河北省石家庄市番茄黄化曲叶病毒及其伴随DNAβ 的分子鉴定及序列分析
2023-05-12韩旭良康忱田哲娟王鹏李亚栋赵雪芳闫洪波刘美霞吴志明
韩旭良,康忱,田哲娟,王鹏,李亚栋,赵雪芳,闫洪波,刘美霞,吴志明*
(1.河北经贸大学,河北 石家庄 050051;2.河北省农林科学院经济作物研究所,河北 石家庄 050051;3.山东省寿光市圣城街道办事处,山东 寿光 262700)
双生病毒(geminivirus)是一种具有孪生颗粒形态的单链DNA 植物病毒[1],在世界范围内普遍发生[2],感病植株主要表现为叶片卷曲窄小、皱缩,叶片和叶脉黄化,茎弯曲,植株生长缓慢、矮化等[3,4]。该病毒已在我国云南、广西、广东、海南、河南、山东、河北等地区的番茄、烟草、番木瓜等多种经济作物上侵染,造成重大的经济损失[5,6]。双生病毒具有长度为2.5~3.2 kb(单组分病毒,只含有DNA-A)和4.8~5.6 kb(双组分病毒,同时含有DNA-A 和DNA-B)的环状基因组,根据昆虫载体和基因组特征的不同,这些病毒可分为7 个属——贝库特病毒属(Becurtovirus)、贝戈莫病毒属 (Begomoviru)、曲顶病毒属 (Curtovirus)、埃农病毒属(Eragrovirus)、马斯特里韦病毒属(Mastrevirus)、拓扑病毒属(Topocuvirus)和特恩库特病毒属 (Turncurtovirus)[7,8]。其中贝库特病毒属病毒(Begomovirus)可以通过烟粉虱进行大范围传播,造成了巨大的经济损失[9,10]。菜豆金色花叶病毒属病毒大多数包含2 条大小相近的组分DNA-A 和DNA-B,其基因组长度为2.5~3.0 kb,无包膜;少数为单组分,只含有DNA-A,长度约为2.8 kb[11]。番茄黄化曲叶病毒(tomato yellow leaf curl virus,TYLCV)是单组分菜豆金色花叶病毒的典型代表,被其侵染的番茄植株会出现叶片卷曲窄小且叶脉泛黄、植株矮小、发育缓慢等症状,严重影响番茄生产[12]。
近年来国内外[13]均发现一些单组分双生病毒伴随有卫星 DNA(DNA Betasatellite,又称 DNAβ),而DNAβ 只有在DNA-A 存在的情况才可以进行复制和在植物细胞组织间移动[14]。不同双生病毒伴随的DNAβ存在较大差异,目前尚无合适的方法对其进行归类[15]。伴随TYLCV 的DNAβ 是一种大小约为1.3 kb 的亚病毒单链环状DNA 分子,该DNAβ 可编码BC1 蛋白[16],而这种蛋白对菜豆金色花叶病毒属部分病毒的致病性具有重要影响[17,18]。
TYLCV 及其DNAβ 对番茄生产有重大影响,对石家庄TYLCV 及其DNAβ 混合侵染情况进行调查并对其传播途径和遗传变异情况进行分析,对指导石家庄番茄生产工作具有重要意义。为了明确TYLCV 及其DNAβ 在河北省石家庄市番茄上的侵染状况与进化起源,对采自石家庄市的感病番茄植株样品进行病毒检测,发现引起石家庄市番茄发生病变的病毒为TYLCV,其中部分TYLCV 分离物伴随有DNAβ。对TYLCV 病毒分离物及其DNAβ 分子进行遗传变异分析,并明确其传播途径,旨为双生病毒及其DNAβ 在河北省的传播和防控提供科学依据。
1 材料与方法
1.1 试验材料
2021~2022 年在河北省农林科学院经济作物研究所的石家庄市大河实验基地,选择出现叶片卷曲皱缩、生长缓慢、黄化、茎弯曲、植株矮化等症状的番茄植株采集4 份叶片,依次编号为1~4。以健康植株叶片为阴性对照;以TYLCV 侵染性克隆浸润法接种的发病番茄植株叶片为阳性对照。
试剂主要有植物基因组提取试剂盒(Plant DNA Isolation Kit,成都福际生物技术有限公司生产) 和pEASY-T1 试剂盒(北京全式金生物技术股份有限公司生产)。
1.2 试验方法
1.2.1 叶片总DNA 提取 将叶片经液氮速冻研磨后,按照植物基因组提取试剂盒说明书上的具体操作,提取番茄叶片基因组DNA;然后,将DNA 溶解于无菌去离子水中,置于-20 ℃冰箱内保存,备用。
1.2.2 引物设计与合成 使用彭燕等[19]设计的TYLCV检测引物(表1)扩增TYLCV 的DNA-A 部分区域后进行测序,根据测序结果设计全长扩增引物扩增DNA-A 全长序列,以总DNA 为模板进行PCR 扩增。PCR 反应条件:94 ℃预变性 5 min,94 ℃变性 30 s,56 ℃退火 30 s,72 ℃延伸 2 min 30 s,35 个循环;72 ℃终延伸10 min。
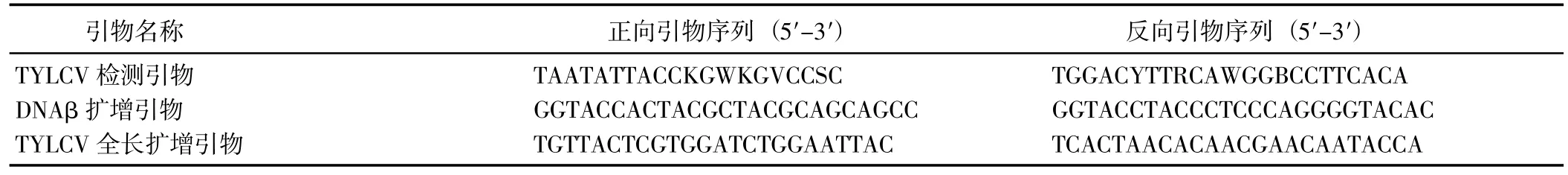
表1 引物信息Table 1 Primer information
使用谢艳等[20]设计的DNAβ 扩增引物扩增DNAβ,以总DNA 为模板进行PCR 扩增。PCR 反应条件:94 ℃预变性 5 min;94 ℃变性 30 s,65 ℃退火 30 s,72 ℃延伸 1 min 15 s,35 个循环;72 ℃终延伸 10 min。
PCR 产物经1%琼脂糖凝胶电泳后进行胶回收。将胶回收产物连接至pEASY-T1 载体上,转化到DH5α 大肠杆菌感受态细胞中,在含有卡那霉素的LB固体培养基中生长12~16 h 后挑取单克隆,并将3 个鉴定为阳性的单克隆交由华大基因公司进行测序。根据测序结果设计反向引物进行TYLCV 病毒扩增,得到TYLCV 石家庄分离物的DNA-A 近全长序列。
1.2.3 序列分析 在NCBI 上选取国内外有代表性的TYLCV DNA-A 序列 (表 2) 和 DNAβ 序列 (表 3),使用MEGA 6.0 软件中的Compute pairwise distance法进行成对序列距离分析,使用MEGA 6.0 软件中的Clustal W 法进行多序列比较分析,然后进行进化树的构建。使用DNAMAN 软件将本次得到的分离物与2015 年TYLCV 石家庄分离物进行序列比对,进一步分析石家庄TYLCV 的变异情况。
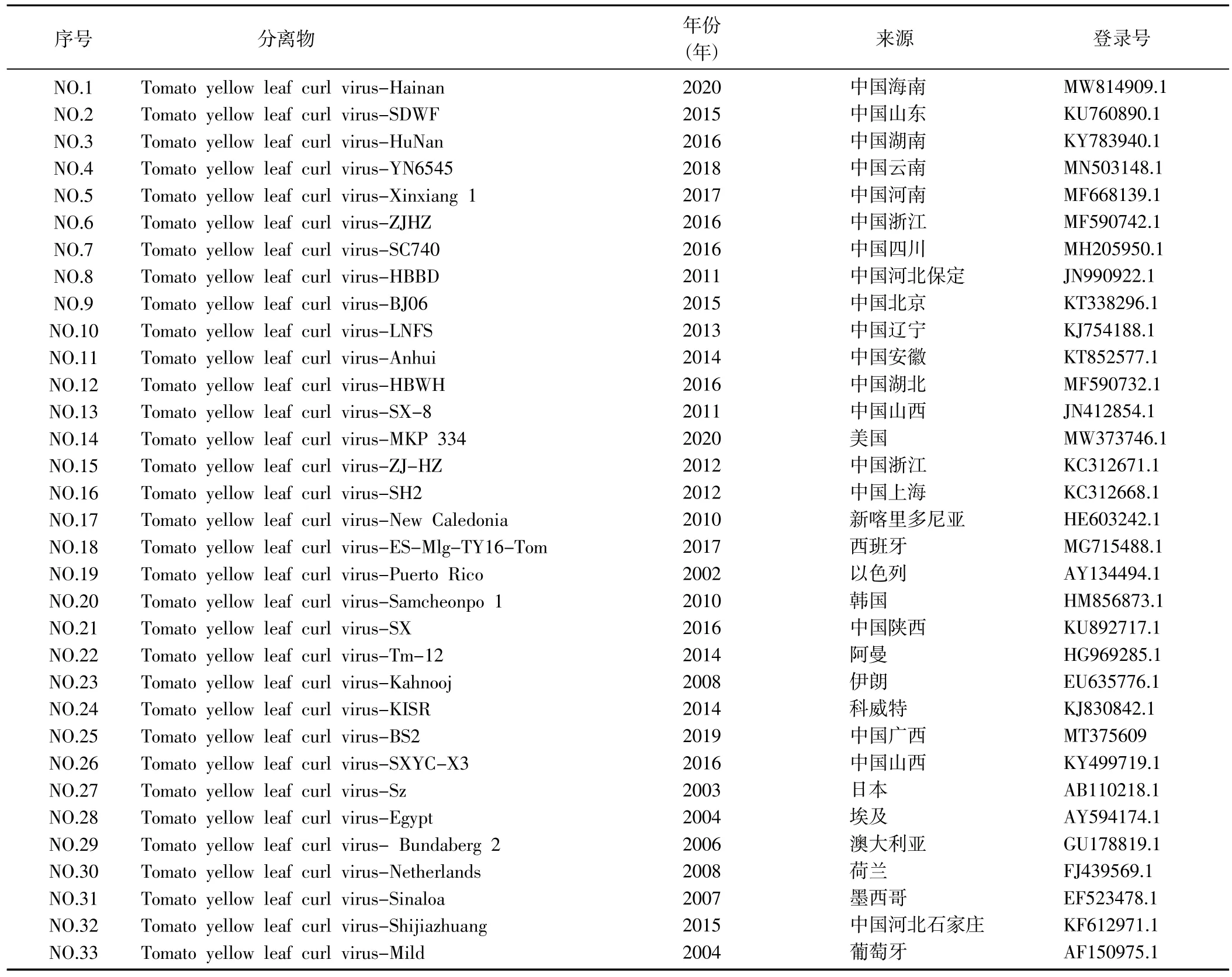
表2 不同年份各地区TYLCV 分离物及其在国际收录库的收录情况Table 2 TYLCV isolates and their inclusion in international databases in different years and regions
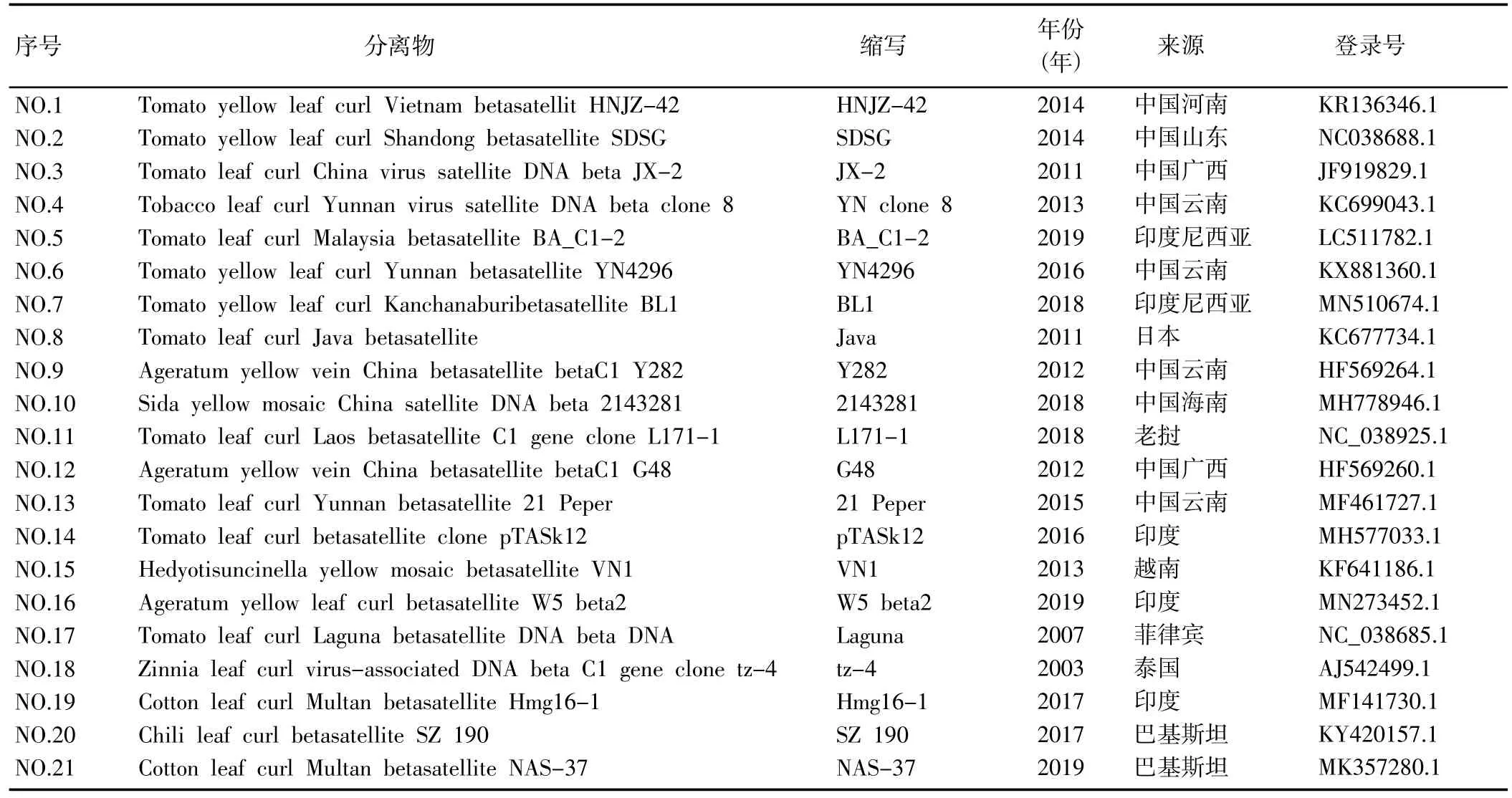
表3 不同年份各地区双生病毒属DNAβ及其在国际收录库的收录情况Table 3 DNA Betasatellite of geminivirus in different years and regions and their inclusion in the international database
2 结果与分析
2.1 TYLCV 石家庄分离物基因组DNA-A 结构特征
使用TYLCV 检测引物对4 个样品基因组DNA 进行扩增,均得到长度约为500 bp 的条带(图1);测序结果表明,4 个样品感染的TYLCV 序列完全一致。根据测序结果设计全长扩增引物,得到TYLCV 石家庄分离物的全长序列,将其命名为TYLCV-SJZ2021。该序列全长为2 781 bp,与已报道的TYLCV 具有相同的结构特征,共有6 个ORFs——编码外壳蛋白的AV1(308 ~1 084 bp),编码移动相关蛋白的 AV2(148~498 bp),编码复制蛋白的 AC1(1 542~2 615 bp),编码转录激活因子的AC2(1 226~1 633 bp),编码复制增强因子的AC3(1 081~1 485 bp),编码复制和转录调控因子的AC4(2 171~2 464 bp)。除此之外,在非编码区(2 616~147 bp)上含有与双生病毒复制起始相关的保守序列TAATATTAC(2 775~2 bp)。
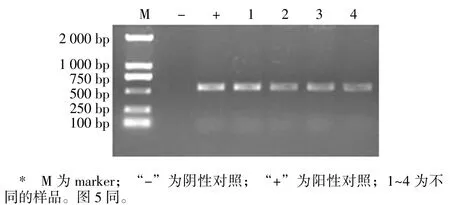
图1 番茄TYLCV 石家庄分离物基因组全长扩增结果Fig.1 Full-length amplification of TYLCV Shijiazhuang isolate
2.2 TYLCV 石家庄分离物基因组DNA-A 相似性比较分析
为了解TYLCV-SJZ2021 在病毒上的分类地位和可能来源,对得到的TYLCV 分离物进行成对序列距离分析(图2)和系统进化树分析(图3)。根据成对序列距离分析结果,可以将本研究选用的TYLCV 病原分离物分为5 个株系——伊朗株系、科威特株系、阿曼株系、以色列株系和葡萄牙株系,TYLCVSJZ2021 与伊朗株系、科威特株系和阿曼株系的相似度均在94%以下,与以色列株系的相似度在97%以上,因此认为TYLCV-SJZ2021 属于以色列株系。

图2 石家庄TYLCV-SJZ2021 与其他TYLCV 分离物成对序列距离分析Fig.2 Pairwise sequence distance analysis of TYLCV-SJZ2021 and other TYLCV isolates
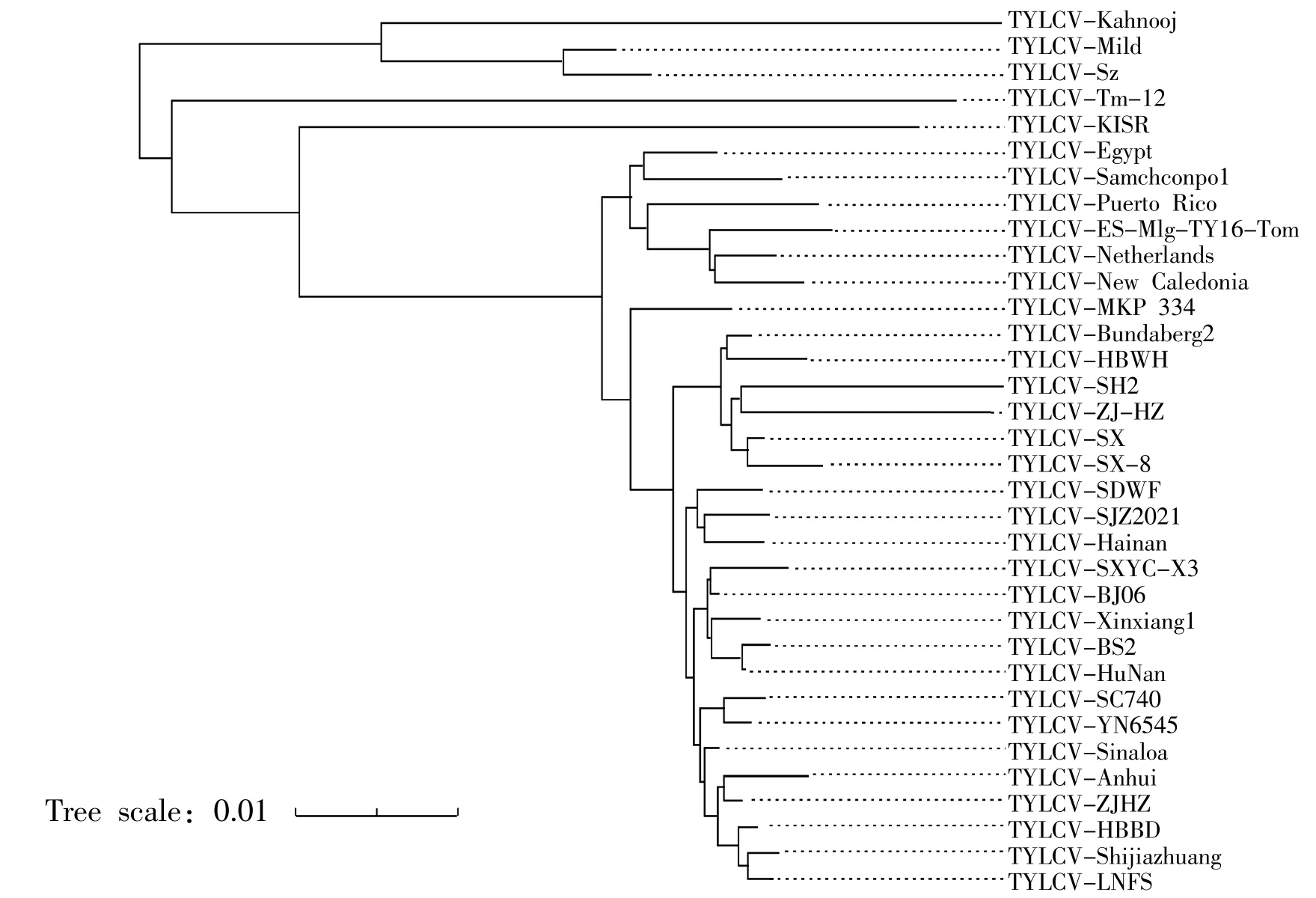
图3 石家庄TYLCV-SJZ2021 与其他TYLCV 分离物的系统进化树Fig.3 Phylogenetic tree of TYLCV-SJZ2021 and other TYLCV isolates
系统进化树分析结果表明,TYLCV-SJZ2021 与国内17 个地区分离物以及澳大利亚和墨西哥TYLCV 分离物聚集在一个分支上,进一步与美国TYLCV 分离物形成一个较大的分支;与新喀里多尼亚分离物、西班牙分离物、以色列分离物、韩国分离物、埃及分离物、科威特分离物和荷兰分离物亲缘关系较近,与伊朗分离物、科威特分离物、阿曼分离物、日本分离物和葡萄牙分离物亲缘关系较远。
将TYLCV-SJZ2021 与2015 年本实验室得到的TYLCV 石家庄分离物[21]进行对比,结果(图4)显示,二者相似度为98.67%,有37 处碱基不同,分别位于13、17、27、28、38、40、42、57、136、146、190、348、393、499、532、595、745、790、880、1 248、1311、1359、1458、1506、1627、1800、1806、1827、1941、2241、2255、2377、2402、2421、2493、2689和2 728 bp。其中,AV1 编码区有8 处碱基不同,AV2 编码区有1 处碱基不同,AC1 编码区有11 处碱基不同,AC2 编码区有6 处碱基不同,AC3 编码区有4 处碱基不同,AC4 编码区有5 处碱基不同;非编码区有2 处碱基不同。

图4 2021 年与2015 年得到的石家庄番茄TYLCV 分离物序列比对Fig.4 Sequence alignment of Shijiazhuang TYLCV isolates in 2015 and 2021
2.3 TYLCV-SJZ2021 伴随的DNAβ分子结构及同源性比较
使用DNAβ 扩增引物扩增DNAβ 的全长序列(图5),将其命名为Betasatellite SJZ2021。测序结果表明,4个感染TYLCV 样品所携带的DNAβ 序列也完全相同。Betasatellite SJZ2021 全长为1 343 bp,其上有编码病毒在细胞间移动相关蛋白的BC1 编码区(279~629 bp);有 1 个腺嘌呤 (A) 富集区 (A-rich,798~1 058 bp),A 最高重复数为13(962~974 bp),该区域对保证DNAβ 分子大小有重要作用;有1 个DNAβ 保守区域(SCR) 和 1 个双生病毒科病毒共有的茎环结构TAATATTAC (60~68 bp)。
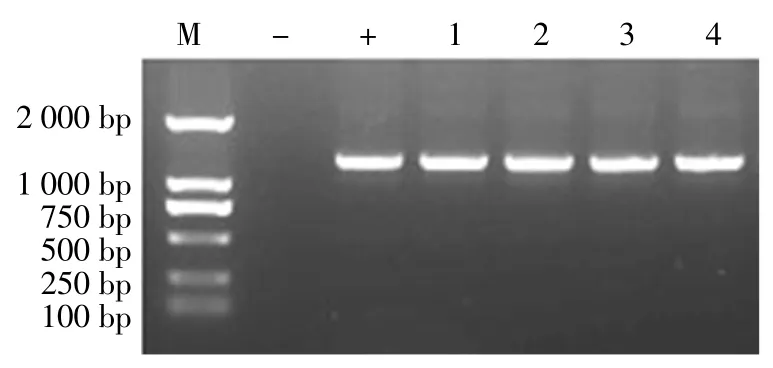
图5 TYLCV-SJZ2021 伴随DNAβ的全长扩增结果Fig.5 The DNAβfull-length amplification result of TYLCV-SJZ2021
对得到的DNAβ 进行成对序列距离分析,结果(图6)表明,Betasatellite SJZ2021 与山东分离物和河南分离物相似度(98%)最高,与其他地区双生病毒DNAβ 分离物相似度均在78%以下。
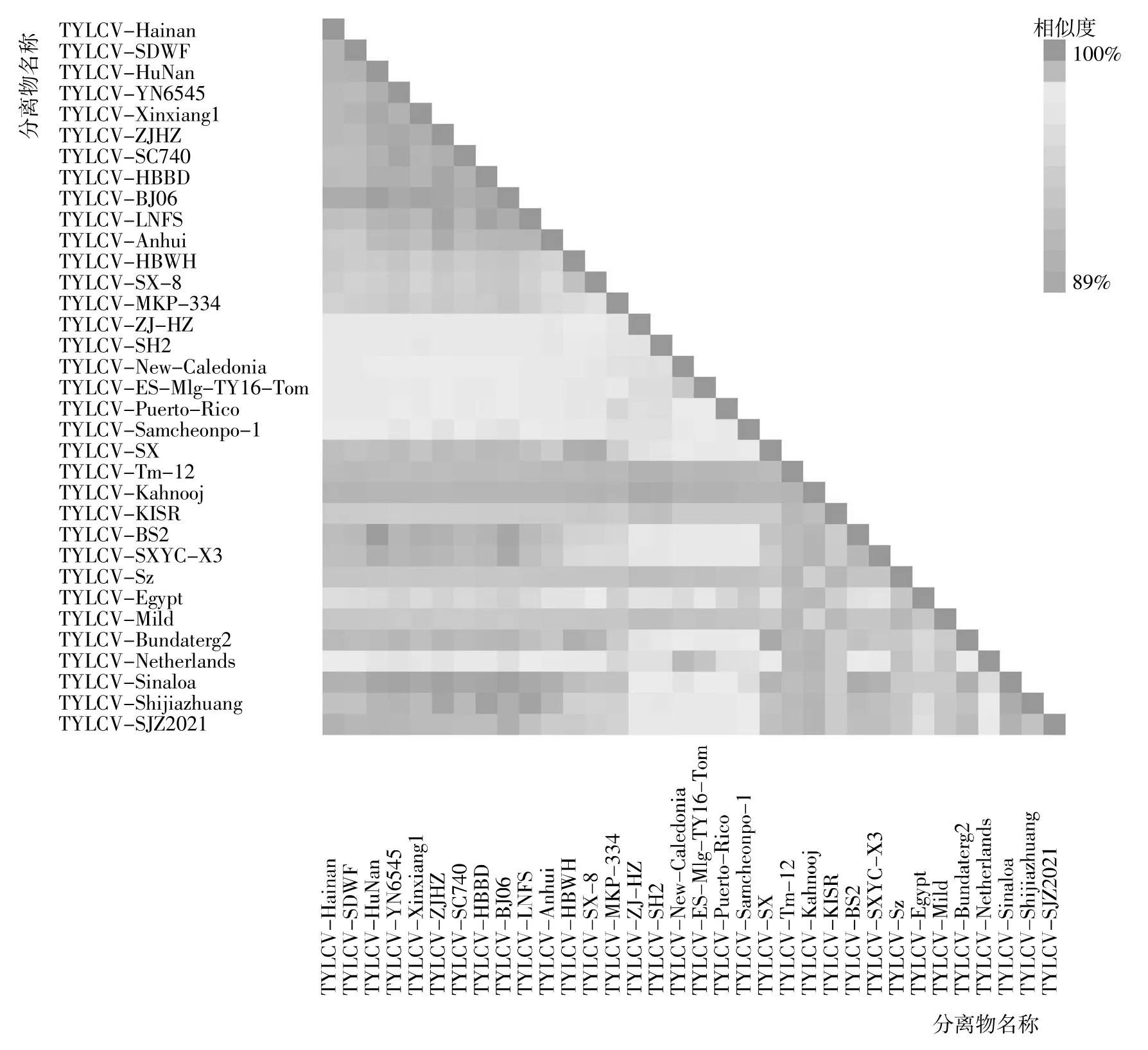
图6 TYLCV-SJZ2021 伴随DNAβ与其他双生病毒属伴随DNAβ成对序列距离分析Fig.6 Pairwise sequence distance analysis between TYLCV-SJZ2021associated DNA betasatellite and other DNA betasatellite of geminivirus
从系统进化树(图7)上来看,TYLCV-SJZ2021伴随DNAβ 与山东地区和河南地区分离物形成一个分支;印度尼西亚与云南地区2 个不同年份的分离物形成一个分支,进而与石家庄、山东和河南地区分离物形成一个较大分支。由此推测,TYLCV-SJZ2021 伴随DNAβ 很有可能由印度尼西亚传入我国云南地区,再由云南地区传入石家庄市。
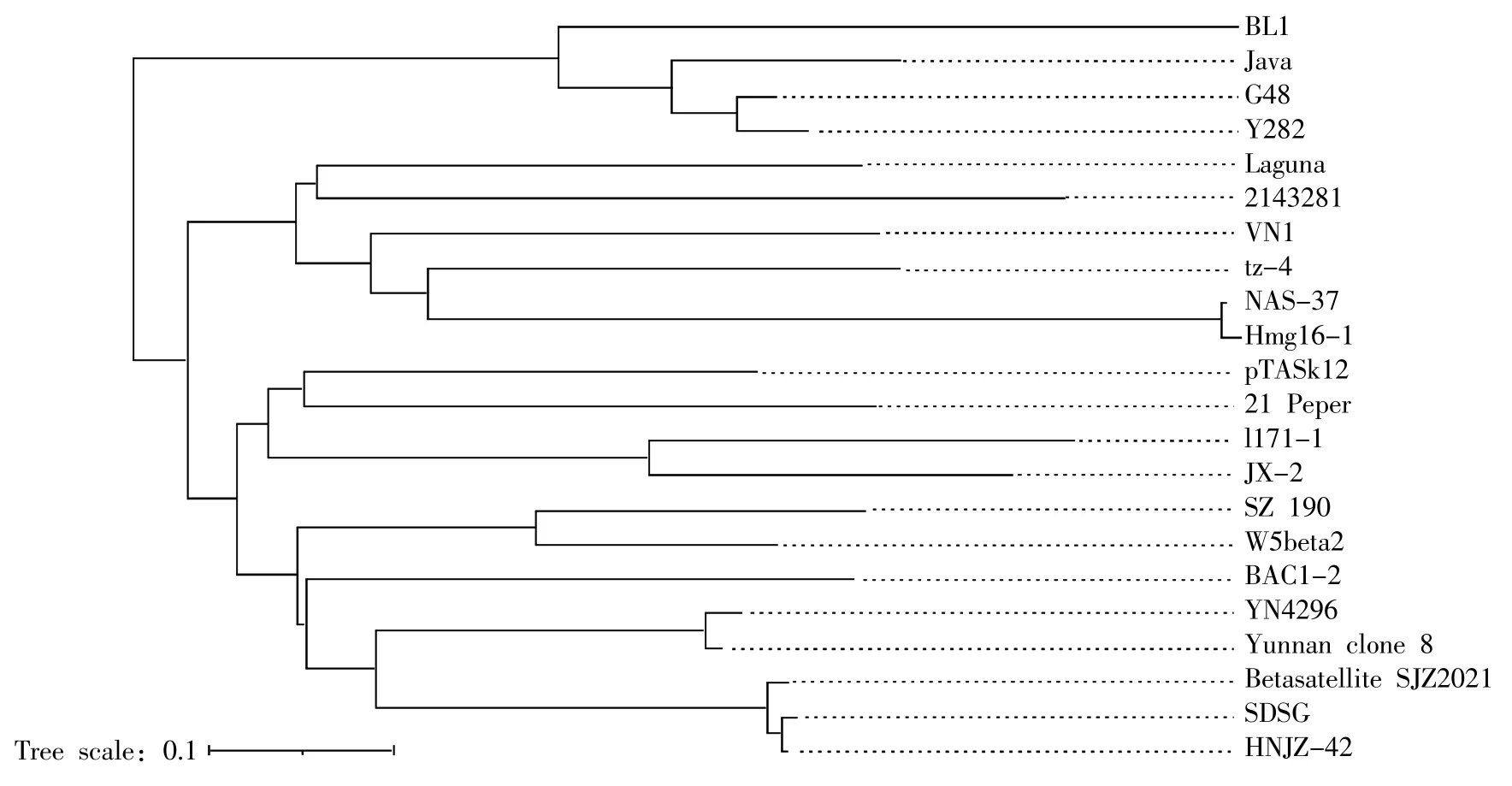
图7 TYLCV-SJZ2021 伴随DNAβ与其他双生病毒属伴随DNAβ的系统进化树Fig.7 Phylogenetic tree of TYLCV-SJZ2021 associated DNA betasatellite and other DNA betasatellite of geminivirus
3 结论与讨论
本研究从分子水平上明确了近几年河北省石家庄市为害番茄的TYLCV-SJZ2021 分离物在geminivirus中的分类地位,并将其归类为以色列株系。双生病毒科病毒在相似度>94%时被认为是同一株系的不同变种,相似度为89%~94%时被认为是不同株系,相似度<89%时则被认为是不同的病毒[22]。本文中的TYLCVSJZ2021 与2015 年山东TYLCV 分离物相似度在99%以上,二者亲缘关系最近,推测可能是通过烟粉虱传播或者在引进山东植物品种时传入;且与本实验室白晓娟等[21]和河北科技大学田鹏等[23]关于河北TYLCV 株系鉴定结果一致,与以色列株系相似度为99%,说明近几年为害河北省的TYLCV 病毒株系主要是TYLCV以色列株系。但是与上述TYLCV 株系鉴定不同的是,上述两者的报道中均明确提出TYLCV 分离物只含有DNA-A 部分,不含有TYLCV 伴随DNAβ 分子,而本研究首次从石家庄番茄病区检测到TYLCV-SJZ2021的伴随DNAβ 分子。本研究得到的TYLCV-SJZ2021与2015 年本实验白晓娟等[21]得到的分离物相比编码区和非编码区都产生了较多变异,这种变异可能与TYLCV DNA-A 分子与其伴随DNAβ 分子的进化及其致病性有关系,伴随DNAβ 促进了感染TYLCV 病毒症状发生的严重程度。
本双生病毒的伴随DNAβ 的变异速度较快。DNAβ 在第一次被报到时只有682 bp,且为无可预测的ORF,且不会对TYLCV 的症状和感染速度产生影响[24]。在2008 年全世界已被报道的双生病毒伴随DNAβ变种超过了50 种且长度达到1 kb 以上[25],2020 年全世界已被报道的伴随DNAβ 变种更是超过了60 种[16]。以本文选取的云南地区TYLCV 分离物伴随DNAβ 分子为例,2012 年长度为1 218 bp[26],2013 年长度为1 349 bp,2015 年长度为 1 360 bp ,2016 年长度为 1 353 bp,DNAβ 因其特性常以78%的相似度作为物种界限,云南分离物除2013 年分离物与2016 年分离物相似度为97%外,其他年份分离物两两对比的相似度远小于78%。在2021 年以前,河北省未有关于TYLCV 伴随卫星DNAβ 分子的报道,结合本文TYLCV-SJZ2021伴随DNAβ 与河南报道的TYLCV 伴随DNAβ 分子相似度(98%)最高,推测该病毒是较近时间内通过烟粉虱由河南省传播至河北省。DNAβ 与不同双生病毒复合侵染不同植物时会产生的作用不同,例如,DNAβ 分子与棉花曲叶热兹拉病毒(cotton leaf curl Gezira virus,CLCuGV)复合侵染棉花时会参与病症的诱导[27],与中国番茄黄化曲叶病(tomato yellow leaf curl China virus,TYLCCNV)或TYLCV 复合侵染番茄时和与秋葵黄皱病毒(okra yellow crinkle virus,OYCrV)复合侵染秋葵时,DNAβ 会引起特定的症状[12,28]。随着DNAβ 的变异,其可能会对番茄及其他农作物产生不确定的为害,因此,定期对其进行检测和分子鉴定,对番茄及其他农作物的生产具有重要意义。
本研究中,根据国内报道的TYLCCNV 和中国番木瓜曲叶病毒(papaya leaf curl China virus,PaLCuCNV)及其伴随DNAβ 进行了检测,但并未检测到TYLCCNV 和PaLCuCNV,说明当前为害河北省石家庄市番茄作物的TYLCV-SJZ2021 主要是TYLCV 以色列株系及其伴随DNAβ 分子共同作用引起的,这为开展河北省番茄抗病材料种质资源鉴定和培育抗TYLCV品种奠定了基础。TYLCV 是导致番茄及其他部分作物减产的重要病毒之一,而伴随DNAβ 可以增加其致病性,随着DNAβ 的变异还会产生不可预测的影响,本文从分子水平上明确了TYLCV-SJZ2021 及其伴随DNAβ 的基因组序列特征和分类地位,为进一步利用其全长序列构建病毒侵染性克隆奠定了材料基础,对于开展抗病毒育种材料接种和加快培育出适合当地种植的番茄抗病新品种具有重要理论和应用价值。
