珠母贝线粒体全基因组密码子偏好性分析
2023-05-06谭传港喻达辉任童童徐志雄何积翠李素萍白丽蓉
谭传港,喻达辉,任童童,徐志雄,何积翠,李素萍,白丽蓉
(北部湾大学海洋学院,广西北部湾海洋生物多样性养护重点实验室,广西 钦州 535011)
同义密码子为编码相同氨基酸的密码子,其在基因中以不同频率出现的现象称为密码子使用偏好性[1]。突变和自然选择是产生密码子偏好性的主要原因[2-3],一般体现在同义密码子的第3 位密码子上[4]。研究密码子偏好性不仅可反映物种的进化关系,对研究自然选择、遗传漂变及基因重组等现象也有重要意义。mtDNA有独立的遗传体系,可自主复制、转录和翻译,有组成简单、母系遗传、进化速度快等特点[5-6],是细胞核外重要的遗传物质。目前,线粒体基因组密码子使用分析已在瓜螺(Melo melo)[9]、文蛤属(Meretrix)[10]等水生动物中得到充分研究。姜艳[11]通过生物信息学方法对20 种小鲵科线粒体基因组密码子的使用模式进行了聚类分析,为小鲵科物种分子系统发育提供了新思路。段晓克[3]分析了鲌亚科线粒体基因密码子,较好地反映鲌亚科鱼类的进化关系。Uddin等[4]分析了鱼纲、鸟类和哺乳动物线粒体ND1 基因的密码子使用偏倚和系统发育,表明ND1 基因在鱼纲、鸟类和哺乳动物中存在差异。珠母贝(Pinctada margaritifera)隶属于软体动物门瓣鳃纲珍珠贝目珍珠贝科珠母贝属[12],主要分布在台湾及广东以南沿海,印度-西太平洋暖海区[13-15],是唯一生产黑珍珠的珍珠贝,其外壳可用于制作贝雕等工艺品,是重要且稀缺的海洋经济贝类。由于过度捕捞,珠母贝自然资源大幅减少,培育有优良性状的亲本繁殖后代对保护珠母贝种质资源有重要意义,而明确密码子偏好性可有效提高外源基因的表达并优化性状决定基因,加速优良亲本的培育进程[9]。笔者研究珠母贝密码子偏好性,探究珠母贝线粒体基因组密码子组成特征和偏好,为提高外源基因表达效率和系统发育提供理论依据,也为珠母贝种质资源保护提供参考资料。
1 材料和方法
1.1 序列材料
珠母贝线粒体基因组CDS(Coding DNA sequence)序列来源于NCBI 数据库(HM467838.1),线粒体基因组总长15 680 bp,包含34 个基因,其中可编码蛋白的CDS有14条。由于,短序列无法正确计算有效密码子数[16],为降低误差,选取10 条大于300 bp 且起始密码子以ATG 开头的非重复序列用于密码子偏好性分析。
1.2 密码子偏好参数计算
以选择的CDS 为研究对象,利用CodonW 1.4.2软件分析密码子偏好参数,包括密码子的GC 比例(N)、密码子在第1、2、3 位上的GC(GC1、GC2、GC3)比例(N)、同义密码子第3位的GC(GC3S)比例、密码子的偏好性指数(Codon bias index,CBI)、密码子的适应指数(Codon adaption index,CAI)、最优密码子使用频率(Frequency of optimal codon,FOP)、有效密码子数(Effective number of codon,ENC,nENC)、相对同义密码子使用度(Relative Synonymous Codon Usage,RSCU)和总平均亲水性(Grand average of hydrophilia,Gravy)。
1.3 密码子偏好性的影响因素
1.3.1 中性绘图分析 用以初步判断影响密码子使用偏好性的因素。先根据密码子的NGC1和NGC2计算出其平均值NGC12,以NGC12为纵坐标,NGC3为横坐标作散点图,每一个点在图中均表示一个基因。通过分析NGC1、NGC2与NGC3之间的关系,初步判断出密码子使用偏好是受突变影响还是由选择造成,如散点图均在对角线上,则密码子偏好性受突变影响,否则受到选择压力的影响[17-19]。
1.3.2 ENC-plot 绘图分析 使用SPSS 22.0 进行ENC-plot 绘图分析,以进一步确定影响密码子偏好的因素。以NGC3S为横坐标,ENC(nENC)为纵坐标,以坐标定位各个基因,建立散点图,并在图中添加ENC 值的标准曲线[20]。如每个基因均分布在标准曲线附近,则密码子偏好仅受到突变作用的影响;如各基因集中分布在标准曲线下方,则密码子偏好受选择影响。标准曲线方程为:
ENC 比值频数(nENC,Ratio)分布情况可更准确体现ENC 期望值(nENC,exp)与ENC 实际值(nENC,obs)的差异,当nENC,exp与nENC,obs接近时表示基因密码子受突变压力的影响,当两者差别较大时则表示其受自然选择的影响。
1.3.3 对应性分析 是一种利用多元统计探究不同基因密码子变异趋势的方法。利用CodonW 1.4.2软件的RSCU 对应分析功能来分析密码子,从而反映密码子使用偏好性特点[21]。
1.4 最优密码子确定
以ENC 偏好性值的大小对基因进行排序,两端各选出20%基因,分别建立高表达基因库和低表达基因库,再计算两库的同义密码子使用度的差值(ΔRSCU),选择ΔRSCU >0.08的密码子作为高表达密码子,选择RSCU >1.00为高频密码子。若ΔRSCU >0.08且在高表达基因库中的RSCU 值>1,则确定其为最优密码子[22]。
2 结果与分析
2.1 密码子组成分析
珠母贝线粒体基因组是总长15 680 bp 的环状DNA分子,其内包含有34条编码蛋白的CDS(表1)。为减少误差,选择以ATG 为起始密码子、长度大于300 bp 的10 个非重复CDS 序列进行密码子偏好性分析。不同基因密码子不同位置的GC 含量并不相同,总GC 比例为41.1%~47.5%,平均43.4%,第1、2、3 位密码子的GC 比 例(NGC)分别是35.2%~51.8%、35.8%~46.8%、37.7%~51.8%,均值分别为45.6%、40.0%、44.1%;NGC3S为33.60%~48.10%,平均40.90%。NU3S、NG3S分别为44.51%、37.81%,分别大于NA3S、NC3S(23.24%和15.19%),表明珠母贝线粒体基因组主要以U(T)/G 结尾,避免以C 结尾(表2)。CBI为-0.184~-0.040,平均值为-0.105。CAI 为0.119~0.181,平均值为0.151。FOP 为0.259~0.359,平均值为0.322。nENC为39.72~54.35,平均值为47.46,均大于35,说明珠母贝基因组密码子偏性较弱。Gravy为0.216 6~1.038 4,平均值为0.741 2(表2)。

表1 珠母贝线粒体基因组特征Table 1 The characteristics of mitochondrial genome of Pinctada margaritifera

表2 珠母贝线粒体基因组密码子主要参数Table 2 Parameters in mitochondrial genome of Pinctada margaritifera
表3 可见,GC1比例和GC2比例极显著负相关(P<0.01),其相关系数为-0.774,与ENC 呈显著正相关(P<0.05),其相关系数为0.666;GC3比例与GC3S、GC比例为极显著正相关(P<0.01),其相关系数分别为0.985、0.886;GC 比例和GC3S比例极显著正相关(P<0.01),相关系数为0.827。CBI 与FOP极显著正相关(P<0.01),相关系数为0.944。

表3 珠母贝密码子主要参数的相关分析Table 3 Correlation analysis of the parameters of codons in coding sequence in Pinctada margaritifera
表4 可见,26 个密码子的RSCU >1.00。第3 位碱基中,有14个以U结尾,9个以G结尾,3个以A结尾。以U 或G 结尾的密码子为23 个,占全部密码子的88.46%,说明珠母贝线粒体基因组密码子偏爱以U(T)或G结尾。

表4 珠母贝线粒体各氨基酸相对同义密码子使用度(RSCU)Table 4 RSCU analysis on protein coding region in Pinctada margaritifera
2.2 密码子使用偏好性的影响因素
2.2.1 中性绘图分析 图1 表明,NGC12为41.00%~45.35%,NGC3为37.70%~51.80%,NGC12与NGC3的相关系数为0154 5,相关性未达显著水平,回归系数为0.142 9,说明NGC12与NGC3为弱相关。从图1 可见,多数基因主要分布在对角线的下方和上方,少数分布在对角线上,说明自然选择是密码子偏向性形成的主要因素。

图1 珠母贝线粒体基因中性绘图分析Fig.1 Neutrality plot analysis of genes on mitochondrial of Pinctada margaritifera
2.2.2 ENC-plot绘图分析 图2可见,代表基因均位于标准曲线下方。表5 显示,50.00%的基因ENCRatio频数为0.15~0.20,20.00%的基因ENCRatio频数为0.20~0.25。可见,ENCexp与ENCobs频数差异较表明珠母贝线粒体基因组密码子偏好受到突变的影响,但更多受选择的影响。
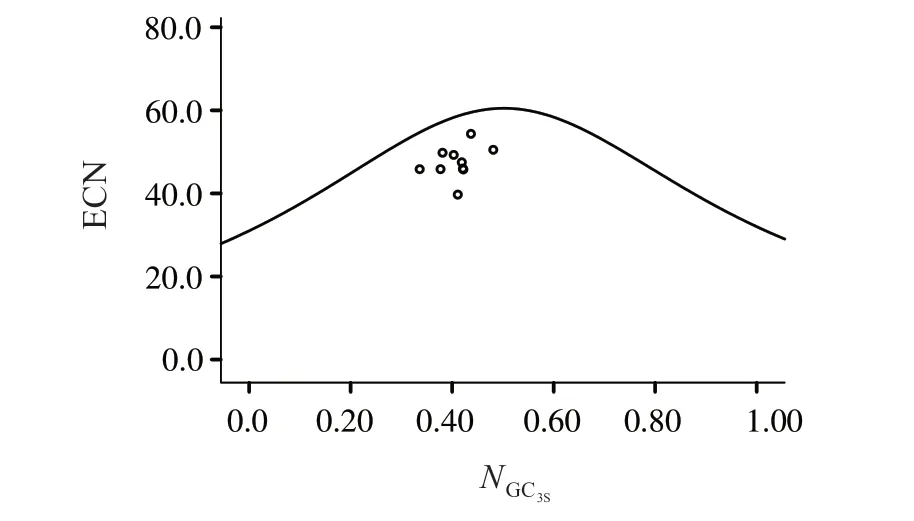
图2 珠母贝线粒体基因密码子ENC-plot绘图分析Fig.2 ENC-plot analysis of codons in mitochondrial genome of Pinctada margaritifera
2.2.3 对应性分析 如图3 所示,第1 向量轴占比为23.43%,第2向量轴占比为14.84%,第2、4轴分别为14.12%和11.47%,4 个向量轴总差异为63.86%,说明第1轴对珠母贝线粒体基因组密码子偏好性造成的影响最大。第1 向量轴与CAI 呈极显著正相关,相关系数为0.895,与GC2比例呈极显著负相关,其相关系数为-0.682,表明碱基差异对珠母贝线粒体基因组密码子偏好性有一定影响。第1 轴与GC1比例、GC 比例、CBI、FOP、ENC 均相关不显著,相关系数分别为0.551、-0.239、0.289、0.363、0.330。以轴1为x轴,轴2 为y轴,建立坐标系。从基因在坐标轴的分布进行分析,基因NAD1、COX2、CYTB分布比较接近,而NAD2、NAD4、ATP6分布比较集中,其他4 个基因则分布比较分散,说明COX1、COX3、NAD6、NAD5与其他基因密码子偏好相差较大。
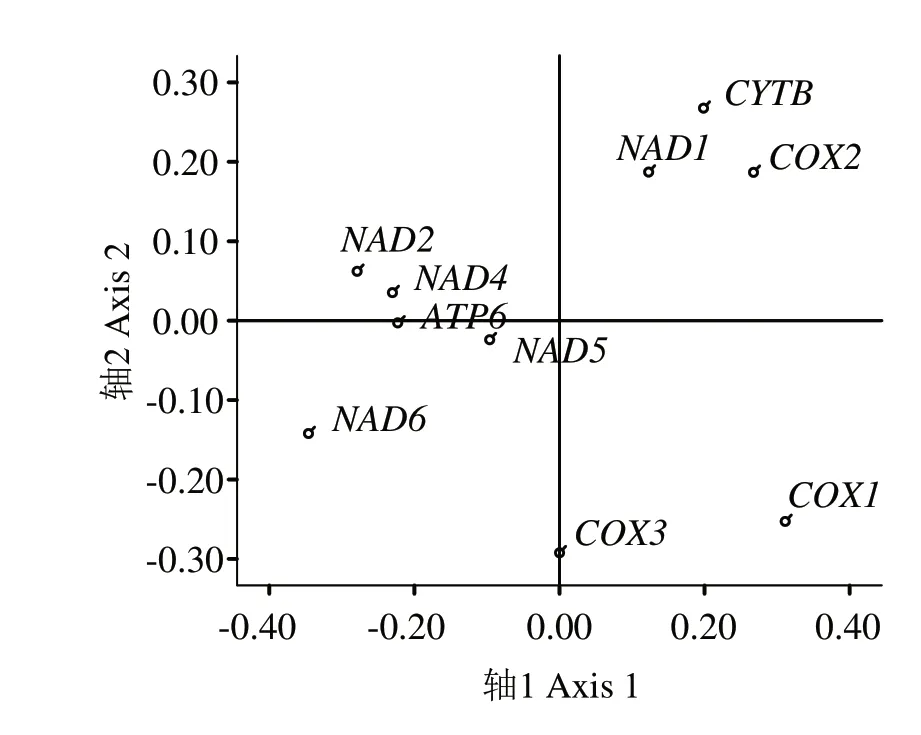
图3 基于RSCU的珠母贝线粒体基因组密码子的对应性分析Fig.3 RSCU-based correspondence analysis of mitochondrial genome codon in Pinctada margaritifera
2.3 最优密码子分析
如表6 所示,将ΔRSCU >0.08 的密码子确定为珠母贝线粒体基因组高表达密码子,最后确定28个密码子为高表达密码子,其中14 个以U 结尾、9 个以G 结尾、3 个以A 结尾。比28 个高表达密码子和26 个高频密码子(表3),将共有的密码子作为最优密码子,最后确定,AGG、AUU、CCU、GAG、GCU、UGA 为最优密码子,其中有2 个密码子以G 结尾,3个密码子以U 结尾,说明最优密码子偏好以U(T)或G 结尾。
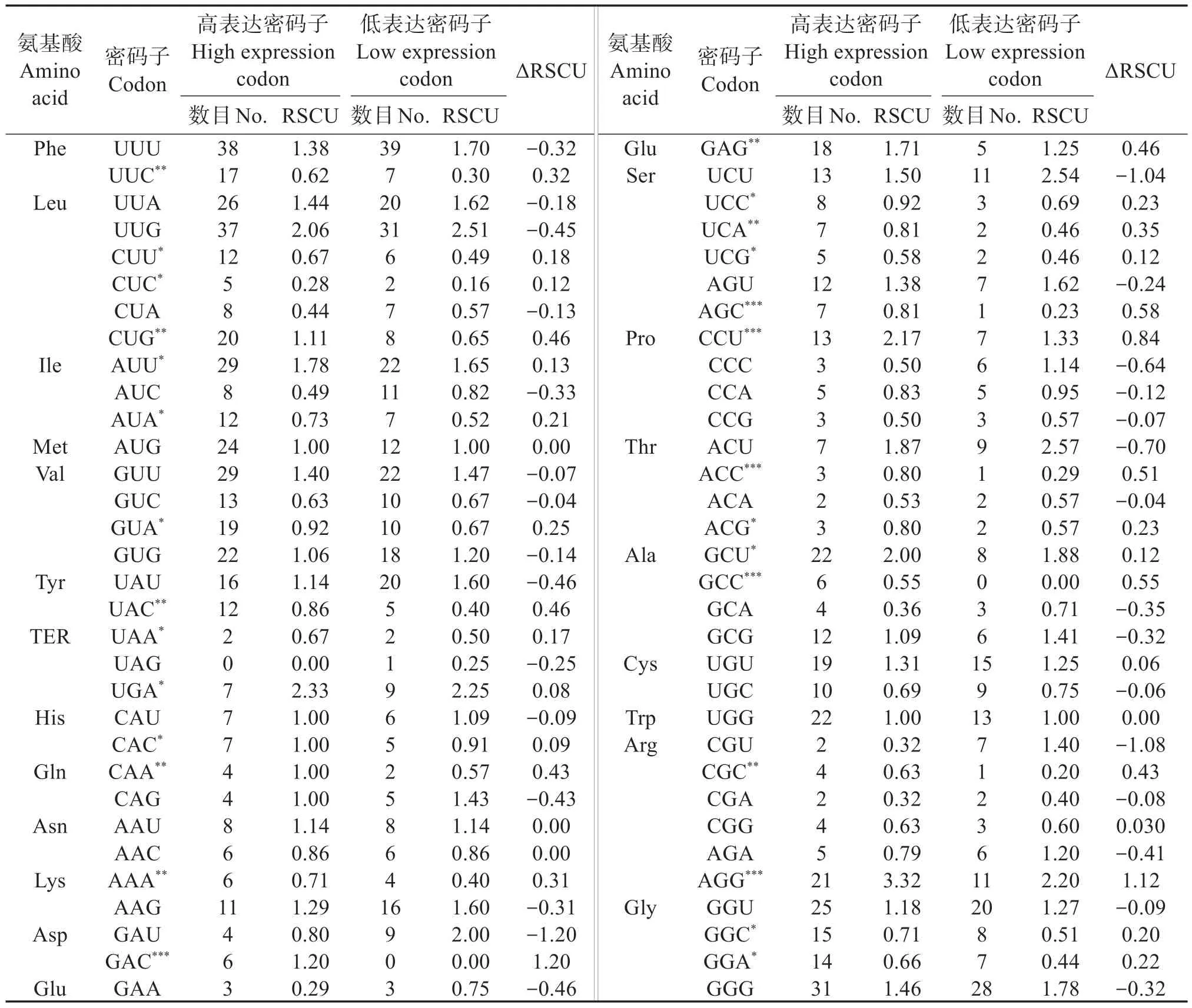
表6 珠母贝线粒体基因组高/低表达基因库的同义密码子相对使用度比较Table 6 Relative Synonymous Codon Usage(RSCU)of genes of mitochondrial genome in Pinctada margaritifera
3 讨论
本研究中,珠母贝mtDNA 编码蛋白基因中,T3S比例为44.51%,G3S比例为37.81%,均大于A3S、C3S比例,可见珠母贝的mtDNA 编码蛋白偏爱以U(T)碱基和G 结尾,避免使用C 结尾。在软体动物中,5种文蛤 属贝类[10]的A3S、G3S、U3S的平均 分别为20.99%、15.09%、59.45%,偏向使用A、U(T)、G结尾,避免使用C 结尾,与珠母贝一致;在鱼类中,斑石鲷(Oplegnathus punctatus)和条石鲷(O.fasciatus)[22]的A3S、C3S的平均比例分别为35.06%、38.60%,更偏向于A、C 结尾;在两栖类中,小鲵科20 种动物[11]均表现为更偏向于T 结尾,其次是A 和C,避免使用G 结尾。表明密码子使用规律在不同物种中有一定差异,而在进化关系较近的物种中,其线粒体基因组密码子使用偏好性相似。珠母贝线粒体基因组ENC 平均值为47.46%,大于标准值35%,表明密码子偏好性较弱。CAI 为0~1,数值越小代表基因密码子适应性越弱,偏向于高表达基因密码子的程度越弱[9],而珠母贝线粒体CAI的平均值为0.151 1,进一步表明珠母贝线粒体基因组密码子为弱偏好性密码子,RSCU 分析结果中,RSCU 值大于1.00 的密码子数目为26 个。其中,第3 位碱基中有14 个以U(T)结尾,9 个以G 结尾;3 个以A 结尾。以U(T)或G结尾的密码子为23 个,占全部密码子的88.46%,进一步说明珠母贝线粒体基因组密码子偏爱以U(T)或G结尾。
影响密码子偏好性的因素较多,主要有自然选择和突变,还有基因序列碱基组成偏好性、tRNA 丰度、GC 含量、基因长度、蛋白疏水性水平以及氨基酸保守性等[23-26]。本研究中,GC12、GC3比例的相关系数为0.393,相关性不显著,说明自然选择在珠母贝mtDNA 密码子偏好性中起主导作用;ENC-plot 绘图中代表基因均位于标准曲线下方,进一步说明珠母贝mtDNA 密码子偏好更多受选择压力的影响,受到突变影响次之。这与斑石鲷和条石鲷[22]、小鲵科20 种动物[11]、5 种文蛤属贝类[10]结果一致。筛选出AGG、AUU、CCU、GAG、GCU、UGA 共6 个密码子为珠母贝线粒体基因组的最优密码子,其中,有3 个密码子以U(T)结尾,2 个密码子以G 结尾,说明珠母贝线粒体蛋白编码基因更偏爱以U(T)/G 结尾的密码子。随着最优密码子的确定还可通过对目的基因的密码子进行优化,提高该基因的表达效率,为未来珠母贝功能基因表达调控研究、蛋白质结构和功能预测等工作提供理论基础,也可为其种质资源保护和人工育苗提供参考。
4 结论
本研究基于生物信息学的方法,从多角度分析珠母贝密码子偏好性,确定AGG、AUU、CCU、GAG、GCU、UGA 共6 个最优密码子,其中有5 个以U(T)或G 结尾,1 个以A 结尾。认为自然选择是造成珠母贝线粒体蛋白编码基因密码子偏好性的主要原因,具体影响机制还有待进一步深入研究。
