分枝期水盐胁迫对紫花苜蓿生长及生理特征的影响
2023-05-09田德龙侯晨丽徐冰任杰张琛
田德龙,侯晨丽,徐冰,任杰,张琛
分枝期水盐胁迫对紫花苜蓿生长及生理特征的影响
田德龙1,侯晨丽2,徐冰1,任杰1,张琛1
(1.水利部牧区水利科学研究所,呼和浩特 010020;2.内蒙古科技大学 能源与环境学院,内蒙古 包头 014010)
【目的】探讨分枝期水盐胁迫对紫花苜蓿生长及生理特征的影响。【方法】以紫花苜蓿为研究对象,于2018—2019年在内蒙古磴口县圣牧高科经济园区内开展盆栽试验,设置3个水分胁迫处理:W1(55%f~70%f)、W2(70%f~85%f)、W3(85%f~100%f),并设置4个盐分胁迫处理:S0(<1.0 g/kg)、S1(1.5 g/kg)、S2(3.0 g/kg)、S3(4.5 g/kg),在苜蓿初花期测量各处理的地上部分、根系生物量以及地上部分植株渗透调节物质和抗氧化酶活性,探究水盐胁迫对紫花苜蓿水分消耗、生长及生理特性的影响。【结果】与单一水分和盐分胁迫相比,水盐胁迫导致苜蓿耗水量和地上生物量进一步降低。在低水处理下,添加适宜的盐分(≤3 g/kg)可提高苜蓿自身的脯氨酸量、Na+量以及过氧化物酶(POD)活性,降低超氧化物歧化酶(SOD)活性,缓解干旱对苜蓿的氧化损害,促进苜蓿生长。其中,S2W1处理下的茎叶脯氨酸量、Na+量及POD活性相比S0W1处理提高74.28%、17.28%和34.60%,SOD活性减少9.87%,耗水量、地上部分生物量相比S0W1处理分别增加6.48%和2.17%。【结论】干旱和盐分复合胁迫条件下,紫花苜蓿表现出一定的交叉适应现象,盐分质量分数在2~3 g/kg、灌水量在田间持水率的70%~85%时,最适宜苜蓿生长。
水盐胁迫;紫花苜蓿;生物量;渗透调节物质;抗氧化酶活性
0 引言
【研究意义】河套灌区作为内蒙古自治区重要的商品粮油生产基地,多年来灌区种植结构单一,加之水资源短缺和土壤盐渍化问题,严重制约了农牧业的发展。紫花苜蓿作为一种优质的豆科牧草,产量高、营养价值好、抗逆性强。然而,分枝期水盐胁迫对紫花苜蓿生长及生理特征的影响尚不清晰,制约了盐渍化地区苜蓿的推广和高质量生产。【研究进展】植物在萌发和幼苗生长初期对盐分胁迫最为敏感,减产风险更大[1-2]。然而,以往研究认为植物对非生物胁迫的响应在整个生长周期中是不同的,提高植物存活率也并不一定意味着可以获得更高的产量[2]。Singh等[3]研究发现,在玉米营养和生殖生长阶段,水分亏缺下植物抗氧化防御系统的能力下降是导致产量下降的主要原因。因此,提高作物关键生育期的耐受性是保障粮食产量的重要途径。游离脯氨酸作为公认的植物渗透调节物质,可平衡液泡渗透势,减小逆境对植株的伤害程度,叶片游离脯氨酸的积累可通过调节糖代谢使植物免受氧化损伤,有助于维持碳同化[2-4]。此外,超氧化物歧化酶(SOD)和过氧化物酶(POD)也是保护细胞免受氧化损伤的主要胞内酶,可迅速清除活性氧(ROS)[4],减少氧化损伤。研究表明,脯氨酸的增加明显提高了植物的耐受性;水分胁迫下,植物会积累大量的脯氨酸进行渗透调节[2-4]。柳燕兰等[5]研究发现,重度干旱胁迫下苗期玉米通过增加根源脱落酸(ABA)量,使叶片抗氧化酶活性提高,同时最大电子传递速率、最大羧化速率、磷酸丙糖利用率及RuBP羧化酶的羧化效率降低,使单株生物量相比正常供水条件下降低46.56%。Al-farsi等[2]研究发现,盐分胁迫下紫花苜蓿通过增加叶片游离脯氨酸量、可溶性总酚量、过氧化氢酶活性和叶片钾量来提高牧草鲜草产量。赵霞等[6]研究发现,长期盐分胁迫下,苜蓿根、茎、叶中的Na+量增多,K+量减少,光合效率减弱,叶绿素和类胡萝卜素量降低。【切入点】紫花苜蓿是一种高产高耗水类牧草,生育期耗水量变化较大(130~170 mm),其中拔节期和分枝期是苜蓿需水量的关键时期[7-9]。Liu等[10]研究发现,在供水有限的条件下,与其他单期灌溉相比,分枝期亏缺灌溉对牧草产量的影响更大。成自勇[11]也认为,苜蓿分枝期对水分胁迫最敏感,该时期干旱胁迫对产量的影响最显著。【拟解决的关键问题】然而,也有学者认为土壤盐分可能会增强其抗旱能力,进而缓解水分胁迫对产量的影响[12-13]。翁亚伟等[13]研究发现,盐分和干旱复合胁迫可以通过增强根系水分吸收并降低根、叶中的ABA量以维持较高的光合能力,进而提高小麦的干旱适应性。目前,关于分枝期紫花苜蓿生长对水盐联合胁迫的响应机理研究较少。因此,本研究通过2 a的盆栽试验(2018—2019年),分析水盐胁迫对苜蓿植株生长指标(株高、地上生物量、根系生物量)、脯氨酸量、Na+量、SOD活性及POD活性的影响;建立水盐胁迫与苜蓿产量、渗透调节物质及抗氧化酶的关系,揭示水盐胁迫下苜蓿植株地上渗透调节物质及抗氧化酶活性对苜蓿生长的调节机制。
1 材料与方法
1.1 试验设计
试验于2018—2019年在内蒙古磴口县圣牧高科经济园区进行。供试苜蓿品种为紫花苜蓿“阿尔冈金”。盆栽试验用土壤的值为0.23 dS/m,pH值为7.5,全氮质量分数为0.19 g/kg,有机碳质量分数为2.35 g/kg。植物在塞式托盘中发芽,当幼苗长出3片叶子时,移栽到花盆中。每个花盆(直径24 cm,高18 cm)中装满7.5 kg风干土,充分灌溉后,每个花盆定植20株,每个处理设置4个重复,盆栽试验采用完全随机区组设计,共设置48盆。每个容器外部均用厚度为20 mm的保温棉包裹,防止外界温度对容器内部水分蒸发的影响。整个生长期采用灌溉器进行灌溉,容器内接有PVC软管的滴灌器(直径1 cm),软管出水口采用迷宫长流道剑柄结构滴箭进行灌溉,可有效减压并避免水冲刷土壤和基质。
盆栽试验设置3个水分处理,分别为:W1处理(土壤含水率为田间持水率的55%~70%);W2处理(土壤含水率为田间持水率的70%~85%),W3处理(土壤含水率为田间持水率的85%~100%)。根据当地土壤类型的实际情况,本试验所用土壤为盐渍土,可溶性盐主要成分为氯化钠(NaCl)和硫酸钠(Na2SO4)(图1)。因此,试验中盐分处理配制了NaCl和Na2SO4的混合盐(NaCl∶Na2SO4=1∶1),分别设置3个盐分处理,混合盐分添加量分别为干土质量的0.2%(S1:1.5 g/kg)、0.4%(S2:3.0 g/kg)、0.6%(S3:4.5 g/kg),对照(S0:<1.0 g/kg),灌溉时混合盐分随水施入。水盐胁迫处理前均采用常规灌溉,使土壤含水率达到田间持水率的85%以上。苜蓿进入分枝期进行水盐胁迫处理,盐分处理2 d后进行水分控制。W3处理含水率保持在85%f~100%f,含水率低于85%f时进行灌水。低水处理和中水处理的土壤含水率保持在70%f~85%f和55%f~70%f,当W2处理含水率低于70%时灌水,当W1处理含水率低于55%时灌水。用烘干法测定土壤初始含水率,控水期间用称质量法监测各处理土壤含水率,保持土壤相对水分量变化幅度在5%内,所有盆栽用自来水灌溉。待苜蓿初花期时在每个处理选择10株长势一致的植株测定苜蓿生长和生化指标。

图1 不同水盐处理下的盆栽试验布置方案
1.2 试验测定指标及方法
苜蓿地上及地下生物量测定:苜蓿初花期时,用刻度尺从底部到顶端测量植株高度(地面以上为株高),然后将整株植物取出,洗净后称量地上部分与地下部分的鲜质量。将一部分鲜草样品放入105 ℃的烘箱中杀青10 min后,于80 ℃烘干至恒质量,分别称量地上部分与地下部分干质量。苜蓿幼苗根茎比=地下干质量/地上干质量。另一部分鲜草样品放入自封袋用液氮速冻。
生化指标测定:称取植株鲜样,每个处理设置3个重复,分别测定脯氨酸量、Na+量、SOD活性和POD活性。脯氨酸量采用酸性茚三酮法测定,Na+量用火焰光度法测定,超氧化物歧化酶(SOD)活性采用氮蓝四唑法测定,过氧化物酶(POD)活性采用愈创木酚显色法测定。各指标具体测定方法参照王宝山[4]的研究。
苜蓿耗水量计算式为:
=Δ++,(1)
式中:为苜蓿耗水量(mm);Δ为盆栽中苜蓿土壤含水率变化量(mm);为灌溉量(mm);为降水量(mm);由于试验期间无降水,忽略降水量。
抗逆指数(s):干旱胁迫强度()是评价抗旱鉴定试验是否达到胁迫环境条件的指标;盐对作物生长的影响可以通过敏感性指数(S)来评价[14]。综合2个指标的计算方法,本研究中水盐胁迫对苜蓿生长的影响通过抗逆指数(s)来评价。s的估计值范围为0~1,s值越大说明胁迫越严重。计算式为:
s1-DW/con, (2)
式中:DW为水盐处理平均生物量(地上生物量、根系生物量)测定值(g);con为对照(无盐分处理,正常灌溉处理)平均生物量(地上生物量、根系生物量)测定值(g)。
1.3 数据分析
采用Microsoft Excel 2010软件和Origin 2018软件对试验数据进行处理并进行图表绘制,利用SPSS 17.0对数据进行统计分析并进行相关分析,采用邓肯新复极差检验法进行多重比较,检验处理间的差异显著性。
2 结果与分析
2.1 水盐胁迫对紫花苜蓿生长的影响
水盐胁迫对地上生物量、根系生物量及根茎比均有显著影响(<0.05),而对株高无显著影响(表1,表2)。相同灌溉水平下,株高随盐分增加呈先增加后降低的变化趋势,S1处理平均株高相比S0处理高1.38 cm,而盐分S3处理平均株高相比S0处理低2.23 cm。这说明低盐分胁迫促进植株生长,但高盐分抑制了苜蓿生长。相同盐分水平下,2 a苜蓿株高、地上生物量和根系生物量均随灌水量的增加而增加,但增产效应随灌水量的增加而下降。相同灌溉水平下,2 a平均地上生物量和根系生物量均随盐分的增加而减小,相较于无盐S0处理,S1—S3处理平均地上生物量依次平均降低11.59%、15.78%、29.82%。但在W1、W2灌溉水平下,S1处理和S2处理的地上生物量高于S0处理,在W1灌溉水平下平均增加4.30%和2.17%,在W2灌溉水平下增加6.55%和5.09%。相对于地上生物量,同一灌水条件下S1处理和S2处理对根系生物量影响程度较小,且平均根系生物量无显著差异(>0.05)。2 a苜蓿平均根茎比随灌水量的增加而减小,随盐度的增加先减小后增大,其中S3处理平均根茎比较S0处理平均提高1.78%。
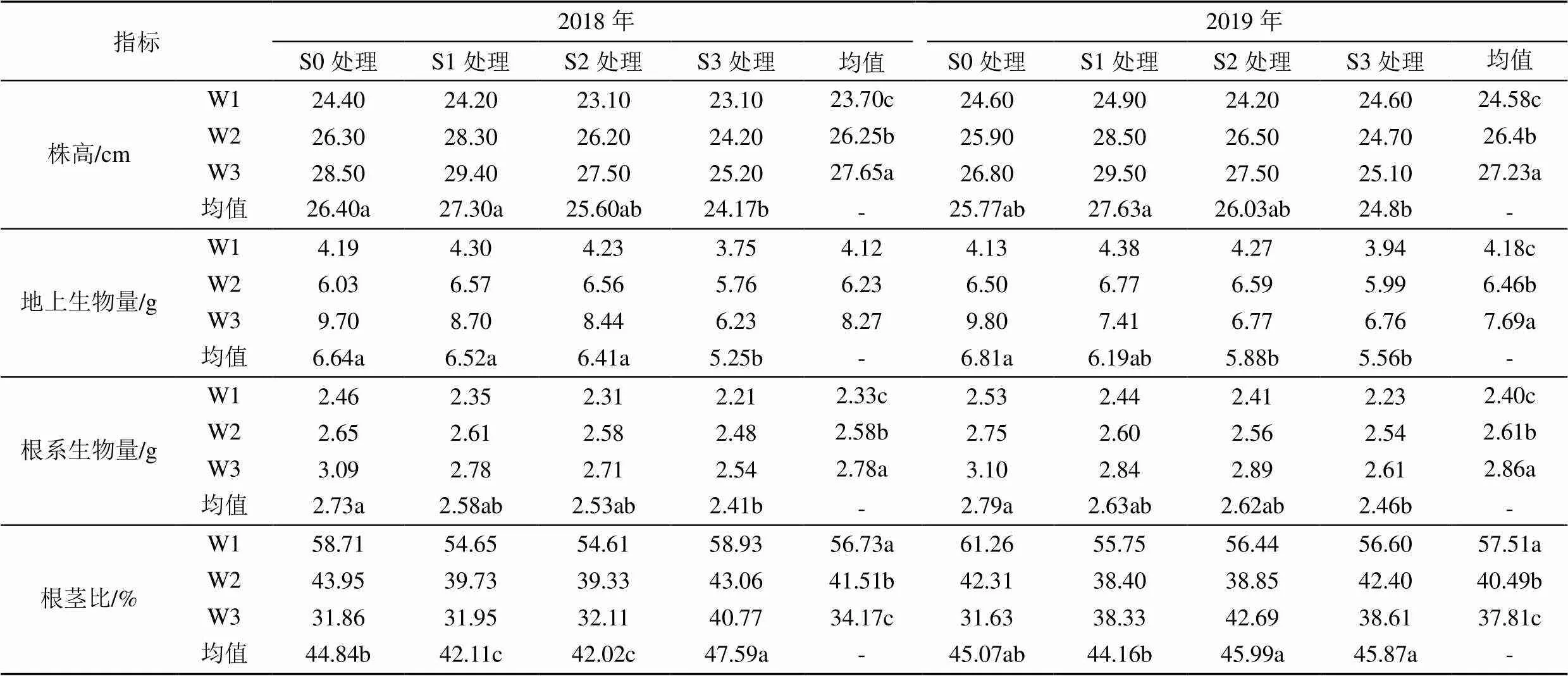
表1 不同水盐处理下苜蓿生物量
注 不同小写字母表示在0.05水平上有显著差异。*、**和***分别表示<0.05、<0.01和<0.001。ns表示无显著差异。

表2 不同水盐处理下苜蓿生物量双因素方差分析结果
水盐胁迫下苜蓿地上与根系生物量s值如图2所示(图中不同小写字母表示在0.05水平上有显著差异)。2 a水盐胁迫下苜蓿地上生物量的s值整体高于根系生物量,平均增加0.19和0.23。相同灌溉水平下,S3处理地上生物量s值显著高于S1处理和S2处理,平均分别增加0.16和0.14,而S1处理和S2处理地上生物量s值无显著差异(>0.05)。相同盐分处理下,W3处理和W2处理地上生物量s较W1处理平均分别降低0.23和0.33,根系生物量s值较W1处理平均分别降低了0.08和0.13。综上,苜蓿茎叶对水盐胁迫的敏感性高于根系,灌水量增加至W2处理及以上时,植株对盐分敏感性减弱。

图2 2018—2019年不同水盐胁迫下苜蓿地上部和根系的抗逆性
2.2 水盐胁迫对紫花苜蓿渗透调节物质的影响
2019年水盐胁迫下苜蓿生化特征(脯氨酸量、Na+量、SOD活性及POD活性)如图3所示。低水处理下,与S0处理相比,低水复合盐分胁迫显著增加了植株脯氨酸量和Na+量,S1—S3处理脯氨酸量依次分别提高30.63%、74.28%、107.15%;Na+量分别提高3.87%、17.28%、64.72%。S3盐分水平下,灌水量增加降低了脯氨酸量和Na+量,相比W1处理,W2处理脯氨酸量、Na+量分别降低49.32%、32.34%;W3处理分别降低57.81%、49.54%。同一水分胁迫下,SOD活性和POD活性随盐分增加呈先增加后减小,其中S1W1处理SOD活性和S2W1处理POD活性均显著高于其他处理(<0.05);相比S0处理,S1处理植株SOD活性增加22.65%,但S2处理和S3处理分别减少9.87%和54.47%。S1—S3处理POD活性较S0处理依次分别提高18.91%、34.60%、5.43%。相同盐分水平下,植株SOD、POD活性呈增加趋势,但各处理SOD活性均显著低于S0W1处理(<0.05),平均降低50.34%;S3W2处理和S3W3处理POD活性相比S0W1处理分别提高18.85%和37.13%。

图3 2019年不同水盐胁迫下苜蓿生化特征
2.3 水盐胁迫下苜蓿耗水量、生长与渗透调节物质的响应关系
2018年和2019年水盐胁迫下苜蓿分枝期—初花期土壤贮水变化量和耗水量如图4所示。除W1灌溉水平,同一灌溉水平下苜蓿耗水量随着盐分的增加呈减小趋势,S1—S3处理耗水量较S0处理平均分别降低3.51%、2.50%和14.07%。当盐分超过3 g/kg,土壤贮水变化量显著增加,苜蓿耗水量显著降低,S3处理平均耗水量较S0—S2处理分别降低13.71%、10.05%和13.23%。低水处理下,适宜盐分胁迫有利于提高苜蓿耗水量,其中S2W1处理耗水量相比S0W1、S1W1处理和S3W1处理分别平均增加6.48%、13.68%和31.39%。

图4 2018—2019年不同处理紫花苜蓿分枝期—初花期土壤贮水量和耗水量变化
由图5可知,相比单一水分胁迫,适宜盐分(<3 g/kg)条件下苜蓿通过增加植株中脯氨酸量、Na+量、SOD活性及POD活性提高了苜蓿耗水量和生物量。当盐分高于3 g/kg时,尽管植株增加了脯氨酸量、Na+量和POD活性,降低了SOD活性,但地上生物量和耗水量均明显下降。增加灌水量有利于缓解盐分胁迫,促进根系对水分吸收,提高耗水量和地上生物量,相对于单一干旱胁迫,地上生物量的s降低0.50。
水盐胁迫下苜蓿脯氨酸量、Na+量、SOD活性、POD活性、耗水量及地上生物量的相关分析如图6所示。水盐胁迫下紫花苜蓿地上脯氨酸量、Na+量与耗水量和地上生物量指数函数关系,相关性较好(2>0.5)。脯氨酸量与Na+量显著正相关(2=0.701);地上生物量与耗水量呈正相关,相关性较好(2=0.797)。然而,SOD活性和POD活性与耗水量的相关性均较弱,不具有统计学意义。

注 数据为变量的相对变化,参考植物在单一干旱胁迫下的生长(S0W1,绿线),y轴是相对刻度。

图6 苜蓿植株脯氨酸量、Na+量、SOD活性及POD活性与耗水量及地上生物量的响应关系
3 讨 论
紫花苜蓿是一种中等耐盐作物,其耐盐阈值为2.0 dS/m[4]。当土壤盐分超过3 g/kg时,盐分胁迫增加导致紫花苜蓿叶片水势、光合速率、蒸腾速率、气孔导度降低[12-13]。土壤水分消耗减少,土壤剩余水量增加,土壤中贮水变化量显著高于其他盐分处理,其原因是高盐浓度下土壤Na+量的增加导致土壤颗粒收缩,胶体颗粒分离和膨胀,土壤体积质量增大,降低土壤孔隙率和饱和导水率[15],作物根区盐分过高会造成渗透胁迫,降低根系对水分的吸收,作物蒸腾显著降低,使得土壤中剩余水量增加[16-17]。刘莎莎等[18]研究表明,土壤中适量浓度Na+盐离子有利于植物通过吸收盐离子来降低渗透势,在消耗较少能量的基础上提高根系吸水能力。适量的盐分和亏水灌溉对苜蓿水分利用会产生正向效应,S2W1处理耗水量相比S0W1处理提高6.48%,且S2W1处理耗水量均高于S1W1处理,可能是由于添加NaCl导致植株吸收和积累Na+量增加,降低了组织渗透势,维持了细胞膨压,从而缓解了干旱胁迫对叶片光合机构的损伤,阻止光合速率的下降,蒸腾持续时间增长,耗水量提高[12]。根系在盐胁迫下根系总长、表面积和体积等参数显著降低,但也有研究[6,16]发现盐胁迫促进侧根发育。本研究表明,2 a苜蓿地上生物量的s整体高于根系生物量,苜蓿茎叶对水盐胁迫的敏感性高于根系,与前人[16]研究结果相同。
盐分与干旱双重胁迫时作物表现出了适应性[4,12-13]。在干旱条件下,植物对Na+的吸收和积累显著增加,并在一定程度上降低了叶片的渗透势和水势,增强从外界介质吸水的动力,以维持细胞膨压和植株的代谢、生长[17]。因此,土壤中适量盐分的存在有利于植株积累更多Na+量参与渗透调节,以改善植株的水分状况和光合性能,保持植株较高的生长速率,减轻干旱对植物的负效应[18]。陈春晓等[17]研究表明,轻度干旱和盐分胁迫下,沙枣幼苗在一定程度上通过减少光捕获、热耗散和酶活性调节稳定光合机构功能,阻止光合速率的下降,造成沙枣幼苗除胞间CO2浓度外的各光合参数及荧光参数其他参数呈降低趋势。重度盐旱胁迫下光系统Ⅱ和抗氧化酶系统损伤,光合速率下降更加显著,导致地上生物量降低。本研究发现在盐分和干旱共胁迫时,适宜的盐分浓度下,植物会表现出比在单一胁迫下生长更好的现象,其中盐分S1处理中株高、地上生物量和根系生物量相比对照小幅度增加,而盐分超过3 g/kg显著降低了苜蓿根系和地上生物量,该结果与前人[13]研究结果相似。相比单一水分胁迫(S0W1处理),增加盐分提高了植株POD活性,而盐分超过3 g/kg时,植株抗氧化酶活性降低,导致活性氧造成的伤害增强,增加灌溉量起到了缓解作用。由于本研究盆栽试验是在苜蓿分枝期进行水盐互交处理,该时期苜蓿对盐分和水分胁迫都具备一定的耐受能力。然而,土壤中盐分超过苜蓿耐盐阈值后,尽管水盐胁迫下苜蓿植株增加脯氨酸量、Na+量及SOD活性,但不足以避免显著的氧化损伤或过氧化物积累,造成耗水量和生物量下降。分枝期紫花苜蓿抗旱性对土壤盐分响应的适宜范围还需进一步研究。
4 结 论
与单一水分、盐分胁迫相比,水盐胁迫导致苜蓿耗水量和地上生物量进一步降低。土壤盐分低于3 g/kg提高了苜蓿株高,但轻微降低了地上部生物量。盐分超过3 g/kg时,苜蓿生长产生抑制作用,显著降低了紫花苜蓿的生物量,增加了土壤水分,导致苜蓿耗水量降低。苜蓿分枝期在干旱和盐分复合胁迫条件下表现出一定的交叉适应现象。与单一水分胁迫相比,添加适量盐分(≤3 g/kg)可以更好地提高苜蓿脯氨酸量和Na+量、SOD活性和POD活性,提高苜蓿生物量和耗水量。土壤全盐量在2~3 g/kg之间时适宜苜蓿生长,灌水量为田间持水率的70%~85%最佳。
[1] ASHRAF M, FOOLAD M R. Pre‐sowing seed treatment—a shotgun approach to improve germination, plant growth, and crop yield under saline and non‐saline conditions[J]. Advances in Agronomy, 2005, 88: 223-271.
[2] AL-FARSI S M, AL-SADI A M, ULLAH A, et al. Salt tolerance in alfalfa landraces of Omani origin: Morpho-biochemical, mineral, and genetic diversity assessment[J]. Journal of Soil Science and Plant Nutrition, 2021, 21(2): 1 484-1 499.
[3] SINGH S, GUPTA A K, KAUR N. Differential responses of antioxidative defence system to long‐term field drought in wheat (L.) genotypes differing in drought tolerance[J]. Journal of Agronomy and Crop Science, 2012, 198(3): 185-195.
[4] 王宝山. 逆境植物生物学[M]. 北京: 高等教育出版社, 2010.
WANG Baoshan. Plant biology under stress[M]. Beijing: Higher Education Press, 2010.
[5] 柳燕兰, 郭贤仕, 马明生, 等. 干旱复水对春玉米幼苗生长和生理特性的影响及其根源ABA调控效应[J]. 干旱地区农业研究, 2019, 37(1): 187-193, 199.
LIU Yanlan, GUO Xianshi, MA Mingsheng, et al. Effects of drought and rewatering on growth and physiological characteristics of maize seedlings and regulation of root-sourced ABA[J]. Agricultural Research in the Arid Areas, 2019, 37(1): 187-193, 199.
[6] 赵霞, 叶林. 盐碱胁迫对紫花苜蓿生长、品质及光合特性的影响[J]. 江苏农业科学, 2017, 45(21): 176-180.
ZHAO Xia, YE Lin. Effects of saline-alkali stress on growth, quality and photosynthetic characteristics of alfalfa[J]. Jiangsu Agricultural Sciences, 2017, 45(21): 176-180.
[7] 孙洪仁, 刘国荣, 张英俊, 等. 紫花苜蓿的需水量、耗水量、需水强度、耗水强度和水分利用效率研究[J]. 草业科学, 2005, 22(12): 24-30.
SUN Hongren, LIU Guorong, ZHANG Yingjun, et al. Water requirement, water consumption, water requirement rate, water consumption rate and water use efficiency of alfalfa[J]. Pratacultural Science, 2005, 22(12): 24-30.
[8] ALBACETE A A, MARTÍNEZ-ANDÚJAR C, PÉREZ-ALFOCEA F. Hormonal and metabolic regulation of source–sink relations under salinity and drought: From plant survival to crop yield stability[J]. Biotechnology Advances, 2014, 32(1): 12-30.
[9] CAO X S, FENG Y Y, LI H P, et al. Effects of subsurface drip irrigation on water consumption and yields of alfalfa under different water and fertilizer conditions[J]. Journal of Sensors, 2021, 2021: 1-12.
[10] LIU M G, WANG Z K, MU L, et al. Effect of regulated deficit irrigation on alfalfa performance under two irrigation systems in the inland arid area of Midwestern China[J]. Agricultural Water Management, 2021, 248: 106 764.
[11] 成自勇. 甘肃秦王川灌区苜蓿草地土壤水盐动态及其生态灌溉调控模式研究[D]. 兰州: 甘肃农业大学, 2005.
CHENG Ziyong. Research on the dynamics of soil water and salt of purple medic grassland and its ecological optimal irrigation control in Qinwangchuan irrigated regions in Gansu[D]. Lanzhou: Gansu Agricultural University, 2005.
[12] 解卫海, 马淑杰, 祁琳, 等. Na+吸收对干旱导致的棉花叶片光合系统损伤的缓解作用[J]. 生态学报, 2015, 35(19): 6 549-6 556.
XIE Weihai, MA Shujie, QI Lin, et al. The mitigating effects of Na+accumulation on the drought-induced damage to photosynthetic apparatus in cotton seedlings[J]. Acta Ecologica Sinica, 2015, 35(19): 6 549-6 556.
[13] 翁亚伟, 张磊, 张姗, 等. 盐旱复合胁迫对小麦幼苗生长和水分吸收的影响[J]. 生态学报, 2017, 37(7): 2 244-2 252.
WENG Yawei, ZHANG Lei, ZHANG Shan, et al. Effects of salt with drought stress on growth and water uptake of wheat seedlings[J]. Acta Ecologica Sinica, 2017, 37(7): 2 244-2 252.
[14] REJILI M, VADEL A M, GUETET A, et al. Effect of NaCl on the growth and the ionic balance K+/Na+of two populations of(L.) (Papilionaceae)[J]. South African Journal of Botany, 2007, 73(4): 623-631.
[15] YUAN C F, FENG S Y, WANG J, et al. Effects of irrigation water salinity on soil salt content distribution, soil physical properties and water use efficiency of maize for seed production in arid Northwest China[J]. International Journal of Agricultural and Biological Engineering, 2018, 11(3): 137-145.
[16] 高玉红, 闫生辉, 邓黎黎. 不同盐胁迫对甜瓜幼苗根系和地上部生长发育的影响[J]. 江苏农业科学, 2019, 47(3): 120-123.
GAO Yuhong, YAN Shenghui, DENG Lili. Effects of salts stress on growth of roots and shoots of melon seedlings[J]. Jiangsu Agricultural Sciences, 2019, 47(3): 120-123.
[17] 陈春晓, 谢秀华, 王宇鹏, 等.盐分和干旱对沙枣幼苗生理特性的影响[J]. 生态学报, 2019, 39(12): 4 540-4 550.
CHEN Chunxiao, XIE Xiuhua, WANG Yupeng, et al. Effects of salt and drought on the physiological characteristics of elaeagnus angustifolia L. seedlings[J]. Acta Ecologica Sinica, 2019, 39(12): 4 540-4 550.
[18] 刘莎莎, 柏新富, 冯春晓, 等. 干旱条件下土壤盐分对大豆生长及光合作用的影响[J]. 大豆科学, 2017, 36(6): 921-926.
LIU Shasha, BAI Xinfu, FENG Chunxiao, et al. Effects of soil salinity on the growth and photosynthesis of soybean under drought conditions[J]. Soybean Science, 2017, 36(6): 921-926.
Combined Effect of Water and Salt Stress on Growth and Physiological Traits of Alfalfa at Branching Stage
Tian Delong1, Hou Chenli2, Xu Bing1, Ren Jie1, Zhang Chen1
(1. Institute of Water Resources for Pastoral Area, Ministry of Water Resources, Huhhot 010020, China; 2.School of Energy and Environment, Inner Mongolia University of Science and Technology, Baotou 014010, China)
【Objective】Alfalfa is sensitive to water stress at branching stage, but how a simultaneous salinity stress affects this sensitivity is not well understood. The objective of this paper is to address this issue.【Method】Pot experiments were conducted in 2018—2019 at the Shengmu High-tech Economic Park, in Dengkou County, Inner Mongolia. It consisted of three water treatments with the soil water content kept at 55%~70% (W1), 75%~85% (W2), and 85%~100% (W3) of the field capacity, respectively. Each water treatment had four salinity stresses with the soil salt content set at 1.0 (S0), 1.5 (S1), 3.0 (S2) and 4.5(S3) g/kg, respectively. In each treatment, we measured osmotic substances, antioxidant enzymatic activities, above- and below-ground biomass at early flowering stage. 【Result】Water and salinity stresses reduced water consumption and aboveground biomass more considerably when they worked collectively than working separately. When the water stress was minor, however, imposing a slight salinity stress with soil salt content ≤3 g/kg can increase the contents of proline and Na+, boosted peroxidase (POD) activity, and reduced superoxide dismutase (SOD). This alleviated oxidative damage of the water stress, thereby promoting crop growth. Compared with S0+W1, S2+W1 increased the contents of proline and Na+, and boosted POD activity, by 74.28%, 17.28% and 34.60%, respectively, while reducing SOD activity by 9.87%. These led to an increase in water consumption and aboveground biomass by 6.48% and 2.17%, respectively.【Conclusion】When water stress is not severe, imposing a slight salinity stress can improve the tolerance of alfalfa to water and salinity stress at branching stage. For all treatments we studied, keeping soil salt content in the range of 2~3 g/kg and soil water content in the range of 70%~85% of the field water capacity was optimal to improve resistance of the crop to both salinity and water stresses.
water salt stress; alfalfa; biomass; osmotic regulating substance; antioxidant enzyme activity
1672 - 3317(2023)04 - 0008 - 07
S27
A
10.13522/j.cnki.ggps.2022403
田德龙, 侯晨丽, 徐冰, 等. 分枝期水盐胁迫对紫花苜蓿生长及生理特征的影响[J]. 灌溉排水学报, 2023, 42(4): 8-14.
TIAN Delong, HOU Chenli, XU Bing, et al. Combined Effect of Water and Salt Stress on Growth and Physiological Traits of Alfalfa at Branching Stage[J]. Journal of Irrigation and Drainage, 2023, 42(4): 8-14.
2022-07-20
内蒙古自治区科技计划项目(2019GG022-03)
田德龙(1982-),男。高级工程师,主要从事节水灌溉理论与农田生态环境研究。E-mail: mkstdl@126.com
责任编辑:韩 洋
