大白菜核不育花药超显微结构观察
2023-05-08岳艳玲郭瑛琪
岳艳玲,郭瑛琪,2
(1.云南农业大学园林园艺学院,昆明 650201;2.中国科学院昆明动物研究所,昆明 650223)
大白菜(Brassica campestrisL.ssp.pekinensis)属于十字花科芸薹属白菜种,原产于中国,有悠久的栽培历史,是我国十分重要的蔬菜作物。大白菜为两性花异花授粉植物,杂种优势非常显著,但大白菜花器比较小,很难通过人工授粉的方式满足F1杂交种子的商业化利用。
雄性不育(male sterility)是指植物雄蕊退化或不能形成有活力花粉的现象,在自然界中广泛存在,是大白菜实现杂种优势利用的重要途径。应用雄性不育系进行杂交种子生产,杂种纯度高,不存在母本自交种子混杂问题。因此,雄性不育发生机理及其应用研究受到生物学家的广泛关注。1995 年,FENG 等[1]发现了一类大白菜核不育的遗传机制,提出了“复等位基因遗传假说”。复等位核雄性不育具有雄蕊退化彻底、不育性稳定等特点,不育度和不育株率均为100%,且不受细胞质影响,没有正、反交遗传效应,是一类比较理想的大白菜雄性不育材料。2005 年,岳艳玲等[2-4]以“复等位基因遗传假说”为理论基础设计了合成转育方法和定向转育方法,成功将该不育基因向不同生态型大白菜中进行转育,育成多个大白菜核基因雄性不育系,并对其雄性不育的败育机制进行研究。10L03是在大白菜复等位基因雄性不育系定向转育过程中经过多代回交获得的近等基因系,不育株雄蕊败育彻底,雌蕊正常,无不良性状伴生,植株结籽特性好。石蜡切片观察发现大白菜花药绒毡层细胞结构异常,四分小孢子不能发育成正常的单核小孢子,最终药腔空瘪,发生败育[5]。为进一步深入研究该雄性不育材料的败育机制,本研究采用透射电子显微技术,观察小孢子败育过程花药绒毡层细胞及小孢子的发育动态,明确不育发生时期及败育细胞的超微特征,为揭示该核不育小孢子的败育机制提供理论依据。
1 材料与方法
1.1 材料
大白菜核不育近等基因系10L03,系统内兄妹交后代育性保持1∶1 分离,可育株自交后代可育株与不育株3∶1分离,材料由本课题组选育而成。试材种植于云南农业大学蔬菜基地大棚内,于盛花期取不同育性的大白菜花序的花药用于固定。
1.2 方法
采用2.5%戊二醛固定液(PBS pH=7.4)进行可育、不育花药前固定,抽真空,4 ℃固定48 h。PBS 浸洗3 次,每次15 min,加入2%的锇酸后固定4 h。PBS 浸洗3 次,每次15 min。梯度乙醇50%、70%、95%、100%逐级脱水,树脂包埋。使用LEICA EM TRIM 树脂修块机进行修块,LEICA EM UC6超薄切片机先切600~1000 nm的半薄切片,1%甲苯胺蓝染色10 s,OLYMPUS CX31 显微镜观察花药发育时期。然后70 nm 连续切片,100 目铜网捞片,醋酸双氧铀染色25 min,双蒸水涮洗,柠檬酸铅染色8~15 min,双蒸水涮洗终止染色,并放入标好号的滤纸上。EOL JEM-1200EX Ⅱ电子显微镜观察,拍照。
2 结果与分析
2.1 可育花药细胞发育过程
透射电镜观察花粉母细胞时期可育花药结构,花药外层细胞分化明显,视野中可以清晰地看到花药中层细胞和绒毡层细胞,绒毡层细胞牙齿状排列紧凑,径向长度5 μm以上,可以看到内质网。花粉母细胞位于药腔中央,细胞排列紧密,形状不规则,细胞核清晰可见,绒毡层和花粉母细胞饱满,内含物充实(图1A)。进一步放大观察绒毡层细胞,细胞核核仁电子密度大,几乎与细胞核等大,细胞质中含有淀粉等物质(图1B)。花粉母细胞较大,可看到核仁和核质,细胞之中含有丰富的内含物(图1C)。
花粉母细胞之后,可育绒毡层细胞核完整,细胞核轮廓清晰,核仁较大,电子密度高(图1D)。花粉母细胞细胞核、核仁明显,核膜完整,清晰可见(图1E)。可育花药继续发育到四分体时期,绒毡层细胞仍可见细胞核,且核仁电子密度明显大于核质,有的绒毡层为单层细胞(图1F),而有的为2层或3层细胞(图1G),绒毡层细胞质内含物丰富(图1F、图1G),细胞间存在细胞间隙,而绒毡层靠近外层细胞的细胞膜可见,放大可看到绒毡层细胞中的线粒体、内质网、分泌小泡(图1H)。四分小孢子圆润饱满,小孢子电子密度大,小孢子表面和细胞间可见胼胝质,小孢子内含物丰富,可见脂滴体(图1I)。
可育花药单核早期小孢子呈三叶草形(图1J)。绒毡层细胞电子密度大,可见细胞核、内质网,细胞边缘有分泌小泡,绒毡层细胞含有淀粉粒等,可见核小体(图1K)。单核小孢子细胞核很大位于小孢子中央,核膜清晰可见,核仁染色深,电子密度大,无明显液泡,但细胞质中可见许多囊泡(图1L)。
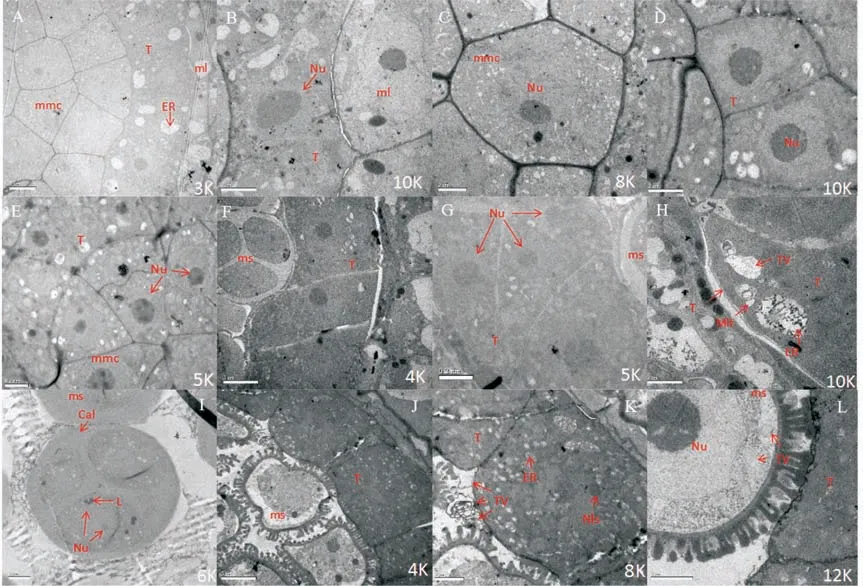
图1 可育花药花粉母细胞至单核早期细胞超微结构Figure 1 Cell ultrastructure of the fertile anther during pollen mother cell to early stage of mononuclear microspore
小孢子继续发育,绒毡层细胞间隙进一步变宽,同一时期绒毡层细胞电子密度比小孢子大(图2A),绒毡层细胞淀粉粒等物质增多,观察到的细胞核无明显核膜,但核仁清晰(图2B),单核小孢子中出现液泡,可见细胞核核仁及核膜、线粒体及分泌小泡,形状从三叶草形逐渐转变为圆形(图2C)。
到了单核晚期,绒毡层细胞电子密度降低,小孢子继续向圆形转变(图2D),绒毡层细胞进一步降解,细胞内的淀粉粒等更为丰富,细胞形态无规则,有的绒毡层细胞隐约可见细胞核,细胞周围有分泌小泡(图2E)。小孢子内有大液泡,液泡膜清楚可见,核膜清晰,核仁色深,细胞质内可见小囊泡,线粒体以及脂滴体(图2F)。
双核期绒毡层细胞出现大量空泡,径向变短,横向变长,此时绒毡层细胞结构基本消失,观察不到完整的细胞器。小孢子基本圆形,可见两个细胞核,液泡变小(图2G)。绒毡层细胞看不到细胞核、细胞器及细胞膜结构,细胞间缝隙进一步变大,绒毡层细胞基本消失(图2H)。小孢子进一步放大观察可见分泌小泡、糖类、脂滴体等(图2I)。
花药成熟时期,可育花药绒毡层细胞结构消失,花粉粒电子密度极大(图2J),绒毡层细胞只剩薄薄的一层,但不同位置薄厚不一,有的绒毡层细胞已经完全消失,外侧细胞含有线粒体和小囊泡(图2K)。花粉粒圆形饱满,电子密度极大,可见细胞核(图2L)。

图2 可育花药单核至成熟期细胞超微结构Figure 2 Cell ultrastructure of the fertile anther during mononuclear microspore to mature stage
2.2 不育花药细胞发育过程
透射电镜观察花粉母细胞时期不育花药结构,视野中可见花药表皮细胞、内壁细胞、中层细胞和绒毡层细胞,绒毡层细胞状排列紧凑呈牙齿状,花粉母细胞位于药腔中央,排列紧凑,形状不规则,细胞核清晰可见,绒毡层和花粉母细胞饱满,内含物充实(图3A)。绒绒毡层细胞核及核仁明显,核膜可见,细胞质中含有淀粉粒等(图3B)。花粉母细胞较大,核仁电子密度大,核质电子密度小,细胞之中含有丰富内含物(图3C)。
花粉母细胞后期,不育绒毡层细胞间出现缝隙,细胞核裂解出现核小体(图3D),说明绒毡层细胞发生了细胞程序性死亡。花粉母细胞的细胞核、核仁以及核膜都明显可见(图3E);花粉母细胞可以完成减数分裂发育成四分体,此时绒毡层细胞膨大,四分小孢子变形(图3F),细胞内有空泡,核内可观察到核小体(图3G),说明此时小孢子发生细胞程序性死亡。绒毡层细胞空泡化明显,细胞核电子密度大,可以看到核小体(图3H)。

图3 不育花药花粉母细胞至四分体时期细胞超微结构Figure 3 Cell ultrastructure of the fertile anther during pollen mother cell to tetrad stage
四分体之后,绒毡层细胞空泡化现象加重,细胞或变形(图4A)或异常膨大(图4C),之后发生降解(图4D)。小孢子有的细胞核皱缩,细胞质中出现大量的小囊泡,胼胝质仍然包裹小孢子(图4B),说明包围四分小孢子的胼胝质并未被及时降解,没能顺利释放单核小孢子。绒毡层细胞和小孢子的败育过程并不同步,特征也不完全一致,有的绒毡层细胞会发生径向伸长继续膨大(图4A、图4C、图4E),有许多分泌小泡,同一腔室中小孢子败育程度也有不同,有的皱缩,有的勉强维持圆形(图4E),但都已看不到任何细胞器和细胞结构,但胼胝质仍然存在。成熟期,不育花药绒毡层细胞和小孢子已经无法分清层次结构(图4F、图4G)。15K 倍数下可见细胞壁层叠状,药腔中可见电子密度极大的细胞残体(图4H)。

图4 四分体之后不育花药败育的细胞超微结构Figure 4 Cell ultrastructure of the fertile anther during the stage after tetrad
3 讨论与结论
花粉发育是有性繁殖植物完成其世代交替的重要生命过程,涉及到一系列的细胞学和生物化学变化,任何一个环节发生改变都可能会导致花粉发育异常,从而引起雄性不育[6]。在大白菜不育系中,败育可发生在小孢子发育的各个时期,如无花粉囊型败育型,发生在孢原细胞时期,造孢细胞无法形成或相互黏连;花粉母细胞败育型,表现为减数分裂异常,小孢子无法分离,四分体无法散开;单核花粉败育型表现为花粉壁形成异常,花粉无活性[7-13]。PU 等[5]利用制作石蜡切片,采用光学显微技术观察10L03 不育花药发育过程,花粉母细胞经减数分裂可以形成四分体,但小孢子发生变形,绒毡层变厚,提前降解,说明10L03 花药不育发生在四分体时期。本研究对10L03 花药细胞超微结构进行观察,发现的不育花药绒毡层细胞细胞核在减数分裂时已经分解形成核小体,说明此时绒毡层细胞已经发生细胞程序性死亡,花药发生败育的时期应是花粉母细胞减数分裂时期,也说明了细胞结构观察结果较组织结构观察结果更准确。
绒毡层是药室的最内层,由雄蕊原基的L2、L3 层共同发育而来,包围着花粉母细胞和小孢子并与其相互作用,绒毡层的发育及其降解对小孢子的发生和发育起着至关重要作用[14-15]。绒毡层缺失、过早或延迟降解,甚至不降解,均可导致植物雄性不育[16]。郭瑛琪等[17]观察发现10L03 不育花药绒毡层提前降解,本研究电镜观察结果与郭瑛琪等观察结果一致,但观察到绒毡层细胞多泡化、膨大空泡化及纵向延伸膨大空泡化3种不同的败育形式。通过透射电镜,本研究观察到10L03不育花药绒毡层细胞在减数分裂时期细胞核裂解形成核小体,败育小孢子细胞也出现核裂解现象,这是一个明显的细胞程序性死亡特征,说明10L03不育花药败育过程中出现异常PCD过程,这与郭瑛琪等[17]采用TUNNEL法检测的结果相一致。
胼胝质(callose)是一种以β-1,3-键结合的葡聚糖,广泛分布于高等植物的筛管、细胞板和胞间连丝,在植物细胞分裂、细胞到细胞的分子运动及抗生物、非生物胁迫等生命活动中发挥着重要的调节作用[18]。胼胝质还是花粉发育的重要调控者特定发育阶段的胼胝质沉积和降解是小孢子发生和花粉形成的重要前提[19]。在减数分裂前,胼胝质开始在花粉母细胞的细胞壁中沉积,减数分裂后四个子细胞由胼胝质隔开,参与胞质分裂,整个四分体都被胼胝质壁包裹着,随后胼胝壁被消化而释放小孢子,花粉外壁形成、花粉萌发和花粉管生长也需要胼胝质的参与[20-21]。四分体周围的胼胝质壁提前降解或延迟降解以及花粉外壁中胼胝质沉积异常均会影响小孢子进一步发育,引起花粉败育,导致雄性不育[22-23]。PU 等[5]利用Aniline blue 染色和荧光检测发现10L03花药败育过程中,包裹着小孢子的胼胝质不能被降解,不能发育成单核小孢子,花药特异表达胼胝质酶基因AtA6的同源基因BraA04g008040、BraA07g009320、BraA01g030220、BraA03g040850在不育花药减数分裂后低表达可能促进了小孢子胼胝质的沉积[5]。胼胝质酶是由绒毡层细胞合成并分泌到药腔中,降解包裹四分体的胼胝质,从而释放单核小孢子。本研究电镜观察到不育花药绒毡层细胞提前发生细胞程序性死亡,导致胼胝质酶合成不足,包裹着小孢子的胼胝质不能被降解,四分小孢子无法释放单核小孢子,小孢子无法进一步发育,最终败育。
从本研究观察结果可以看到,绒毡层细胞败育起始早于小孢子败育,说明绒毡层细胞提前发生细胞程序性死亡是导致小孢子败育的重要原因。可育花药发育过程中,绒毡层细胞质电子密度大,含有大量的内质网、线粒体等细胞器;不育花药败育过程中,绒毡层细胞质的细胞器数量明显低于可育花药,细胞内空泡化严重,细胞器少,提前降解,说明不育花药绒毡层细胞无法持续提供小孢子发育所需的物质,因而包裹四分小孢子的胼胝质不能被降解,无法释放单核小孢子,小孢子无法进一步发育进而发生败育。
