桃蚜取食抗、感辣椒品种的茉莉酸途径基因表达差异分析
2023-05-16税军刘小强陈青梁晓伍春玲刘迎姚晓文乔阳毛立杰陈银华
税军 刘小强 陈青 梁 晓 伍春玲 刘迎 姚晓文 乔阳 毛立杰 陈银华


关键词:桃蚜;抗、感辣椒品种;密度;取食时间;茉莉酸;合成途径;信号途径;基因表达
中图分类号:S436.418.2 文献标识码:A
辣椒(Capsicum annuum L.)近年来逐步成为我国产值最高的蔬菜作物,年总产值超700亿元。我国是世界上种植辣椒面积最大的国家,2020 年辣椒播种面积约占世界播种总面积的40%[1]。2020 年海南辣椒生产占其瓜菜播种总面积的13.1%[2],辣椒产业是热带特色高效农业、乡村振兴及南繁种业发展中的重要组成部分。
桃蚜(Myzus persicae Sulzer)是世界上分布最广的蚜虫之一,以口针吸食寄主植物幼嫩组织汁液,并排泄蜜露,诱发煤污病,而且可传播115 种植物病毒[3]。桃蚜在海南辣椒上,一年可发生20 代以上[4],世代重叠且快速繁殖,严重威胁茄科辣椒属作物的生产安全。海南省作为南方冬季辣椒北运主产区之一,伴随着该地区辣椒产业的快速发展,桃蚜的为害愈发严重。当前登记防蚜药剂种类较少,大剂量高频率不合理的药剂防治所导致的农产品安全、产地环境安全和抗药性等问题十分突出。因此,亟需寻找新的桃蚜防控策略和途径。
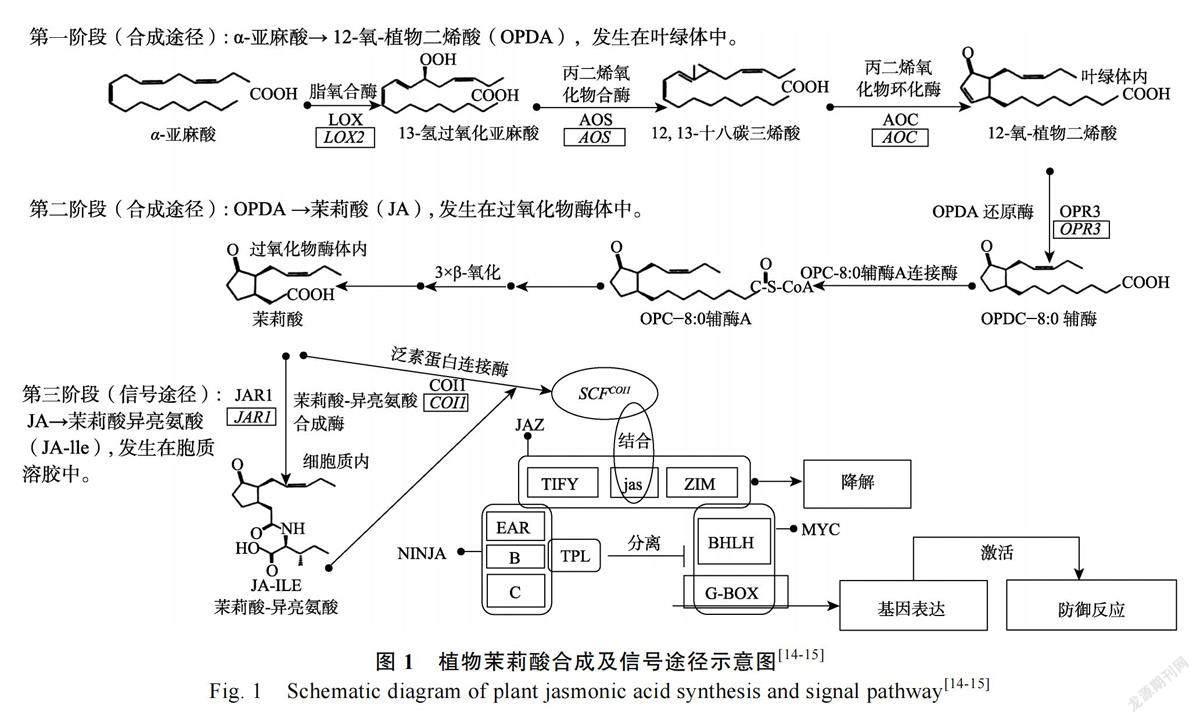
诱导抗性(induced resistance, IR)介导的防御反应在植物抵御植食性昆虫取食为害中发挥重要作用,是害虫绿色防控治理体系中极为重要的一项举措[5],其介导的防御反应主要通过茉莉酸(JA)途径来完成[6]。JA 作为脂质抗性激素在植物抵御害虫胁迫的过程中发挥关键作用[6],害虫取食会激活植物JA 合成途径,促进JA 含量的积累,进一步诱导植物防御反应相关信号途径基因,从而导致植物其他抗虫性物质含量的积累,进而提高植物对害虫的抗性[7]。JA 的合成起源于α-亚麻酸(α-LeA),脂氧合酶(LOX)催化α-亚麻酸并控制JA 合成的第一步。当昆虫取食或为害植物时,植物体内脂氧合酶基因LOX 被激活,誘导JA合成和积累,而生成的JA 又可进一步激活LOX基因[8]。丙二烯氧化物合成酶(AOS)是JA 合成的第2 个关键酶,受JA 的反馈调节,昆虫取食可激活AOS 基因的启动子,造成其在植物体内上调或下调表达[9]。丙二烯氧化物环化酶(AOC)在番茄机械损伤诱导信号中起着关键作用,番茄植株受伤后诱导AOC 在植株体内表达迅速上升[10],表明AOC 可能参与JA 合成。OPDA 还原酶(OPR)相关基因OPR3 的转录受JA 诱导,JA 诱导条件下能促进OPR3 基因表达[11]。JAR1 催化JA 形成共轭物JA-Ile[12],参与JA 信号级联传递。在拟南芥的酵母免疫沉淀实验中证明泛素蛋白(COI1)可能是JA-Ile 的受体[13]。在存在JA-Ile 的情况下,JAZ 蛋白通过Jas 结构域与COI1 结合,JAZ 蛋白随后被降解,NINJA-TPL 复合物与MYC2 转录因子分离,从而启动JA 响应基因的转录,参与植物JA 信号途径介导的防御反应(图1)。
迄今为止,对于桃蚜与辣椒互作时,是否影响JA 合成及信号途径基因表达的蚜口密度和取食时间效应仍不明确。本研究以抗、感蚜辣椒品种为参试材料,系统比较分析不同密度桃蚜取食不同时间后,JA 合成及信号途径基因的表达量在抗、感辣椒品种中的变化趋势及其在抗、感蚜辣椒品种间的差异,以初步明确引起抗、感蚜辣椒品种JA 合成及信号途径基因显著差异表达的蚜口密度和取食时间范围,为阐明JA 合成及信号途径在辣椒抗蚜性中的重要作用以及抗蚜辣椒种质资源鉴定评价提供理论依据与参考指标。
1 材料与方法
1 材料
1.1.1 供试桃蚜 以烟草(Nicotiana tabacum L.)长期室内继代饲养(>54 代)于中国热带农业科学院环境与植物保护研究所特色热带害虫实验室。饲养条件为:温度(24±2)℃,相对湿度为75%±5%,光照为16 h /8 h(L/D)。选择发育历期相同、大小一致的无翅雌成蚜开展后续实验。
1.1.2 供试辣椒 抗蚜辣椒种质猪大肠(ZDC)、感蚜辣椒种质大羊角椒(DYJJ)抗性遗传稳定由本实验室提供,抗、感蚜辣椒品种皆属于长辣椒变种(Capsicum annuum L. var. longum Bailey)。供试猪大肠和大羊角椒辣椒以种子进行有性繁殖,统一育苗,条件设置为:温度24 ℃,光照为16 h/8 h(L/D),相对湿度70%±5%。育苗后3~4片真叶时移栽幼苗至含种植土(土壤∶泥炭土∶珍珠岩=1∶1∶1)的育苗盆中,每周浇水2~3 次,8 周后选取生长发育状况基本一致的健康植株用于实验。
1.2 方法
1.2.1 JA 合成及信号途径基因选择 根据KEGG 网站(https://www.kegg.jp/kegg/)中已经发布的JA 合成及信号通路,并结合相关的文献报道,选取在植物JA 合成及信号调控研究中最为广泛的6 个基因作为候选基因,用于辣椒JA 合成及信号基因表达差异与辣椒抗蚜性的相关显著性分析,分别为LOX2(脂氧合酶基因)、AOS(丙二烯氧化物合成酶基因)、AOC(丙二烯氧化物环化酶基因)、OPR3(OPDA 还原酶基因)、COI1(泛素蛋白连接酶基因)、JAR1(茉莉酸-异亮氨酸合成酶基因)。
1.2.2 辣椒接虫与取样 辣椒移栽8 周后,选取长势基本一致的猪大肠和大羊角椒植株用于实验。每株辣椒选取上部发育成熟和叶片大小相对一致的3 张叶片,用毛笔从烟草上小心挑取桃蚜雌成虫接于叶背。使用蚜虫生态盒轻轻地夹住叶片,使得叶片的正常生理活动不受影响,并且保证桃蚜不迁移(图2)。以未接桃蚜前的辣椒植株相同部位叶片为对照,每张叶的桃蚜密度分别设置为20、30、40、50、60 蚜/叶,每个桃蚜密度和蚜害时间均设置3 个重复。分别在未接桃蚜前(0 h)、接桃蚜后6、12、24、48、72 h 的6 个时间点采集叶片。
1.2.3 RNA 提取及cDNA 第一条链的合成 参照植物总RNA 提取试剂盒(TIANGEN,美国)说明书提取辣椒叶片RNA,利用D2000 DNA 标准分子量Ladder(Solarbio,中国)和琼脂糖凝胶电泳检测RNA 的纯度和完整性。取1.0 μg RNA经ToloScript RT EasyMix for qPCR 试剂(TOLOBIO,中国)消除gDNA 并反转录合成cDNA。
1.2.4 实时荧光定量PCR 分析 根据GenBank中已经发布的辣椒JA 信号途径基因LOX2、AOS、AOC、OPR3、COI1 和JAR1 序列设计qPCR 引物(表1)。cDNA 样品经RNase-free ddH2O 稀释5倍后作为qPCR 的模板,以辣椒CaActin 作为内参基因(表1)。qPCR 反应体系的配制参照2 × Q3SYBR qPCR Master Mix 试剂盒(TOLOBIO,中国)。qPCR 反应条件为:预变性95 ℃ 30 s;循环反应:95 ℃ 10 s,60 ℃ 30 s,40 次循环;溶解曲线:95 ℃ 15 s,60 ℃ 60 s,95 ℃ 15 s。使用LightCycler? 96 仪器(Roche,瑞士)程序采集溶解曲线。分别以未受蚜害的辣椒JA 合成及信号途径基因的表达量归一化设置为1.0,蚜害后的JA基因的表达量变化情况以为害前的相对倍数表示,以LIVAK 等[16]的2?ΔΔCT 方法计算分析,每个处理均设置3 个生物重复,每个生物重复设置3个技术重复。
1.3 数据处理
采用 Excel 软件进行数据汇总整理,使用统计学软件DPS(V15.10)进行不同蚜口密度和不同为害时间的基因表达量差异分析, 采用Duncans 新复极差法方法进行数据间的多重比较(显著性水平均为P=0.05)。
2 结果与分析
2.1 LOX2 表达量差异分析
抗、感蚜辣椒品种在不同密度及不同时间下被桃蚜取食后JA 合成途径基因LOX2 表达量变化差异显著(图3)。除ZDC 在40 蚜口密度外,随蚜口密度的增加和为害时间的延长,抗、感蚜辣椒品种LOX2 表达量在总体上均呈先显著上升再显著后下降的趋势;除60 蚜口密度外,抗、感蚜品种中LOX2 表达量均在不同为害密度的同一为害时间下达到峰值;其中抗、感蚜品种中LOX2表达量均在30 蚜口密度下为害48 h 达到峰值,较对照分别增长11.79 倍及10.96 倍(图3A、图3B)。进一步比较抗、感品种之间LOX2 的表达量差异,发现在20、50 及60 蚜口密度下,ZDC 表达量显著低于DYJJ,而在30 及40 蚜口密度下,ZDC 表达量显著高于DYJJ(图3C)。
2.2 AOS 表达量差异分析
抗、感蚜辣椒品种在不同密度及不同时间下被桃蚜取食后JA 合成途径基因AOS 表达量变化差异显著(图4)。除40 蚜口密度外,随蚜口密度的增加和为害时间的延长,抗、感蚜辣椒品种AOS 表达量在总体變化趋势上基本一致;在50蚜口密度下,抗、感品种表达量差异最显著,均在为害6 h 达到峰值,较对照分别增长6.91 倍及14.27 倍(图4A、图4B)。进一步比较抗、感品种之间AOS 的表达量差异,发现除50 蚜口密度外,其余蚜口密度下ZDC 表达量均显著高于DYJJ(图4C)。
2.3 AOC 表达量差异分析
抗、感蚜辣椒品种在不同密度及不同时间下被桃蚜取食后JA 合成途径基因AOC 表达量变化差异显著。除40 蚜口密度外,随蚜口密度的增加和为害时间的延长,抗、感蚜辣椒品种AOC 表达量在总体变化趋势上基本一致,呈显著上升再显著下降,再略微回调最终下降的趋势;其中抗、感蚜品种中AOC 表达量均在30 蚜口密度下为害6 h 达到峰值,较对照分别增长1.34 倍及1.41 倍;(图5A、图5B)。进一步比较抗、感、品种之间AOC 的表达量差异,发现在20、30、40、50 和60 蚜口密度下,ZDC 表达量均显著高于DYJJ(图5C)。
2.4 OPR3 表达量差异分析
抗、感蚜辣椒品种在不同密度及不同时间下被桃蚜取食后JA 合成途径基因OPR3 表达量变化差异显著(图6)。随蚜口密度的增加和为害时间的延长,抗、感蚜辣椒品种OPR3 表达量在总体上均呈先显著上升再显著下降的趋势;除50 及60 蚜口密度外,抗、感蚜品种中OPR3 表达量均在不同为害时间下达到峰值;其中抗、感蚜品种中OPR3 表达量均在50 蚜口密度下为害6 h 达到峰值,较对照分别增长1.31 倍及7.92 倍(图6A、图6B)。进一步比较分析抗、感品种之间OPR3的表达量差异,发现在20、50 及60 蚜口密度下,ZDC 表达量显著低于DYJJ,而在30 及40 蚜口密度下,ZDC 表达量显著高于DYJJ(图6C)。
2.5 COI1 表达量差异分析
抗、感蚜辣椒品种在不同密度及不同时间下被桃蚜取食后JA 信号途径基因COI1 表达量变化差异显著。随蚜口密度的增加和为害时间的延长,抗、感蚜辣椒品种COI1 表达量在总体变化趋势上基本一致,呈现先下降后上升最终下降;在40蚜口密度下,抗感品种表达量差异最显著,均在为害24 h 达到谷值,比对照分别降低0.52 倍及2.73 倍(图7A、图7B)。进一步比较抗、感品种之间COI1 的表达量差异,发现在50 蚜口密度下,ZDC 表达量均显著高于DYJJ,而在20、30、40及60 蚜口密度下,ZDC 表达量均显著低于DYJJ(图7C)。
2.6 JAR1 表达量差异分析
抗、感蚜辣椒品种在不同密度及不同时间下被桃蚜取食后JA 信号途径基因JAR1 表达量变化差异显著(图8)。随蚜口密度的增加和为害时间的延长,抗、感蚜辣椒品种JAR1 表达量在总体变化趋势上基本一致,呈现先上升再下降的趋势;在60 蚜口密度下,抗、感品种表达量差异最显著,均在为害72 h 达到谷值,较对照分别降低1.83倍及1.75 倍(图8A、图8B)。进一步比较抗、感品种之间JAR1 的表达量差异,发现在各个蚜口密度下,ZDC 表达量均显著低于DYJJ(图8C)。
3 讨论
JA 介导的应对植食性昆虫的防御途径在植物免疫防御反应中发挥重要作用[17],JA 是诱导作物体内系统抗性产生抗虫反应的关键信号分子,JA 信号通路又在植物防御的激活中起着主导作用,且JA 合成及信号防御途径由多基因控制[18-19]。植物主要通过JA 途径介导的防御反应抵御刺吸式害虫,刺吸式害虫的虫口密度和为害时间能显著影响作物JA 合成途径基因的表达[20]。
本研究发现,抗、感蚜辣椒品种在不同蚜口密度为害下,LOX2 表达量均呈现先显著升高后随为害时间的延长逐渐恢复至为害前水平。LOX 基因表达量呈现相似的变化。同样在马铃薯蚜虫取食番茄叶片和桃蚜取食拟南芥叶片[21],桃蚜取食抗、感辣椒叶片[22]以及二斑叶螨取食抗、感木薯品种[23]中得以报道。这些研究表明LOX2 基因作为JA 合成中途径的上游调控基因在植物应答昆虫取食为害中发挥重要作用。在抗、感大豆材料抗虫差异中研究发现,大豆的抗性差异可能与AOS基因的表达密切相关[24]。AOS 基因可能参与了水稻抗二化螟、稻纵卷叶螟、褐飞虱的防御反应,沉默获得的AOS 水稻品系及其T1 代种子证明了AOS 在水稻抗虫防御中起到了积极防御作用[25]。
本研究发现,AOS 基因表达量在抗、感蚜辣椒品种中随为害时间的延长呈现明显的先上升后降低的趋势;50 蚜口密度下为害6 h 时感蚜辣椒品种相比对照增长14 倍之多,随后剧烈下降,表明感蚜品种DYJJ 体内AOS 基因在应对桃蚜的取食为害时转录水平变化剧烈,这可能是感虫品种中JA初始含量较低,为了应对抑制蚜害的需要从而通过AOS 基因的快速上调最终响应其体内JA 快速合成,但这种高水平的转录并不持久。本研究中,在不同蚜口密度下,抗蚜辣椒品种的AOC 表达量相对感蚜品种随为害时间的延长下降速度较为缓慢,且在抗蚜品种中表达量显著高于感蚜品种。与AOC 在甘蔗抗粘虫的表达研究中基本一致[26]。在本研究中OPR3 均随为害时间的延长,在感蚜辣椒品种DYJJ 中的表达量显著低于抗蚜品種ZDC。在水稻抗虫性研究中也发现OPR3 在中抗水稻品种中能够过量表达,增强对二化螟的抗性,OPR3 的转录受JA 诱导,在创伤等能够诱导JA的胁迫反应,而且也能促进OPR3 基因表达[27]。水稻COI1 基因沉默的突变株实验表明了其COI1基因在水稻防御反应中发挥关键作用[28]。本研究发现COI1 和JAR1 的表达量在抗虫品种中显著低于感虫品种,可能COI1 和JAR1 在抗、感应答桃蚜取食为害中具有较高的协同作用,桃蚜取食为害对抗蚜辣椒品种的COI1 和JAR1 基因表达抑制作用较强,推测该基因可能在辣椒对桃蚜的防御反应中也扮演重要角色。
通过分析上述6 个JA 途径基因表达量,发现桃蚜取食抗、感辣椒品种在不同蚜口密度和为害时间下,JA 合成途径相关基因LOX2、AOS、AOC、OPR3 在抗蚜辣椒品种ZDC 中的表达量总体上显著高于感蚜品种DYJJ。推测JA 合成途径基因调控介导的防御反应在抗虫品种ZDC 中可能比感蚜品种DYJJ 的应答桃蚜取食为害时作用更直接、强烈。与之相似的,有研究表明烟粉虱在不同虫口密度下取食抗、感辣椒品种植株时,抗性品种内LOX 及AOC 的表达量也显著高于感虫品种[29]。此外,同一蚜口密度下各个基因表达量的峰值和谷值在抗、感辣椒品种中出现的时间基本一致,但随着蚜口密度的上升,抗、感辣椒品种各基因表达量峰值的出现在时间上均有明显前移的趋势,推测可能随着桃蚜虫口数量的上升,对植物迅速产生了更强烈的胁迫压力,导致在较高蚜口密度下辣椒JA 途径基因表达量的峰值出现在较早为害时间内。这一结果与WANG 等[30]的研究相似,即高水平的食草强度将持续增加植物初始诱导抗性。
本研究初步阐明了基于JA 合成及信号途径的辣椒抗蚜性分子机理,同时表明AOS、AOC 及JAR1 基因可能具有作为鉴定评价辣椒抗蚜性水平的分子指标的潜力以及为辣椒抗蚜种质资源评价与创新利用提供理论依据。然而,本研究仅比较了单个抗蚜和感蚜辣椒品种之间的基因表达差异,上述评价指标在抗、感蚜辣椒品种群体中是否具有通用性和适应性仍需进一步证实。
