Development and characterization of a novel common wheat–Mexico Rye T1DL·1RS translocation line with stripe rust and powdery mildew resistance
2023-05-08LlJiaojiaoZHAOLiBoyaFUYuZHANGShufaLlUShuhuiYANGQunhuiWUJunLlJiachuangCHENXinhong
Ll Jiao-jiao, ZHAO Li, LÜ Bo-ya, FU Yu, ZHANG Shu-fa, LlU Shu-hui, YANG Qun-hui, WU Jun, Ll Jiachuang, CHEN Xin-hong
Shaanxi Key Laboratory of Plant Genetic Engineering Breeding, College of Agronomy, Northwest A&F University, Yangling 712100, P.R.China
Abstract Rye (Secale cereale L., 2n=2x=14, RR) is a significant genetic resource for improving common wheat because of its resistance to multiple diseases and abiotic-stress tolerant traits.The 1RS chromosome from the German cultivated rye variety Petkus is critical in wheat breeding.However, its weakened disease resistance highlights the need to identify new resources.In the present study, a novel derived line called D27 was developed from common wheat and Mexico Rye.Cytological observations characterized the karyotype of D27 as 2n=42=21 II.Genomic in situ hybridization indicated that a pair of whole-arm translocated Mexico Rye chromosomes were inherited typically in the mitotic and meiosis stages of D27.Experiments using fluorescence in situ hybridization (FISH) and gliadin electrophoresis showed that D27 lacked wheat 1DS chromosomes.They were replaced by 1RS chromosomes of Mexico Rye, supported by wheat simplesequence repeat markers, rye sequence characterized amplified region markers, and wheat 40K SNP array analysis.The wheat 1DS chromosomes could not be detected by molecular markers and wheat SNP array, but the presence of rye 1RS chromosomes was confirmed.Agronomic trait assessments indicated that D27 had a higher tiller number and enhanced stripe rust and powdery mildew resistance.In addition, dough properties analysis showed that replacing 1DS led to higher viscosity and lower dough elasticity in D27, which was beneficial for cake making.In conclusion, the novel cytogenetically stable common wheat–Mexico Rye T1DL·1RS translocation line D27 offers excellent potential as outstanding germplasm in wheat breeding programs focusing on disease resistance and yield improvement.Additionally, it can be valuable for researching the rye 1RS chromosome’s genetic diversity.
Keywords: common wheat, disease resistance, dough properties, rye, translocation line
1.lntroduction
Common wheat (TriticumaestivumL., 2n=42, AABBDD) is the world’s most important crop that is affected by increasingly severe diseases (Zahooret al.2020).China’s two main wheat diseases are stripe rust caused byPucciniastriiformisf.sp.tritici(Pst) and powdery mildew caused byBlumeriagraminis(DC.) E.O.f.sp.tritici(Bgt), both harmful to wheat in all its growth stages (Kumaret al.2019).Stripe rust and powdery mildew are airborne fungal diseases that detrimentally affect wheat yield and quality, characterized by widespread occurrence, rapid spread, good adaptability, and high infectiousness.Since the 1980s, Chinese wheat breeders have developed a series of improved disease-resistant varieties, including Xiaoyan 6, Jimai 22, and Zhoumai 16 (Li 2010).However, the increasingly smaller genetic diversity of wheat varieties and variations in disease races or isolates have led to the accelerated recession of resistance and increased the severity of disease because of long-term breeding with single parents (Wang Y Zet al.2020).Therefore, it is necessary to expand the range of available breeding resources and develop novel multi-diseaseresistant wheat materials to relieve these problems.
Many studies have confirmed that numerous excellent genes found in species related to wheat can be used to improve the resistance to diseases and yield in wheat.These genes are usually transferred into the genetic background of wheat by distant hybridization and using alien chromosomes as carriers (Liuet al.2014).Wheatalien addition, substitution, and translocation lines are the three most common types of hybrid derivatives, and their stability is theoretically greater in the same order (Liet al.2019).Therefore, wheat-alien translocation lines with outstanding agronomic traits are valuable for wheat improvement.Examples include a wheat–HaynaldiavillosaT6AL·6VS translocation line, a wheat–PsathyrostachyshuashanicaT5D·5Ns translocation line with wheat powdery mildew resistance (Chenet al.1995; Li J Cet al.2020), a wheat–Aegilopsspeltoidestranslocation line with enhanced resistance to stem rust (Fariset al.2008), and a wheat–ThinopyrumelongatumT7DL·7EL translocation line with Fusarium head blight resistance (Wang H Wet al.2020).These desirable traits are difficult to obtain by traditional breeding, so the development of novel wheat-alien translocation lines was necessary.
Rye (SecalecerealeL., 2n=2x=14, RR) is a close relative of wheat.It is regarded as a key genetic resource for wheat improvement because it naturally possesses many genes that confer protection against various wheat diseases, including powdery mildew, rust (stripe, leaf, and stem), and sharp eyespot (Wanget al.2009).In addition, rye is characterized by a high dietary fiber content, lysine-rich kernels, long spikes, and strong stems (Rabinovich 1998).The first distant hybridization between rye and common wheat was conducted successfully by the British breeder Wilson in the 1870s (Rabinovich 1998).Subsequently, octaploid and hexaploidTriticalehybrids were created.OctaploidTriticale(2n=8x=56, AABBDDRR) had some disadvantages, such as low selffertility and unfilled grains, and thus it could not be used directly in production (Weimarck 1973; Ren and Lelley 1991).The hexaploidTriticale(2n=6x=42, AABBRR) produced by genomic substitution gradually became the focus of research because it combined the beneficial traits of both parents (Merker 1975; Rabinovich 1998).Several wheat–rye derived lines were developed by using octaploid or hexaploidTriticaleto selfing or crossing with wheat cultivars.Examples include the classical wheat–rye T1BL·1RS translocation line with superior resistance to wheat rust disease (Qiet al.2016), a wheat–rye 6R translocation line with resistance to powdery mildew and stripe rust (Li G Ret al.2020), and a wheat–rye T7BS·7RL translocation line with resistance to stripe rust and powdery mildew (Renet al.2020).Many wheat–rye T1BL·1RS translocation lines have now been developed, but T1DL·1RS translocation lines have rarely been reported in previous studies.
In this study, we developed a novel wheat–rye T1DL·1RS translocation line with superior resistance to stripe rust from the BC2F9progenies of common wheat cultivated variety (cv.) W770B×ryecv.Mexico Rye.The objectives of this study were: (1) to determine the chromosomal composition and duplication processes in the new translocation line using cytogenetic methods; (2) to clarify the homoeologous group of alien chromosomes using multiple molecular markers; and (3) to evaluate the agronomic performance and dough properties of this line in order to predict its possible utility.
2.Materials and methods
2.1.Plant materials
The plant materials comprised rye (S.cerealeL., 2n=2x=14, RR)cv.Mexico Rye, common wheatcv.W770B and Chinese Spring (CS), 1BL·1RS translocationcv.Shaan 229, hexaploidTriticale(2n=6x=42, AABBRR)cv.Lanxiaohei (LXH).Wheat–rye derived line D27 was developed from a cross between common wheatcv.W770B×ryecv.Mexico Rye, of which W770B was the female parent.The F1hybrid was recovered by embryo rescuing and chromosome doubling and then backcrossed to the wheat parent W770B twice.Until BC2F9, line D27 was screened for its morphological stability.CS, LXH, and Shaan 229 were used as control materials.The common wheat materials were provided by the Wheat Improvement Center in Yangling, China, and the wheat–Mexico Rye derived line was developed in our laboratory.All materials were deposited in the College of Agronomy, Northwest A&F University, China.The mature grains were tempered at 14% (w/w) moisture and ground into flour in a Brabender Quadrumat Senior mill.Then, sieved through a 200-mesh sieve and stored at 4°C for at least 1 wk for later use.
2.2.Cytological analysis
Sampling was conducted at appropriate plant growth stages.The root tips were cut when the roots reached a length of 1–2 cm at 25°C.Young panicles were sampled from the spikes when the distance between the flag leaf and the second leaf was 3–4 cm, or the spike length was 4–5 cm.Samples were treated with Carnoy’s fixative I for over 3 days before transferring to 70% ethanol and storing at –20°C.As a preliminary treatment, the apical meristems were rinsed three times with clear water and then digested in 2% cellulase plus 1% pectinase at 37°C for 60 min.Each root tip cell mixture was treated with 20 mL of acetic acid as a solvent.Anthers were stained with 1% aceto-carmine.Microscopic examinations were conducted using an Olympus microscope (BH2, Japan) at 400 times magnification.
2.3.In situ hybridization analyses
Slides with good split phases were treated with an ultraviolet light cross-linker at 1 200 mJ cm–2for 5 min to attach the chromosomes firmly on the slides from being washed away in the next steps and to improve probe binding rate duringinsituhybridization (Mccreery and Helentjaris 1994; Wang Y Zet al.2020).Genomicinsituhybridization (GISH) was conducted as described by Wetzel and Rayburn (2000), and rye genomic DNA was labeled with Dig-nick Translation Mix (Roche, Germany) for more than 1.5 h as a probe.The probe binding site was visualized using Anti-Digoxigenin-Fluorescein Fab fragments (Roche, German).In non-denaturing fluorescenceinsituhybridization (ND-FISH) (Fuet al.2015), Oligo-probe pSc119.2 (6-FAM-5´) and pTa535-1(TAMRA-5´) were synthesized in Sangon Company (Shanghai, China).Two Oligo-probes were used to identify the chromosomal composition of line D27.Details of the fluorescent karyotypes of the two probes were provided by Tanget al.(2014) and Fuet al.(2015).Each slide was hybridized with the Oligo-probes mixture (40 ng pSc119.2, 30 ng pTa535-1, and 10 mL 1×TE/2×SSC).Fluorescent signals were observed and captured using a microscope (Olympus BX60) with a Photometrics SenSys CCD camera.Slides with excellent observations by NDFISH were soaked in 70% alcohol under light for more than 24 h and used for sequential GISH analysis.
2.4.Analysis using sequence characterized amplified region (SCAR) markers and simple sequence repeat (SSR) molecular markers
To further determine the chromosomal composition of line D27, a total of 183 pairs of high-specificity SSR markers distributed on 21 pairs of wheat chromosomal arms were screened to analyze the variation of chromosome structure, according to Röderet al.(1998), Somerset al.(2004), and online markers tool (https://wheat-urgi.versailles.inrae.fr/).In addition, 31 pairs of rye genomespecific SCAR markers (Shimizuet al.1997; Chaiet al.2006) were selected to confirm the homoeology of the rye chromosomes introduced in line D27.The PCR products of the SCAR and SSR markers were electrophoresed on 8% polyacrylamide at a constant voltage of 170 V for 2 h and 1% agarose gels at a constant voltage of 120 V for 20 min.
2.5.Wheat 40K SNP suspension array analysis
Genomic DNA was extracted from line D27, and its parents were hybridized with a wheat 40K SNP array using the GenoBaits method by MOLBREEDING Biotechnology Company (Shijiazhuang, China).Single nucleotide polymorphism (SNP) loci in this array were distributed over 21 pairs of wheat chromosomes.Different bases (e.g., AA, TT, CC, GG, AT, and AG) indicated different genotypes in specific positions, and “NA” denoted the absence of this position in the chromosome.NA ratio=Number of SNP markers with NA genotype/Number of total SNP markers.SigmaPlot V12.5 (SYSTAT, USA) was used for data analysis, and MapChart V2.2 (Wageningen University, Netherlands) was used for chromosome mapping.
2.6.Analysis of gliadin subunits in line D27 and its parents
The rye 1RS chromosome carries thew-secalingene, thereby allowing the effective identification of the presence of the 1RS chromosome.Acid-polyacrylamide gel electrophoresis (A-PAGE) was applied to determine the composition of the gliadin subunits, including the secalin gene-specific subunit band from rye.A-PAGE was conducted as described by Zhaoet al.(2010) with slight modifications.The endosperm was cut from one seed each from line D27 and its parents to extract gliadin.The extraction mixture was kept in the dark for 12 h before centrifugation at 14 000 r min–1for 10 min, and the supernatant contained gliadin in the solution.Electrophoresis was conducted at a constant current of 80 V for 16 h.Coomassie brilliant blue was used to visualize the bands, and distilled water was used for decolorization.
2.7.Evaluation of morphologic traits and disease resistance
Six agronomic traits comprising the plant height, spike length, spikelets per spike, tiller number, kernels per spike, and thousand-kernel weight were assessed for line D27 and its parents in the field.The test materials were planted on the farm of Yangling Wheat Experimental Base and arranged separately in a completely randomized block design.Three rows were planted in each sample with a length of 1.5 m and a row spacing of 0.3 m apart at 6 grains per row.In two successive years, five plants were tested for each material with three replicates to ensure that accurate results were obtained.Moreover, we tested the resistance of line D27 and its parents to stripe rust and powdery mildew in a growth chamber at the seedling and adult stages.Five plants of each material were planted in one pot surrounded by susceptible control Chinese Spring and repeated three times.The virulent physiological races ofPstandBgtwere provided by the College of Plant Protection, Northwest A&F University, China.
Three virulent physiological races ofPst(CYR32, 33, 34) were used for inoculation, where the mixture ofPsturedospores and starch was prepared at a ratio of 1:50 by mass.In the seedling stage, the infection types (ITs) were recorded on the 15th day after shaking the powder mixture onto trifoliate stage plants.The reactions of the materials to stripe rust were scored according to Anet al.(2019) on a scale from 0 to 4, where 0, 0;, 1, 2, 3, and 4 denote immune, nearly immune, highly resistant, moderately resistant, moderately susceptible, and susceptible, respectively.The ITs were appended with “–” or “+” to denote lighter or heavier infections, respectively.In the adult stage, the reactions were recorded on the 21th day after smearing the powder mixture on the flag leaves.The responses to infection of materials were recorded as the percentage of thePstspores covered the total area of the leaves at the same position (secondary leaf) on each plant.
In the seedling stage, the powdery mildewBgtisolate E09 was used for inoculation, where the pathogen spores were dusted onto the leaves in the two-leaf stage.Cultivation was conducted at 22°C and 70% humidity in an incubator for 15 days before recording the reactions toBgt(Sheng 1988).In particular, ITs were assigned on a scale from 0 to 4, where 0, 0;, 1, 2, 3, and 4 denote immune, nearly immune, highly resistant, moderately resistant, moderately susceptible, and susceptible, respectively.In the adult stage, the responses to powdery mildew were scored based on the percentage of the total area of leaves in the same leaf position covered by spores (Li J Cet al.2020).The ITs were appended with “–” or “+” to denote lighter or heavier infections, respectively.
2.8.Analysis of starch gelatinization properties and dough viscoelastic properties
The gelatinization properties of material starch were performed by a rapid viscosity analyzer (RVA) (Perten, TechMastet, Sweden), referencing Gaoet al.(2021).A suspension of uniformly mixed samples and water was placed at an initial temperature of 50°C, heating to 95°C, held for 2.5 min, and cooling to 50°C.A thermal cycle for Windows Software was used to analyze the obtained pasting curve.The viscoelastic properties of the dough were analyzed by a rotational rheometer (DHR-1, USA), as previously described by Guoet al.(2021).The storage modulus (G´) and loss modulus (G´´) were measured with a 1% strain in the frequency range of 0.1 to 10 Hz at 25°C, and the loss tangent (tanδ)=G´´/G´ was calculated.Each sample was tested in triplicate.
3.Results
3.1.Cytogenetic observations D27
Root tip cells (RTCs) in the metaphase of mitosis were used to count the number of chromosomes.The RTCs from line D27 had a chromosomal number of 42 (Fig.1-A).Chromosomal pairing and division were observed in pollen mother cells (PMCs).In metaphase I of meiosis in the PMCs, line D27 had 21 bivalents without a univalent or other configuration (Fig.1-B; Table 1).In anaphase I of meiosis in line D27, the homoeologous chromosomes segregated in PMCs and moved to the cell poles in a normal manner (Fig.1-C).These results indicate that line D27 was a cytologically stable line with a normal number of chromosomes, pairing, and division.

Table 1 Chromosome numbers and pairing at meiotic phases of line D27
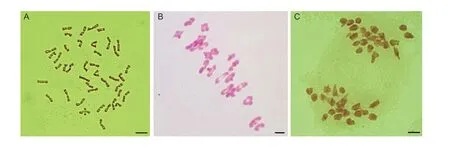
Fig.1 Microscopic observation of chromosomes in root tip cells and pollen mother cells.A, mitotic metaphase, chromosome morphology in line D27, 2n=42.B, meiotic metaphase I, bivalents in line D27, 2n=21 II.C, meiotic anaphase I, homologous chromosomes in D27, 2n=21+21.Scale bar, 10 mm.
3.2.GlSH analysis
GISH analysis was conducted using the whole genomic DNA from Mexico Rye as the probe and wheat parent W770B as the block to identify whether line D27 contained rye chromosomes.GISH based on the RTCs showed that line D27 contained two chromosome segments with yellow–green fluorescent signals, and the two alien chromosomes combined with two common wheat chromosomes to form two wheat–rye translocated chromosomes (Fig.2-A).In metaphase I of meiosis, a bivalent with a partial ring hybridization signal appeared on the equatorial plate in the PMCs (Fig.2-B).Subsequently, two chromosomes with partial hybridization signals moved to the cell poles in meiosis anaphase I (Fig.2-C).In the last stage of meiosis, each individual gamete expressed a fluorescent signal in the nucleus (Fig.2-D).Therefore, the GISH results obtained through RTCs showed that line D27 was a wheat–rye translocation line, and the results based on PMCs showed that the wheat–rye recombinant chromosome was genetically stable in line D27.
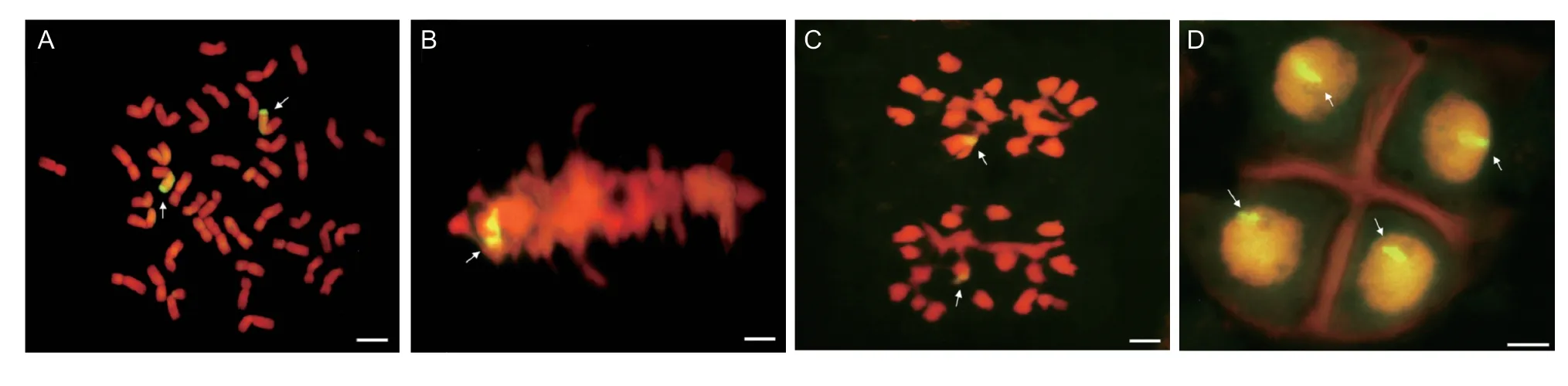
Fig.2 Genomic in situ hybridization (GISH) analysis of line D27.A, GISH analysis of chromosomes in root tip cells.Two chromosome segments were detected as alien chromosomes because they produced yellow-green fluorescent signals.B, GISH analysis in meiosis metaphase I.A ring bivalent with a partial fluorescent signal was observed when the chromosomes were on the equatorial plate.C, GISH analysis in meiosis anaphase I.Two chromosomes with partial hybridization signals moved to the poles.D, each progeny cell produced a fluorescent signal.GISH experiment using the total DNA from Mexico Rye as the probe and W770B as the block.Propidium iodide was used to counterstain chromosomes.Arrows indicate the alien chromosomes.Scale bar, 10 mm.
3.3.FlSH and sequential GlSH analysis
ND-FISH was conducted to determine the chromosomal composition of line D27 based on comparisons with the FISH karyotype of Chinese Spring wheat (Tanget al.2014).We found that line D27 produced the same FISH patterns as its wheat parent W770B, except that chromosome 1D was recombined with Mexico Rye 1RS and wheat 1DL (Fig.3-A, arrows).Sequential GISH was then performed to verify the presence of the rye chromosome (Fig.3-B), and the two chromosomes expressed fluorescent signals in their short arms.The breakage and combination sites were located near thecentromere according to the FISH karyotypes for rye and W770B (Fig.3-C and D), as shown in the diagram in Fig.3-E.Thus, line D27 had wheat–Mexico Rye 1DL·1RS translocated chromosomes.
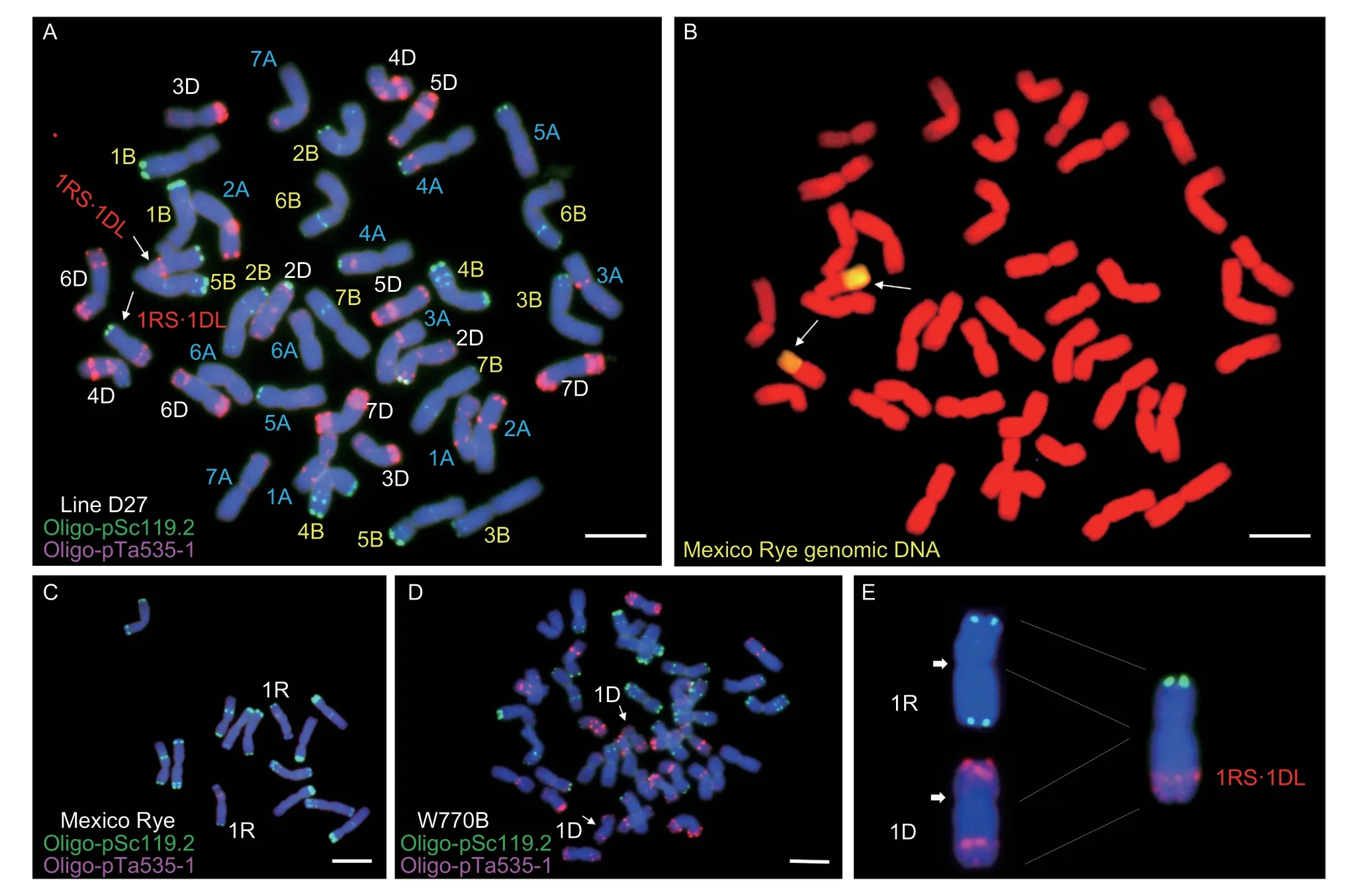
Fig.3 Fluorescence in situ hybridization (FISH) and sequential genomic in situ hybridization (GISH) analysis of line D27.A, FISH karyotype of D27.Arrows indicate the recombinant 1DL·1RS chromosome in a wheat background.B, sequential GISH for the same slide.Two chromosomal short arms with fluorescent signals from Mexico Rye are indicated by the arrows.C, FISH karyotype of Mexico Rye.D, FISH karyotype of common wheat W770B.Arrows indicate the normal 1D chromosome.E, the diagrammatic sketch of translocation in D27.Arrows indicate the breakage sites in the 1R and 1D chromosomes.In the FISH experiments, 4´,6-diamidino-2-phenylindole was used to counterstain chromosomes.Scale bar, 10 mm.
3.4.Analysis of molecular markers in line D27
We selected 31 pairs of R genome-specific SCAR markers to rapidly confirm the homoeology of translocated rye chromosomes in line D27.Four pairs of markers distributed in the homoeologous group 1 amplified 1RS chromosome-specific bands in Mexico Rye, LXH, and line D27 (Table 2; Fig.4, arrows).Therefore, the alien chromosomes introduced in line D27 might be the short arm of the rye 1RS chromosome.
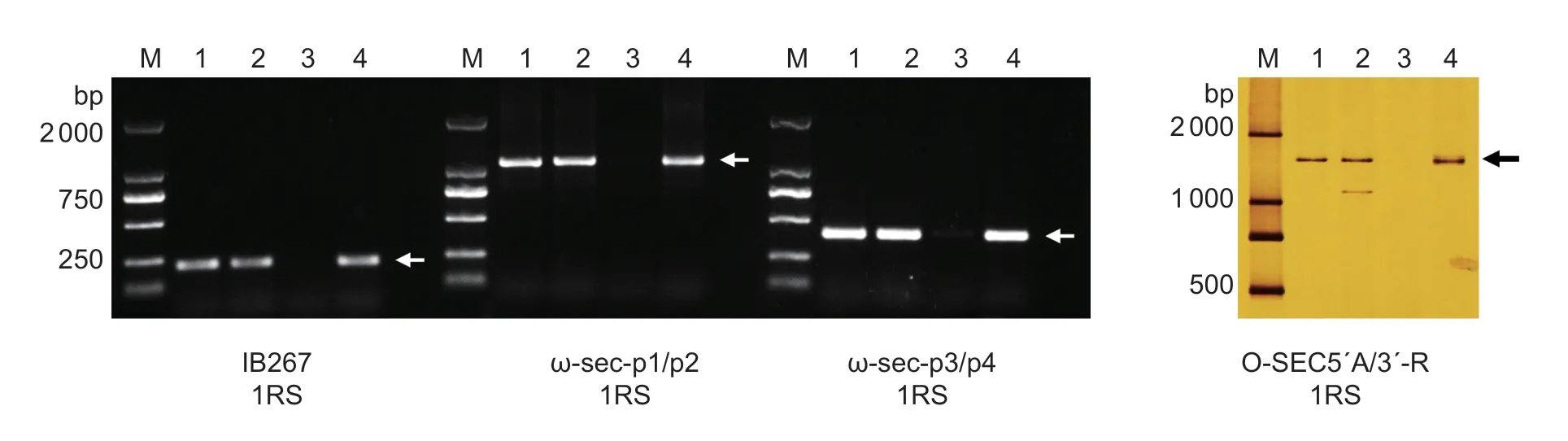
Fig.4 Analysis of alien chromosomes using sequence characterized amplified region (SCAR) markers.Four pairs of SCAR markers comprising IB267, ω-sec-p1/p2, ω-sec-p3/p4, and O-SEC5´-A/3´-R located on the 1RS chromosome amplified unique bands in Mexico Rye, Lanxiaohei (LXH), and line D27.Lane M, DL2000 marker; lane 1, Mexico Rye; lane 2, LXH; lane 3, W770B; lane 4, D27.Arrows indicate the 1RS chromosome-specific bands.
In addition, 183 pairs of SSR markers distributed in the chromosomal short and long arms of seven wheat homoeologous groups were used to identify translocated wheat chromosomes in line D27.Some markers amplified different bands in 1DS, but the markers in the other homoeologous groups amplified the same wheat chromosome-specific bands in line D27 and its wheat parent W770B.Four pairs of SSR markers in 1DS only amplified wheat 1DS chromosome-specific bands in W770B, but these bands were absent in LXH and line D27 (Table 2; Fig.5, arrows).Four pairs of SSR markers in 1DL amplified similar bands in W770B and line D27.These results indicate that line D27 lacked wheat 1DS chromosomes.

Fig.5 Analysis using simple sequence repeat (SSR) markers to genotype line D27.The SSR markers (Barc149, Xwmc432, Xgwm33, and Xgwm337) located on 1DS did not amplify wheat 1DS chromosome-specific bands in Mexico Rye and D27.However, the SSR markers (Xwmc429, Xwmc590, Xcfd63, and Xgwm232) located on 1DL amplified similar bands in W770B and D27.Lane M, DL2000 marker; lane 1, Mexico Rye; lane 2, W770B; lane 3, D27.Arrows indicate the missing 1DS chromosome-specific bands.
3.5.Comparison using 40K SNP array
In total, 40 478 valid SNP loci were tested in 21 wheat chromosomes.The genotype of each locus is shown in Appendix A.Line D27 is a translocation line, so some SNPs located on the translocated chromosome might have been lost.The NA ratio on each chromosome is presented in Table 3 and Fig.6-A.Wheat W770B had a low NA ratio (<10%) because of its intact wheat genome, but Mexico Rye had a high NA ratio (>70%) because itlacked the wheat genome.Derived line D27 had a similar NA ratio compared to its parent W770B, except for an unusual NA ratio in the 1D chromosome.In order to compare the genotypes in an objective manner, the SNPs on the 1D chromosome were arranged based on their physical positions, as shown in Fig.6-B.Clearly, line D27 had the same genotype as its alien parent rye in the short arm of the 1D chromosome but the same as its wheat parent W770B in the long arm.Therefore, the results confirmed that line D27 lacked wheat 1DS chromosome.
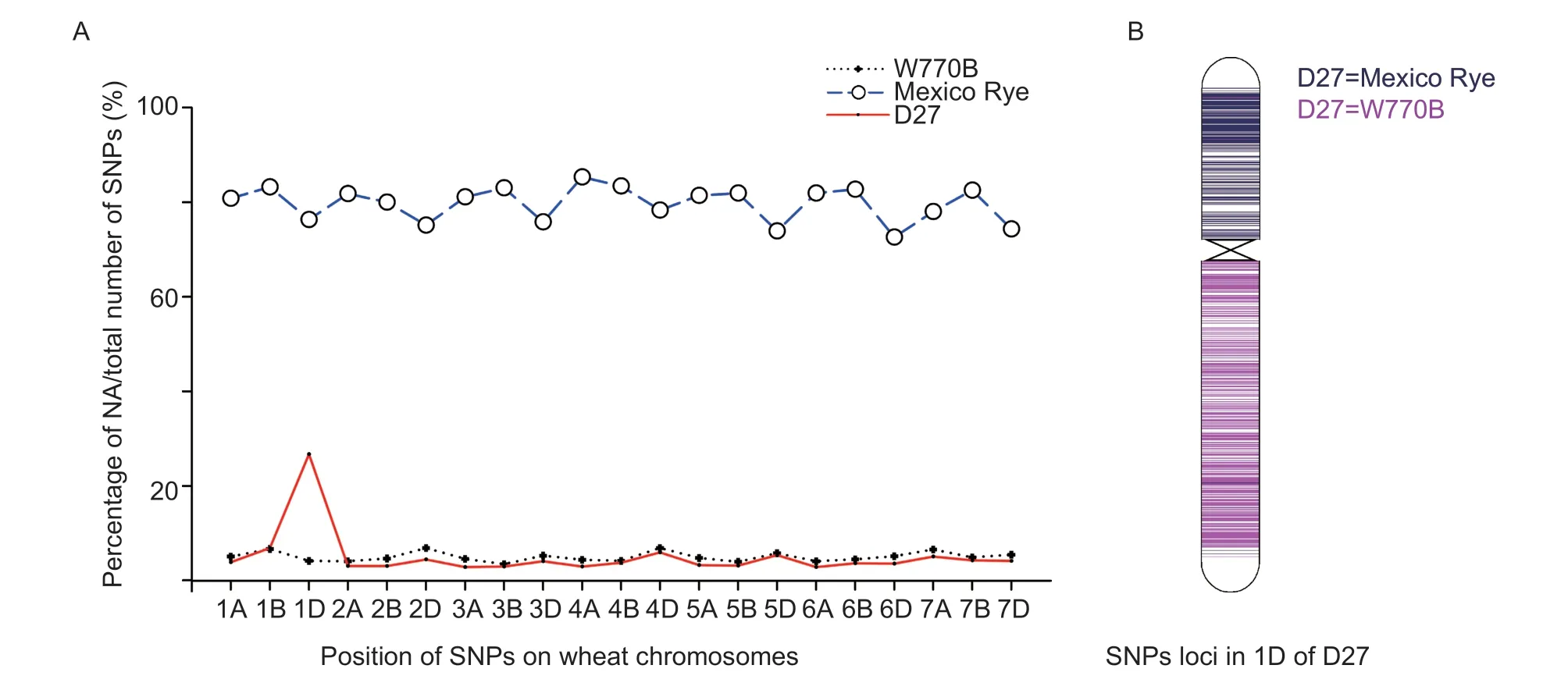
Fig.6 Analysis of the chromosomal composition of line D27 using a wheat 40K SNP array.A, percentages of deletion genotype SNP markers in each chromosome (null allele (NA) ratio).The clear difference in the 1D chromosome indicates a chromosomal rearrangement in this position.B, the same SNP genotype markers in D27 and their parents were on the 1D chromosome according to their physical positions.

Table 2 Rye sequence characterized amplified region (SCAR) markers and wheat simple sequence repeat (SSR) markers used in this study to analyze the chromosomal composition of line D27
3.6.Gliadin subunit patterns
A-PAGE was used to separate the gliadin proteins from line D27 and its parents.As shown in Fig.7, Mexico Rye, LXH, and line D27 all produced the same specific γ-gliadin bands.Clearly, line D27 produced the same protein band pattern as its common wheat parent W770B, but it also produced the ryeSec-1gene bands, which are regarded as characteristic of rye 1RS.The genes encoding secalin map onto the short arm of the rye 1R chromosomes, so line D27 possessed rye 1RS chromosomes.
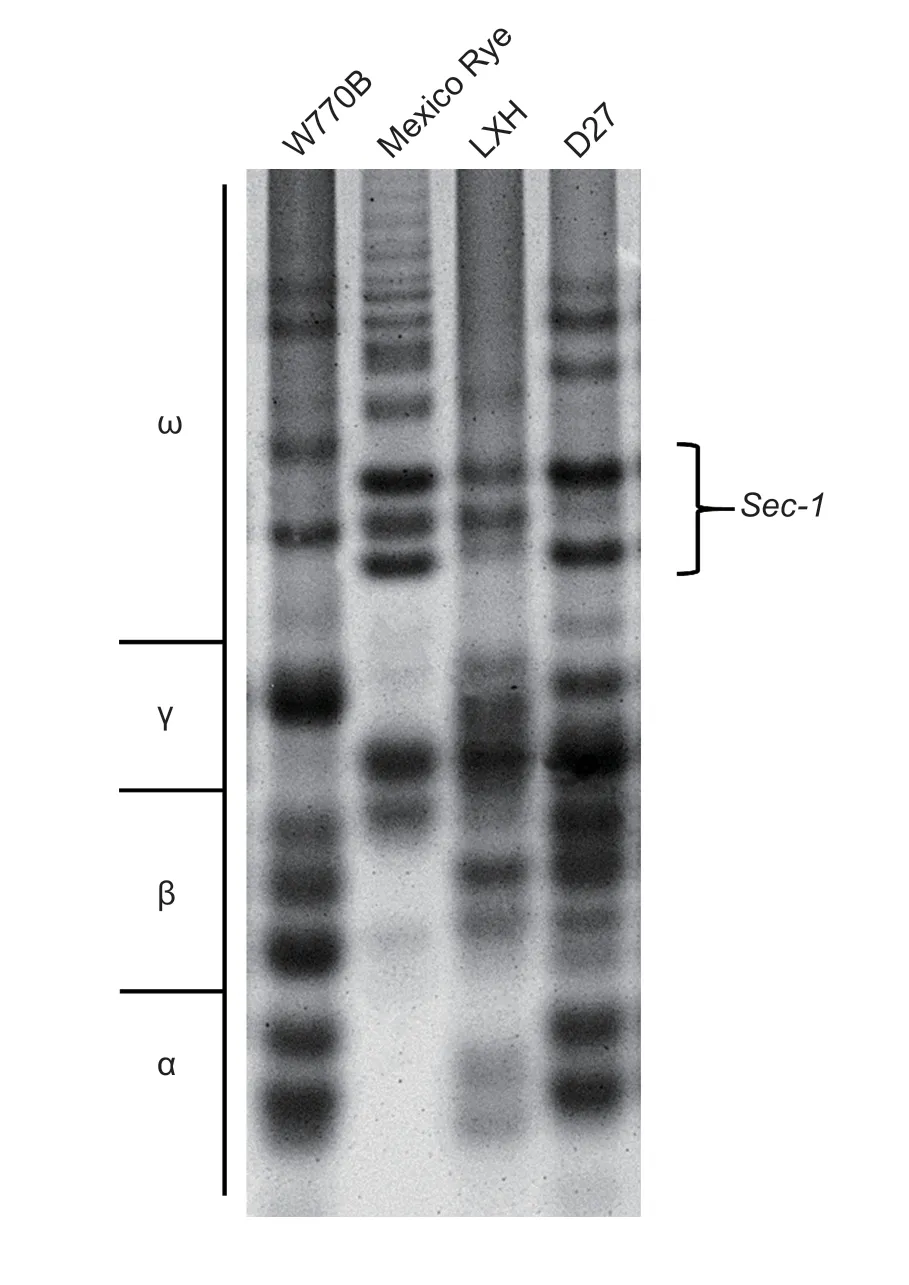
Fig.7 Analysis of gliadin subunits using acid-polyacrylamide gel electrophoresis.Line D27 produced a similar subunit pattern to its wheat parent W770B, but it also included some subunit bands from Mexico Rye such as a Sec-1 gene-specific band.
3.7.Agronomic traits and disease resistance performance
The agronomic traits assessed for line D27 and its parents are shown in Fig.8-A–C and Table 4.Line D27 was similar to wheat W770B in terms of its morphological traits, such as the plant height, spike length, spike type, and kernel weight (atP=0.05 andP=0.01), rather than its alien parent Mexico Rye.The tiller number and kernels per spike of line D27 were higher than that of W770B.The results showed that line D27 was mainly inherited from wheat W770B, but the increased tiller number and kernels per spike might have been caused by the introduction of the rye 1RS chromosome.
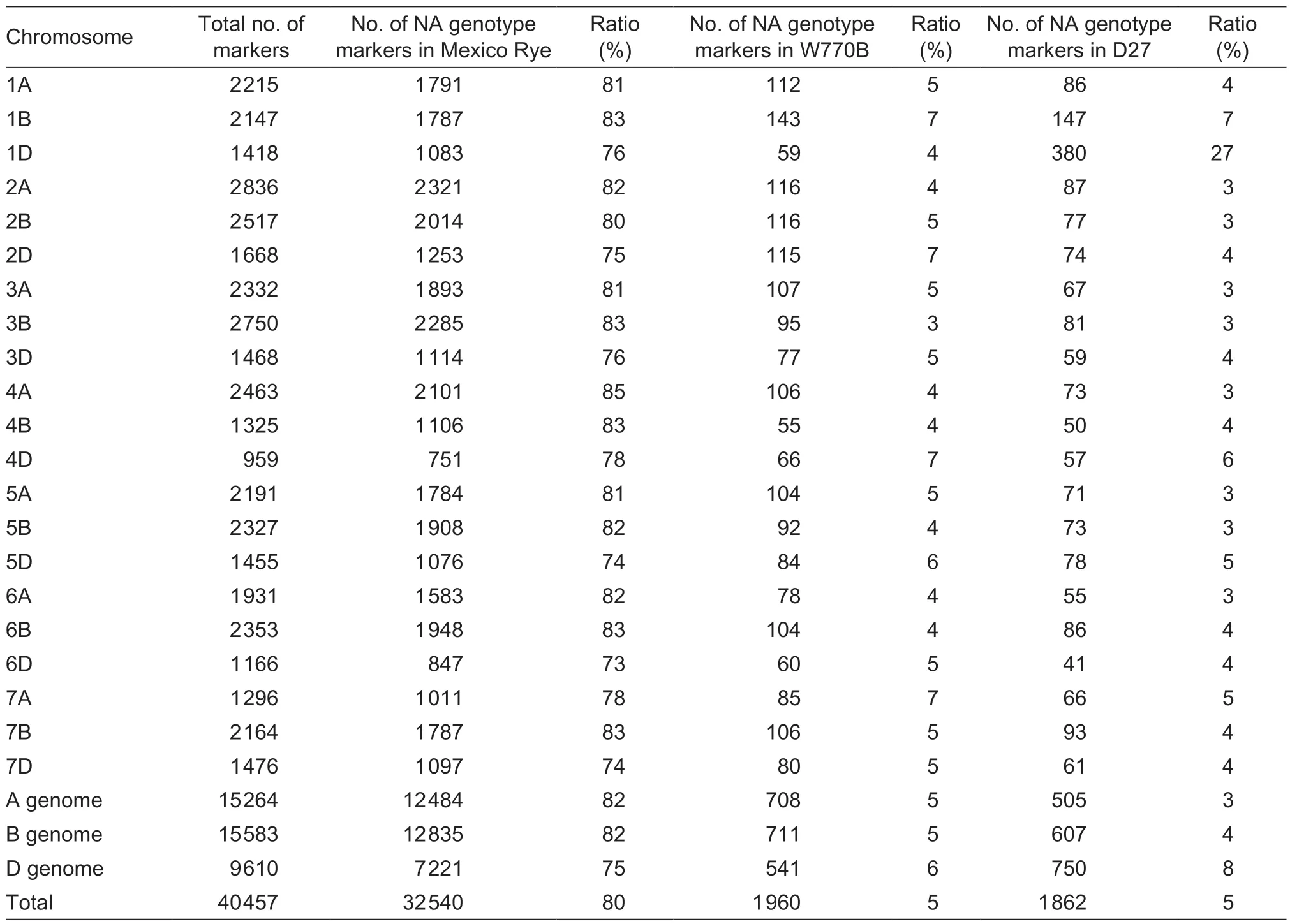
Table 3 Wheat 40K single nucleotide polymorphism (SNP) array analytical data for Mexico Rye, wheat W770B, and line D271)

Table 4 Comparison of the agronomic traits of line D27 and its parents
The responses of the materials to mixedPstraces were tested under the same conditions in the seedling and adult stages by artificial inoculation.The ITs determined for the four materials were as follows: in the seedling stage (Fig.8-D), IT=4 for the susceptible control CS, IT=3 for W770B, IT=0; for D27, and IT=0 for Mexico Rye; and in the adult stage (Fig.8-E), D27 had significantly fewer spores attached to its secondary leaf than wheat parent W770B and T1BL·1RS variety Shaan 229.Thus, line D27 was almost immune to stripe rust at the seedling and adult stages in the same manner as its alien parent Mexico Rye.
We also evaluated the resistance of the four materials toBgtisolate E09.The alien parent rye was immune to powdery mildew both at the seedling and adult stages, but the resistance to disease was different in line D27.The ITs determined for the four materials were as follows: at the seedling stage (Fig.8-F), IT=4 for Chinese Spring, IT=3 for W770B, IT=1 for D27, and IT=0 for Mexico Rye; and at the adult stage (Fig.8-G), even though D27 had some spores on its leaf, it still expressed better resistance to E09 isolate than its wheat parent W770B.

Fig.8 Agronomic traits and disease tolerance.A, mature plants.B, spikes of plant materials.C, spikelets and kernels.D, symptoms in response to wheat stripe rust in the seedling stage.E, symptoms in response to wheat stripe rust in the adult stage.F, symptoms after inoculation with Bgt isolate E09 in the seedling stage.G, symptoms after inoculation with Bgt isolate E09 in the adult stage.1, Mexico Rye; 2, W770B; 3, D27; 4, Chinese Spring; 5, Shaan 229.
3.8.Starch gelatinization and dough viscoelastic properties of line D27
The starch gelatinization properties of the material were measured by a rapid viscosity analyzer (RVA) (Fig.9).The results showed the gelatinization curve first increased, decreased, and then increased in the program-measured range.The peak viscosity of starch in Mexico rye was significantly lower than that in other materials, where W770B was the highest and Shaan 229 was slightly higher than D27.These results indicated that the gelatinization properties of different materials were related to genotype.The dough viscoelastic properties of materials were measured by rheometer (Fig.10).The results showed the value of dough modulus and tan δ in four materials all presented frequency dependence.In detail, the dough modulus (G´ and G´´) increased with the increase in frequency, but the tan δ value presented a trend of decreasing and then increasing.Among them, W770B showed the highest value of dough modulus and the lowest value of tanδ, and Mexico Rye was on the contrary.In addition, although the value of dough modulus in D27 was less than Shaan 229, its value of tanδ was higher.
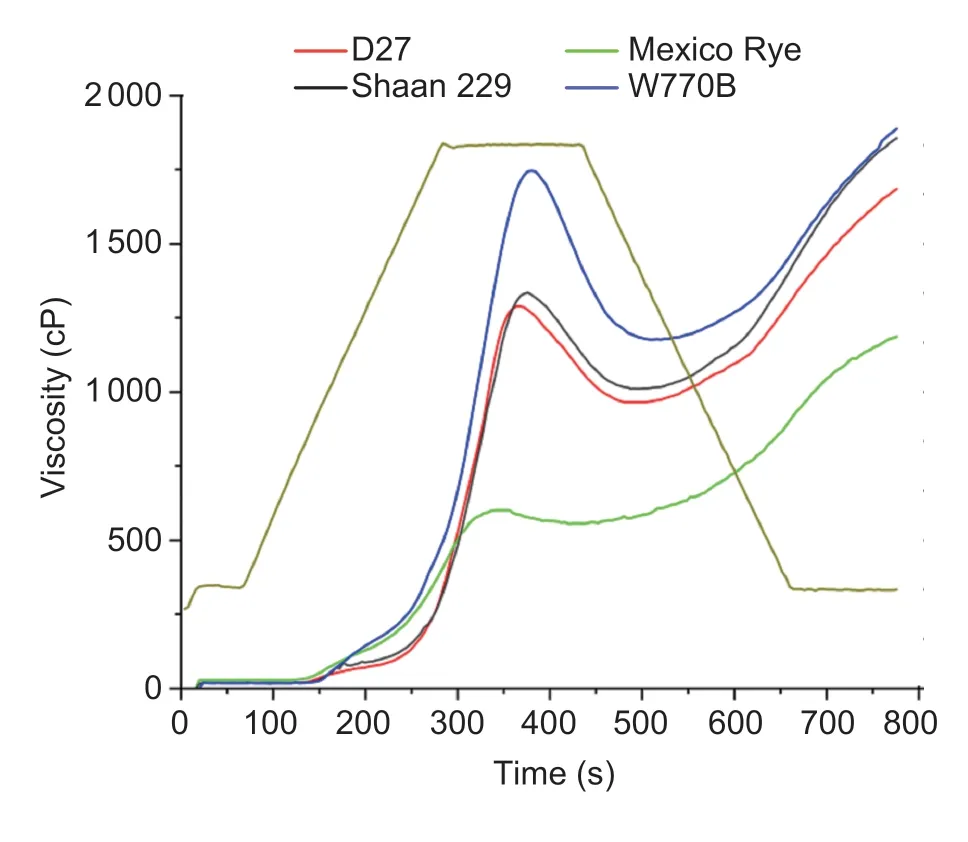
Fig.9 Differences in rapid viscosity analyzer (RVA) gelatinization curves of starch among materials.
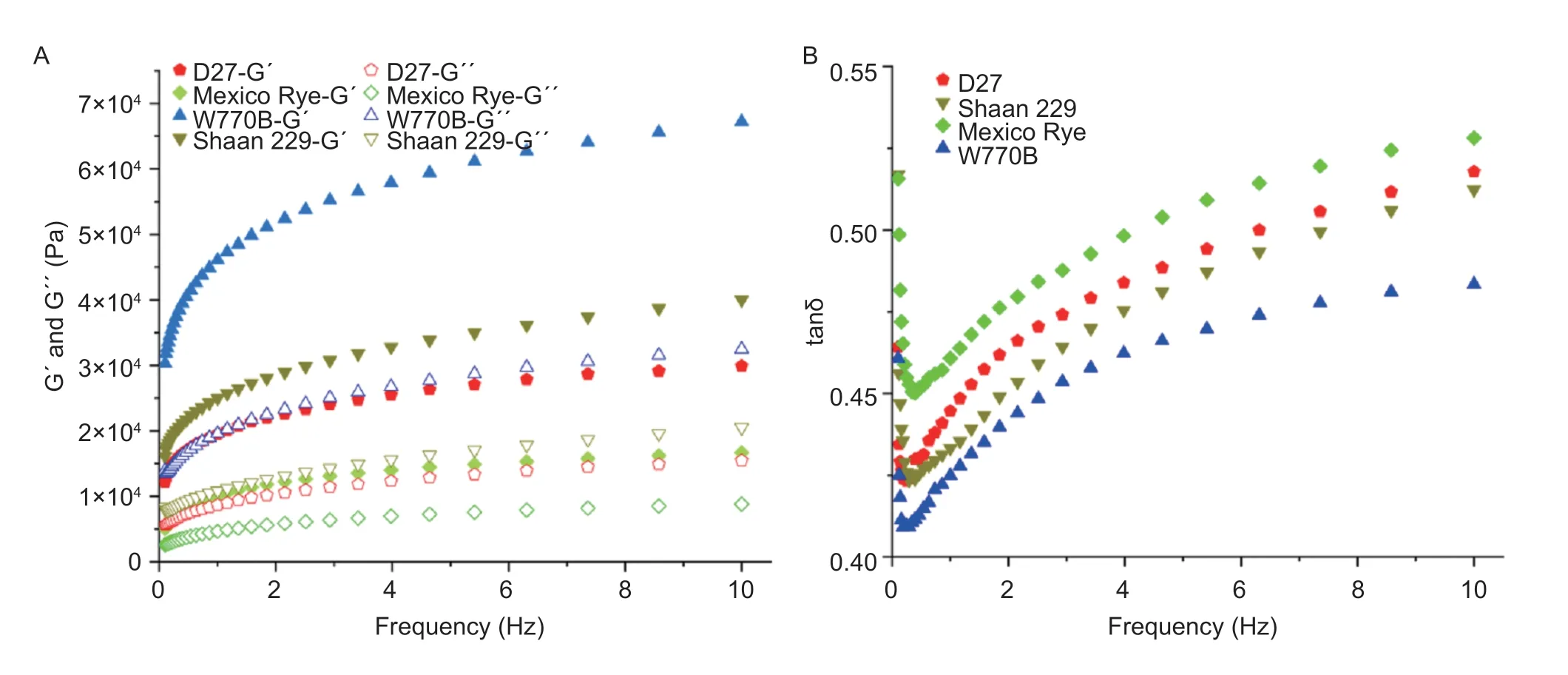
Fig.10 Differences in rheological properties of dough between different materials.A, the storage modulus (G´) and loss modulus (G´´) of materials.B, the tanδ value of materials.
4.Discussion
4.1.New genetic type 1R chromosome still has great potential to improve wheat breeding
Among the species related to wheat, rye has made important contributions to wheat breeding throughout the world.Various types of wheat–rye derived lines have been developed, including 1R, 4R, 6R, and 7R addition lines (Schneideret al.2016; Yanget al.2016; Anet al.2019); 1R(1A), 1R(1B), 1R(1D), 2R(2D), 5R(5A), 5R(5B), 6R(6A), 6R(6B) and R genome (D genome) substitution lines (Bielig and Driscoll 1970; Leeet al.1970; Krasilovaet al.2012; Liet al.2016); T1AL·1RS, T1BL·1RS, T2BS·2RL, T2DS·2RL, T4R·5D, T5A·5R, T6AL·6RS, and T7DS·7DL-1RS translocation lines; and some introgression lines (Liet al.2006, 2015; Leiet al.2013; Fuet al.2014; Zhanget al.2020).In particular, T1BL·1RS translocation lines are favored by breeders because the 1RS chromosome contains desirable genes that can enhance the disease resistance of wheat (Renet al.2012).Many high-yielding and disease-resistant varieties are regarded as key parents because of their 1BL·1RS chromosomes, such as Kavkaz and Predgornia (Haoet al.2008).Indeed, statistics show that the 1BL·1RS chromosome is present in about 1 050 wheat cultivars around the world (Schneideret al.2016).However, there are only a few reports about wheat–rye T1DL·1RS translocation lines, according to Koebneret al.(1986) and Lukaszewski (2000).Valuable genes have been transferred to wheat, mainly from the cultivated rye variety Petkus Roggen, where the 1RS chromosome contains theYr9,Pm8,Sr31, andLr26resistance genes (Kozubet al.2017).Previous studies suggested that new pathogenic races had led to the gradual loss of function forYr9andPm8genes.However, recent studies show that there are great genetic differences among different subspecies ofSecaleand different local varieties of cultivated rye, which means the 1RS chromosomes in different rye varieties may possess different alleles that mediate responses to diseases.Thus, the 1R chromosome still has great potential for wheat breeding (Qiet al.2016).In the present study, we identify a novel derived line called D27 where the 1DS chromosomes are replaced by the 1RS chromosomes from Mexico Rye, and the line exhibits desirable resistance to wheat stripe rust and powdery mildew.
4.2.The wheat–Mexico Rye translocation line D27 inherits stably
The offspring developed by distant hybridization usually have undesirable characteristics, such as chromosomal number variation and disordered pairing or inheritance, which might lead to unstable morphological traits (Cifuentes and Benavente 2009).Therefore, it was necessary to ensure that the chromosomal composition and transmission were stable based on classical cytogenetic observations and GISH analysis.Observations of RTCs showed that line D27 had 42 chromosomes, with two translocated chromosomes comprising whole-arm rye chromosomes and whole-arm wheat chromosomes.Further observations of PMCs showed that the translocated alien chromosomes shared the same homoeology.They could be duplicated, paired, segregated, and inherited normally as common wheat chromosomes.The results demonstrated that wheat–rye derived line D27 was a genetically stable translocation line and a potential germplasm resource.
4.3.Multiple markers show the 1DS of D27 is replaced by 1RS
Chromosomal instability and genomic rearrangements have been clearly demonstrated in wheat–rye derived lines (Szakács and Molnár-Láng 2010).The FISH analysis is the most reliable and intuitive method for detecting chromosomal structure rearrangements and replacements inTriticeaespecies.It can accurately distinguish 42 wheat chromosomes based on fluorescent signals with different colors distributed on each chromosome (Sidhuet al.2008).The FISH analysis can be conducted with various types of Oligoprobes, such as pAs1 (A and D genomes), GAA (B genome chromosomes), pTa535 (A and D genome chromosomes), pSc119.2 (B genome chromosomes), oligo-B (B genome chromosomes), Oligo-D (D genome chromosomes), and pTa71 (1B, 6B, and 5D chromosomes) (Tanget al.2014, 2018).Using a combination of Oligo-pSc119.2 and Oligo-pTa535, our study showed that the 1DS chromosomes in line D27 had been replaced by rye 1RS chromosomes, and the results were supported by sequential GISH.About 83–92% of repetitive sequences exist in the genomes ofTriticeaespecies, and they are useful for detecting related species and interspecific hybridization progeny (Anamthawat-Jónsson and Heslop-Harrison 1993; Zhaoet al.2021).The similarity of rye and wheat genomes facilitates the widespread adaptation of Oligo-probes such as pSc119.2, pTa71, and pAWRC.1 (rye and wheat centromeres), but the different locations of repetitive sequences lead to diverse FISH karyotypes (Fuet al.2015).Therefore, the recombination of 1D and 1R chromosomes was successfully detected in line D27.
After demonstrating the presence of a pair of translocated chromosomal arms in line D27, we elucidated the homoeologous group of the alien chromosomes.According to a previous study, different types of genomespecific molecular markers could be applied to determine the homoeologous relationships between introduced alien chromosomes and lost wheat chromosomes.For example, a wheat–rye 4R addition line was identified using SCAR and specific-locus amplified fragmentsequencing (SLAF-sequencing) markers (Anet al.2019), and a 1R(1B) substitution line and T1BL·1RS translocation line were identified by using SSR and SCAR markers (Liet al.2016; Qiet al.2016).Thus, SCAR and SSR markers were designed based on the specific sequences of rye and wheat chromosomes.Four pairs of SCAR markers for 1RS and other rye genotypes successfully detected the Mexico Rye 1RS chromosomes in line D27.In addition, some SSR markers on the wheat 1DS chromosomes amplified D genome-specific bands in W770B but not in line D27.The introduction of the rye secalin gene degrades the baking quality of wheat (Grayboschet al.1990), but it can be used to identify the rye 1R chromosome.Thus, the secalin gene located on the rye 1RS chromosome can be regarded as a tag to detect the presence of this chromosome.We used A-PAGE to detect the presence of aSec-1genespecific band, and the results demonstrated that line D27 possessed rye 1RS chromosomes.These results indicated that line D27 was a wheat–rye T1DL·1RS translocation line.
In the early stages of breeding, the introduction of desirable agronomic traits into wheat usually occurredviamany rounds of hybridization and long-term screening, but without understanding the underlying molecular mechanism (Rasheedet al.2018; Chenget al.2021).PCR-based molecular markers have made great contributions to gene discovery and assisted selection in breeding, but their shortcomings limit their further application (Kamaluddinet al.2017; Griffithset al.2022).The release of the complete genome of common wheat and decreasing costs of high throughput sequencing mean that array-based genotyping techniques are increasingly employed for gene mapping, targeted genotyping, and molecular-assisted selection (Consortium 2018; Sunet al.2020).Four wheat-alien derived lines were accurately identified using different types of wheat SNP arrays by Liet al.(2019, 2021), Wang Y Zet al.(2020), and Baiet al.(2021).In this study, the genotypes of line D27 and its parents were compared at each locus with wheat 40K SNPs, which covered 21 pairs of chromosomes.The results, together with molecular markers and NDFISH analysis, showed that line D27 was a wheat–rye T1DL·1RS translocation line.Compared with other studies, we found that the 40K array is a newly developed liquid SNP array, and it provides a different analytical method compared with the previous 15K and 55K solid SNP arrays (Liet al.2019, 2021).In particular, solid SNP arrays are mainly used to compare the heterozygosity rate of SNPs on each chromosome, whereas liquid SNP arrays are generally used to compare the deletion rates of SNPs on each chromosome.
4.4.Stripe rust resistance of D27 is superior to its wheat parent and traditional 1B/1R lines
Except for B chromosomes, the characteristics of plants are usually changed after introducing or replacing with the alien chromosome(s) (Camachoet al.2000).However, three sets of chromosomes give the wheat genome greater buffer capacity to tolerate alien chromosomes (Joneset al.2008).The changes in wheat may include morphological traits, stress tolerance, and ripening time (Wanget al.2019; Efremovaet al.2021).Excellent traits would increase the value of wheat-alien derived lines.Line D27 exhibited high resistance to wheat stripe rust and improved resistance to powdery mildew strikingly better than its wheat parent and traditional T1BL·1RS variety Shaan 229, thereby demonstrating that the resistance genes in line D27 came from Mexico Rye.Similarly, the 1RS chromosome ofSecaleafricanumprovides superior resistance toYr9-virulent races, thePm17gene on the 1RS chromosome from Insave Rye is different from thePm8gene of Petkus, and Imperial Rye carrying theSrRgene exhibits greater stem rust resistance than that with theSr31gene on the 1RS chromosome (McIntoshet al.1995; Schlegel and Korzun 1997; Yanget al.2009).Therefore, it would be useful to investigate the potential variability in resistance genes on the 1RS chromosome from Mexico Rye in line D27.
4.5.Mexico Rye 1RS make raising thermal stability of starch and less elastic of dough
Wheat starch has a structure of semi-crystalline granule that makes starch suspension express gelatinization after being heated to a certain temperature, which is related to the taste of food.Therefore, the gelatinization properties of wheat starch can be used to evaluate the quality of wheat thermal processing (Akramet al.2011).Zhang and He (2002) showed that peak viscosity and breakdown were two important indexes of gelatinization properties.The peak viscosity was positively correlated with the elasticity, toughness, and eating quality of flour products.The breakdown reflected the stability of starch paste after being heated, and a smaller breakdown meant better thermal stability of starch paste, so better smoothness and soft cotton of flour products (Crosbie 1991).In this study, the peak viscosity of starch in common wheat 7182 was higher than that in the two 1RS translocation lines and Mexico Rye, which indicated that the elastic-related quality of recipient wheat deteriorated due to the introduced rye chromosomes.However, from the breakdown, D27 was significantly lower than that of Shaan 229 and wheat parent W770B, which indicated that, because of the translocation between 1RS and 1DS, the starch of D27 was more tolerant to high temperatures, and its flour had better thermal stability.
The viscoelastic properties of dough show that the value of G´ and G´´ increased with the increase of frequency in the scanning range, and G´ was higher than G´´ (Lopes-da-Silvaet al.2007).In addition, tanδ was found to decrease at low frequencies, which may be caused by insufficient hydration of dough (Kimet al.2008).Subsequently, tanδ increased gradually at high frequencies, which indicated that the viscosity of dough gradually plays a role with increasing frequency and makes the dough system more fluid (Singh and Singh 2013; Sarabhaiet al.2017).In this study, G´ and G´´ of Mexico Rye were the lowest, but tanδ was the highest.This result indicated that materials containing R genomes had higher viscosity and lower elasticity.Similarly, compared with W770B, D27 and Shaan 229 containing 1RS also showed greater viscosity and lower elasticity, but the degree of viscoelastic change of D27 was more obvious than that of Shaan 229.This phenomenon supported previous studies that the genome D>B>A in terms of contribution to dough elasticity (Dhaliwalet al.1987; Guptaet al.1994).It was worth noting that the slope of D27 in tanδ was the highest, consistent with the results of gelatinization properties.The reason might be that its starch rapidly bound to water, which increased the interlacing phenomenon of starch particles and improved viscosity.D27 lost its 1DS, and Shaan 229 lost its 1BS due to inter-chromosomal translocation, which caused D27 to express less elasticity but more viscosity.According to the report, strong gluten wheat dough shows higher elasticity and is suitable for making bread, noodles, and dumplings, while low gluten wheat dough shows higher viscosity and is suitable for making cakes, biscuits, and cookies (Huebneret al.1999).Therefore, for breeding application, D27 with high dough viscosity is more conducive to the cultivation of low-gluten wheat variety materials.
5.Conclusion
We conducted cytogenetic, molecular marker, and agronomic analyses of line D27 and characterized its chromosomal composition, stable inheritance, exogenous source, and agronomic traits.As a result, line D27 was identified as a novel wheat–rye T1DL·1RS translocation line with a higher tiller number, superior stripe rust resistance, and higher dough viscosity than its wheat parent.Therefore, line D27 represents a valuable germplasm source for breeding multi-resistant and highyield wheat.Additionally, it provides a new rye 1RS chromosome genotype for further research.
Acknowledgements
This study was supported by the National Natural Science Foundation of China (31771785) and the Key Research and Development Program of Shaanxi, China (2018ZDXM-NY-006).
Declaration of competing interest
The authors declare that they have no conflict of interest.
Appendixassociated with this paper is available on https://doi.org/10.1016/j.jia.2022.08.039
杂志排行

Journal of Integrative Agriculture的其它文章
- Herbicidal activity and biochemical characteristics of the botanical drupacine against Amaranthus retroflexus L.
- Developing a duplex ARMS-qPCR method to differentiate genotype l and ll African swine fever viruses based on their B646L genes
- The effects of maltodextrin/starch in soy protein isolate–wheat gluten on the thermal stability of high-moisture extrudates
- Elucidation of the structure, antioxidant, and interfacial properties of flaxseed proteins tailored by microwave treatment
- Effects of planting patterns plastic film mulching on soil temperature, moisture, functional bacteria and yield of winter wheat in the Loess Plateau of China
- lnversion tillage with straw incorporation affects the patterns of soil microbial co-occurrence and multi-nutrient cycling in a Hapli-Udic Cambisol