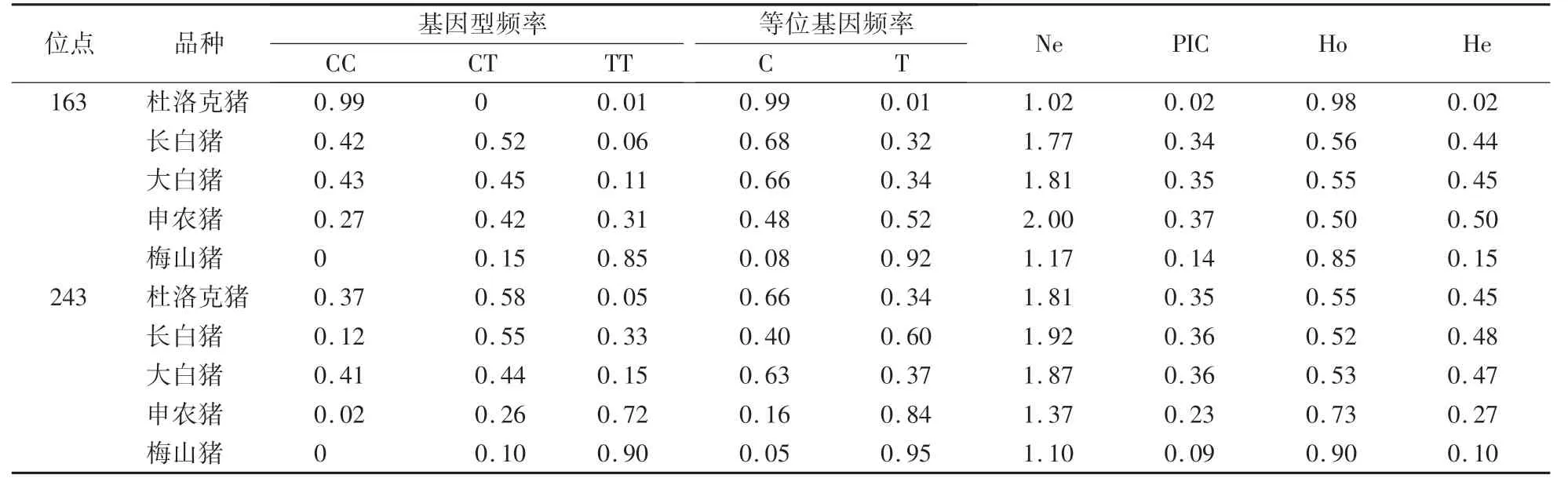
表2 GRB7 基因在5 个猪群体内的基因型频率、等位基因频率及群体遗传结构Table 2 Genotype and gene frequency and population genetic structure indices of GRB7 gene in five pig populations
243 位点在杜洛克猪群体内CT 基因型最多,其次为CC 基因型,只有少量TT 基因型;在长白猪群体内CT 基因型最多,TT 基因型略少,CC 基因型最少;在大白猪群体内CC 基因型和CT 基因型呈中态分布,TT 基因型最少;在申农猪群体内TT 基因型最多,CT 基因型略少,CC 基因型最少;在梅山猪群体内TT 基因型最多,CT 基因型很少,无CC 基因型。 243 位点在梅山猪和申农猪群体内T 等位基因比例较高,在长白猪群体内T 等位基因为优势等位基因,在杜洛克猪和大白猪群体内C 等位基因为优势等位基因。 243位点突变PIC 值在杜洛克猪、长白猪和大白猪群体内为0.25—0.50,表现为中度多态,在申农猪和梅山猪群体内的PIC 值低于0.25,表现为低度多态。
2.4 GRB7 基因163 位点和243 位点突变合并基因型频率的分布
如表3 所示,在杜洛克猪群体内GRB7 基因163 位点和243 位点突变合并基因型有4 种,其中CCTC型为优势基因型,CCTT 型和CTCT 型频率较低;在长白猪群体内有6 种,其中CCTC 型为优势基因型,CCTT 型和TTTT 型频率较低;在大白猪群体内有6 种,其中CTCT 型为优势基因型,CCTT 型和CCTC 型频率最低;在申农猪群体内有6 种,其中TTTT 型为优势基因型,CCTC 型和CCCC 型频率较低;在梅山猪群体内有3 种,其中TTTT 型为优势基因型,CTTT 型和CTCT 型频率较低。
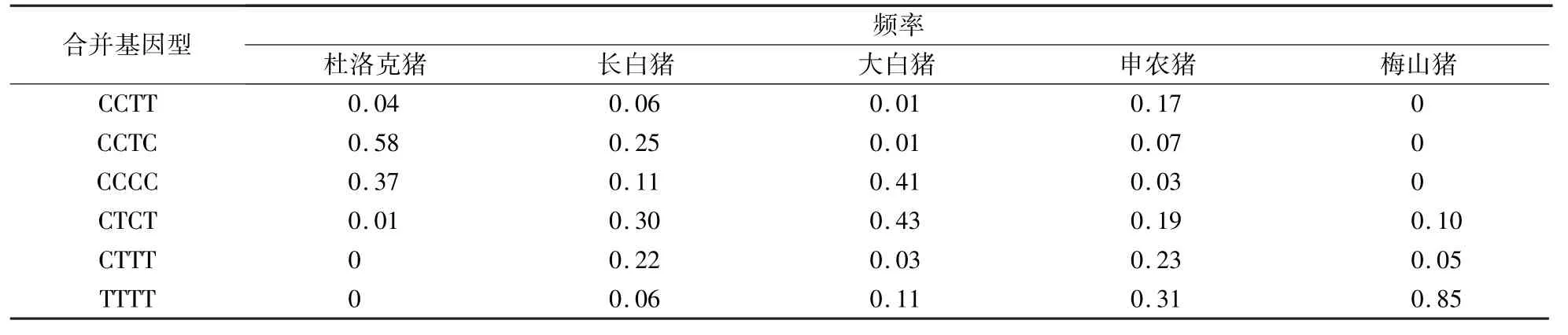
表3 GRB7 基因163 位点和243 位点突变合并基因型频率Table 3 The frequency distribution of GRB7 combined genotype
3 讨论
GRB7 基因作为信号联结蛋白,参与细胞增殖和肿瘤转移过程,在乳腺癌、食管癌和胃肠道间质瘤组织中高度表达,可以作为癌症预后标志物[10-13]。 有研究表明,小鼠和绵羊GRB7 基因在动物早期胚胎附植过程中发挥作用[14-15]。 杨金娥[7]研究发现,猪GRB7 基因163 位点和243 位点突变的CC 基因型个体的红细胞数目比TC 基因型个体的红细胞数目多。
合并基因型的效应不是单个基因型效应的简单相加,群体的遗传改良还受合并基因型频率影响[16]。本试验发现,GRB7 基因163 位点和243 位点突变所产生的优势基因型、等位基因频率和合并基因型及频率在5 个不同猪群体内不一样。GRB7 基因TTTT 合并基因型在申农猪和梅山猪群体内为优势基因型,在杜洛克猪和长白猪群体内CCTC 合并基因型频率较高,在大白猪群体内CTCT 合并基因型频率较高。 申农猪是长白猪×(大白猪×二花脸猪)为基础群构建的母本新品系,含有中国地方猪种二花脸猪血统,二花脸猪和梅山猪都属于太湖猪,是仔数高产品种,但申农猪和梅山猪的背膘厚比外来品种杜洛克、长白猪和大白猪高[17-22],可能与猪GRB7 基因163 位点和243 位点突变有关[8]。 下一步将检测TTTT 合并基因型与申农猪和梅山猪的背膘厚的关系。
