珠江口伶仃洋春季鱼类群落结构及其与环境因子的RDA分析
2023-04-12刘文瑜黄建强戴国贤刘国强雷学铁周永东叶永昌
刘文瑜,黄建强,戴国贤,刘国强,雷学铁,周永东,叶永昌*
(1.东莞市自然保护地服务中心,广东东莞 523000; 2.国家海洋局北海海洋环境监测中心站, 广西北海 536000)
河口地处咸淡水交汇处,是淡水生态系统与海洋生态系统间的重要交界面,具有丰富的生物多样性和多样的水文特征,是许多重要海洋、河口和淡水鱼类必不可少的栖息地,它不仅为多种经济鱼类提供索饵、繁殖和育苗的场所,同时也是洄游型鱼类的重要洄游通道[1-3]。珠江口位于广东省中南部,是中国南部最大的河口区,也是南海近海的重要渔场组成部分,鱼类资源丰富[4]。然而,近年来,周边城市经济的快速增长,珠江口的水体富营养化和环境污染等问题日益突出,高强度的渔业生产捕捞使得渔业资源衰退严重,鱼类群落结构也受到严重的影响,河口生态完整性逐渐恶化[5]。
鱼类作为水生态系统的高级消费者,是河口生态系统的重要组成部分,根据下行效应,其群落结构的变化会对水域生态系统的结构和功能产生多方面的影响,在维持生态系统的健康稳定方面发挥着至关重要的作用[6]。诸多研究表明,环境因子是驱动鱼类群落结构变化的重要因素,例如,河口水体较高的浑浊度有助于鱼类躲避捕食者[7];水温不仅影响河口鱼类的洄游,温度梯度变化也会导致鱼类群落产生差异[8-9]。此外,环境及生物因素与鱼类间复杂的交互作用决定了鱼类的聚集行为,且不同的鱼类对环境因素的响应存在区别[10-12]。因此,掌握鱼类群落结构变动及其主要影响因素,对鱼类资源的恢复、鱼类栖息地研究、渔业资源评估及管理政策的制定具有重要作用。
目前,关于珠江口鱼类群落结构与环境因子的相关性研究较少[11,13],仅在相关文献中对鱼类的组成、多样性等有所涉及[4,14-15]。本文基于2021年珠江口伶仃洋的渔业资源调查数据,分别采用相对重要性指数(index of relative importance, IRI)、Shannon-Wiener多样性指数以及Pielou均匀度指数等对该海域的鱼类群落结构进行研究分析,并利用环境数据对本区域鱼类分布进行冗余分析(redundancy analysis,RDA),研究了鱼类与环境因子的相关性。这将为珠江口伶仃洋海域的渔业资源合理开发利用、生物多样性的保护及海域生态修复等提供宝贵的基础资料。
1 材料与方法
1.1 调查方法
2021年3月在珠江口伶仃洋海域小潮期(3月7—8日)和大潮期(3月9—10日)各开展1次渔业资源底拖网调查(网目25 mm,网宽2.8 m)和水质样品采集,共布设10个调查站位(图1)。各站拖速在3~3.5 kn之间,拖时1 h。渔获物鉴定主要参考《珠江鱼类志》[16]和《南海鱼类志》[17],鱼类种名及分类参考Fish Base(http://www.fishbase.org)。水温(T)、pH、盐度(Salt)、溶解氧(DO)、无机氮(TIN)、叶绿素a(Chl.a)等环境因子样品的采集、处理均按照《海洋监测规范》(GB 17378-2007)有关技术要求执行。水样采集根据现场水深决定采样层次,当水深0~10 m时,仅采集表层水样,采集深度为水深0.5 m处;水深超过10 m时,采集表、底层水样,表层为水深0.5 m处,底层为水深距海底0.5 m处。
1.2 数据分析
1.2.1 鱼类生态类型
根据食性(营养结构)特征可将调查鱼类划分为杂食性(omnivore,O)、肉食性(carnivorous , C)、植食性(herbivorous, H)和滤食性(filter feed, F)4类[18];根据垂直空间的分布差异可将调查鱼类划分为中上层(upper-middle, U)、中下层(lower-middle, L)和底层(benthic, B)3类[19]。
1.2.2 群落优势种
采用相对重要性指数(IRI)描述群落生态优势度[20],将IRI值大于1 000的物种定义为优势种,100≤IRI≤1 000的物种定义为常见种,10≤IRI<100的物种定义为一般种,1≤IRI<10的定义为少见种,小于1定义为稀有种。计算公式为:
IRI=(Ni+Wi)×Fi
(1)
式(1)中,Ni是第i种鱼类占渔获总数的个体数量百分比,Wi是第i种鱼类占渔获总量的个体质量百分比,Fi为第i种鱼类出现站数占调查总站数的百分比。
1.2.3 群落多样性
应用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)分析鱼类群落结构多样性特征。分别按以下公式进行计算:
Pi=Ni/N
(2)
(3)
J=H′/log2S
(4)
式(2)~(4)中:Pi为某站位中第i种的个体数占该站位总个数的比例,N表示样品的总个体数,Ni表示第i个物种的个体数量,S为群落中的物种数量。
1.2.4 鱼类群落结构特征及其与环境因子的相关性分析
鱼类多样性统计分析主要使用Primer 5.0软件完成,并使用SPSS 17.0进行大、小潮期鱼类多样性的单因素方差分析(one-way analysis of variance,ANOVA)。利用R version 4.0.5中的vegan包进行不同潮水期各站位的相似性分析(analysis of similarities, ANOSIM)以及鱼类丰度和环境数据冗余分析。分别对环境因子和鱼类丰度数据进行log(x+1)和Hellinger转化。对物种进行趋势对应分析(detrended correspondence analysis,DCA),根据每个轴的梯度长度(lengths of gradient, LGA)选择适宜的排序方法。当LGA﹤3,则进行RDA分析;LGA﹥4,进行典范对应分析(canonical correlation analysis, CCA);3≤LGA≤4,2种方法均可采用。根据DCA分析结果,本研究第一长轴3≤LGA≤4,因此,采用RDA分析。通过envfit函数检验判断环境变量的显著性,进而判断环境因子对鱼类群落的影响。可视化图形主要采用Arc GIS、R和GraphPad 8.0软件制作。
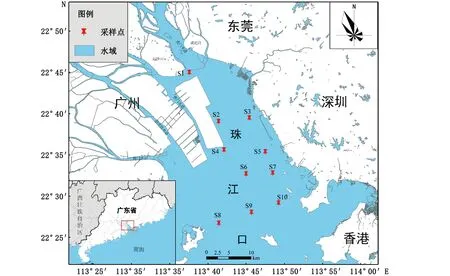
图1 调查地点与采样点分布图Fig.1 Map of the study region and sampling sites
2 结果与分析
2.1 种类组成
本次调查共采集到鱼类2 309尾、19.31 kg,共鉴定出鱼类33种,隶属于13目(系)15科30属(表1)。其中,鲱形目(Clupeiformes)和虾虎鱼目(Gobitiformes)鱼类物种数最多,各有6种,各占鱼类总种数的18.18%;其次为真鲈形系(Eupercaeia),共5种,占鱼类总种数的15.15%;鲽形目(Pleuronectiformes)1科3种,占鱼类总种数的9.09%;其余各目物种数量在1~2种之间,共占鱼类总种数的39.39%。
小潮期共捕获鱼类23种,隶属于10目(系)12科22属,其中有8种鱼类仅在小潮期捕获到,分别为斑鰶(Konosiruspunctatus)、太平洋鲱(Clupeapallasii)、日本小沙丁鱼(Sardinellazunasi)、褐篮子鱼(Siganusfuscescens)、六丝钝尾虾虎鱼(Amblychaeturichthyshexanema)、线纹鳗鲶(Plotosuslineatus)、星点东方鲀(Takifuguniphobles)和鲬(Platycephalusindicus);大潮期共捕获鱼类25种,隶属于12目(系)12科23属,其中有10种鱼类仅在大潮期捕获到,分别为鳊(Parabramispekinensis)、日本发光鲷(Acropomajaponicum)、棘头梅童鱼(Collichthyslucidus)、鳞鳍叫姑鱼(Johniusdistinctus)、叫姑鱼(J.grypotus)、斑尾刺虾虎鱼(Synechogobiusommaturus)、长丝虾虎鱼(Myersinafilifer)、弓斑东方鲀(Takifuguocellatus)、大鳞鮻(Planilizamacrolepis)和长鳍凡鲻(Osteomugilcunnesius);2个潮水期均捕获到的鱼类15种。就渔获物尾数而言,小潮期渔获物尾数为1 449尾,高于大潮期863尾。ANOSIM分析结果显示,大、小潮期(P=0.089)以及不同站点间(P=0.154)的物种组成没有显著差异。

表1 鱼类群落组成及生态类型Tab.1 Fish composition and ecological guilds of the Lingdingyang Estuary
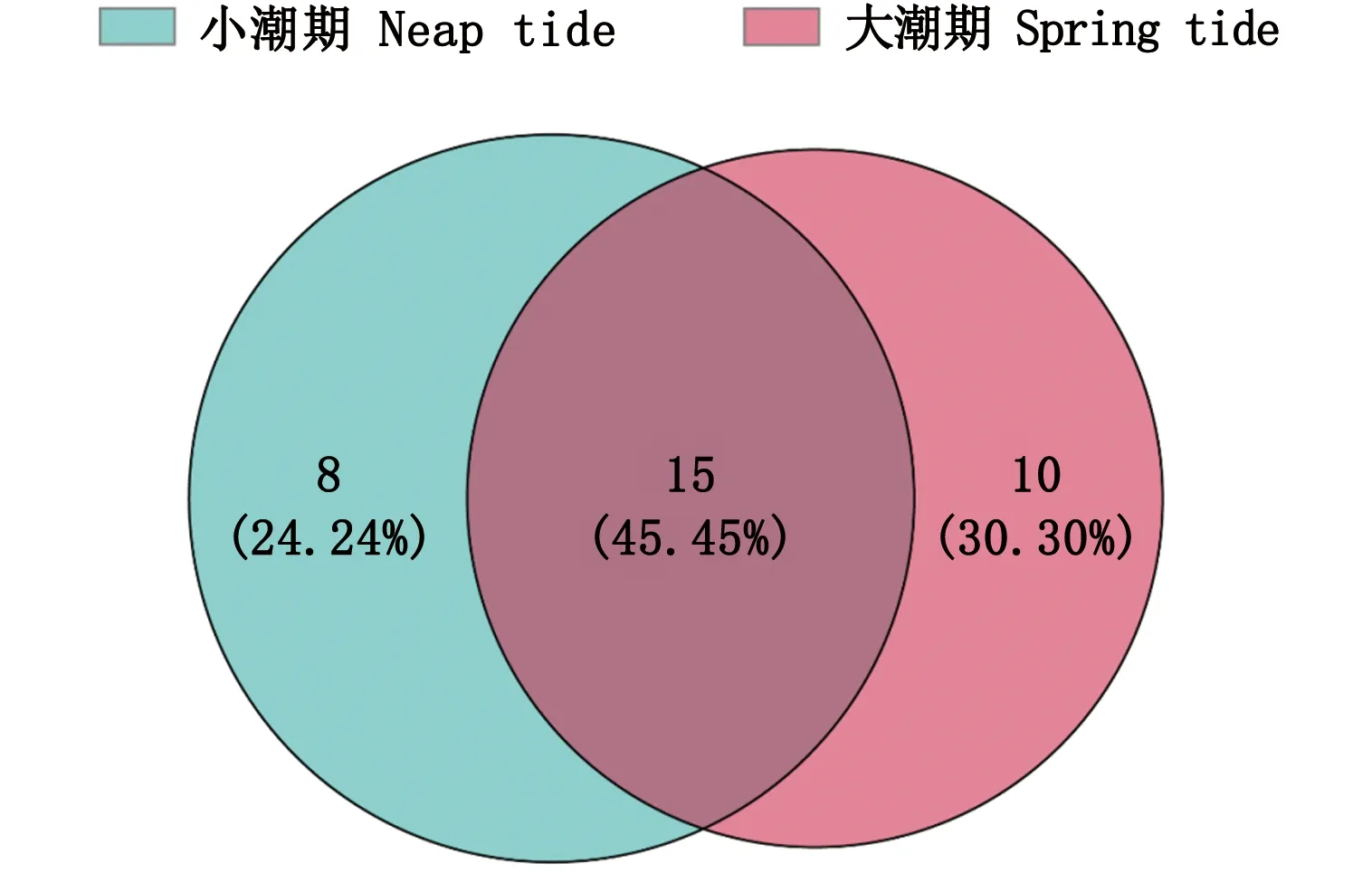
图2 小潮期与大潮期的鱼类种类数Fig.2 Number of fish species in neap tide and spring tide
2.2 优势种组成
根据相对重要性指数统计,大潮期共有优势种3种,其中颈斑鲾(Nuchequulanuchalis)为第一优势种,也是大潮期的独有优势种,总尾数百分比和质量百分比分别为26.35%和27.48%;第二、第三优势种分别为凤鲚(Coiliamystus)和小头栉孔虾虎鱼(Ctenotrypauchenmicrocephalus),分别占总渔获尾数的14.60%和12.70%,占总渔获质量的15.73%和5.62%(表2)。
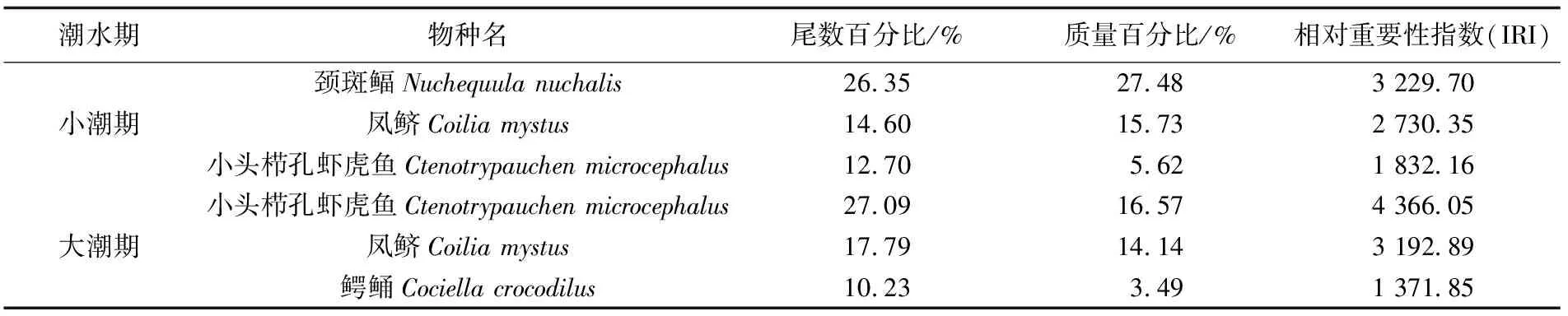
表2 珠江口伶仃洋不同潮水期鱼类优势种组成Tab.2 The dominant species of fish community of the Lingdingyang Estuary in neap tide and spring tide
小潮期优势种也有3种,优势度从高至低依次为小头栉孔虾虎鱼(IRI = 4 366.05)、凤鲚(IRI = 3 192.89)和鳄鲬(Cociellacrocodilus,IRI =1 371.85),总尾数百分比分别为27.09%、17.79%和10.23%,质量百分比分别为16.57%、14.14%和3.49%。其中,鳄鲬为小潮期的独有优势种。
2.3 鱼类群落生态类型
从摄食习性看,调查海域肉食性鱼类居多,共计23种,滤食性和杂食性鱼类各5种,未调查到植食性鱼类;其中,小潮期共调查到肉食性鱼类17种,占小潮期鱼类总种数的73.91%,滤食性鱼类5种,占小潮期鱼类总种数的21.74%,杂食性鱼类1种,占4.35%;大潮期调查到肉食性、滤食性和杂食性鱼类分别为18、3和4种,分别占大潮期鱼类总种数的72.00%、12.00%和16.00%;2个潮水期调查到的肉食性鱼类占比差异不大,但大潮期杂食性鱼类占比略高于小潮期(图3a)。
从栖息水层看,调查海域以底层鱼类为主,共26种,中上层和中下层鱼类相对较少,分别有3和4种;其中,小潮期共调查到底层鱼类17种,占小潮期鱼类总种数的73.91%,中上层鱼类4种,占17.39%,杂食性鱼类2种,占8.7%;大潮期共调查到底层鱼类21种,占大潮期鱼类总种数的84.00%,中上层和中下层鱼类各2种,各占大潮期鱼类总种数的8.00%;大潮期底层鱼类占比大于小潮期,而中上层鱼类占比则低于小潮期(图3b)。
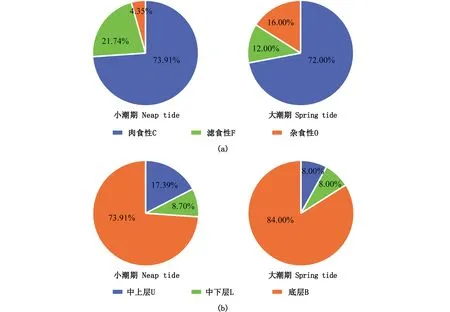
图3 珠江口伶仃洋海域鱼类生态类群分布Fig.3 Distribution of fish ecological guilds of the Lingdingyang Estuary
2.4 鱼类群落多样性
各站点鱼类多样性情况如图4所示。由结果可知,小潮期的Pielou均匀度指数在0.23 ~ 0.98之间,最低和最高值分别出现在S6和S5站点;大潮期Pielou均匀度指数在0.39 ~ 0.86之间,最低和最高值分别出现在S10和S7站点。ANOVA分析结果显示,不同潮水期的均匀度没有显著的差异(P=0.818)。
小潮期Shannon-weiner多样性指数在0.77~2.44之间,最小值出现在S6,最大值出现在S8;大潮期Shannon-weiner多样性指数在1.23 ~ 2.54之间,最小值出现在S10,最大值出现在S3。ANOVA分析结果显示,不同潮水期的多样性指数没有显著差异(P=0.208)。
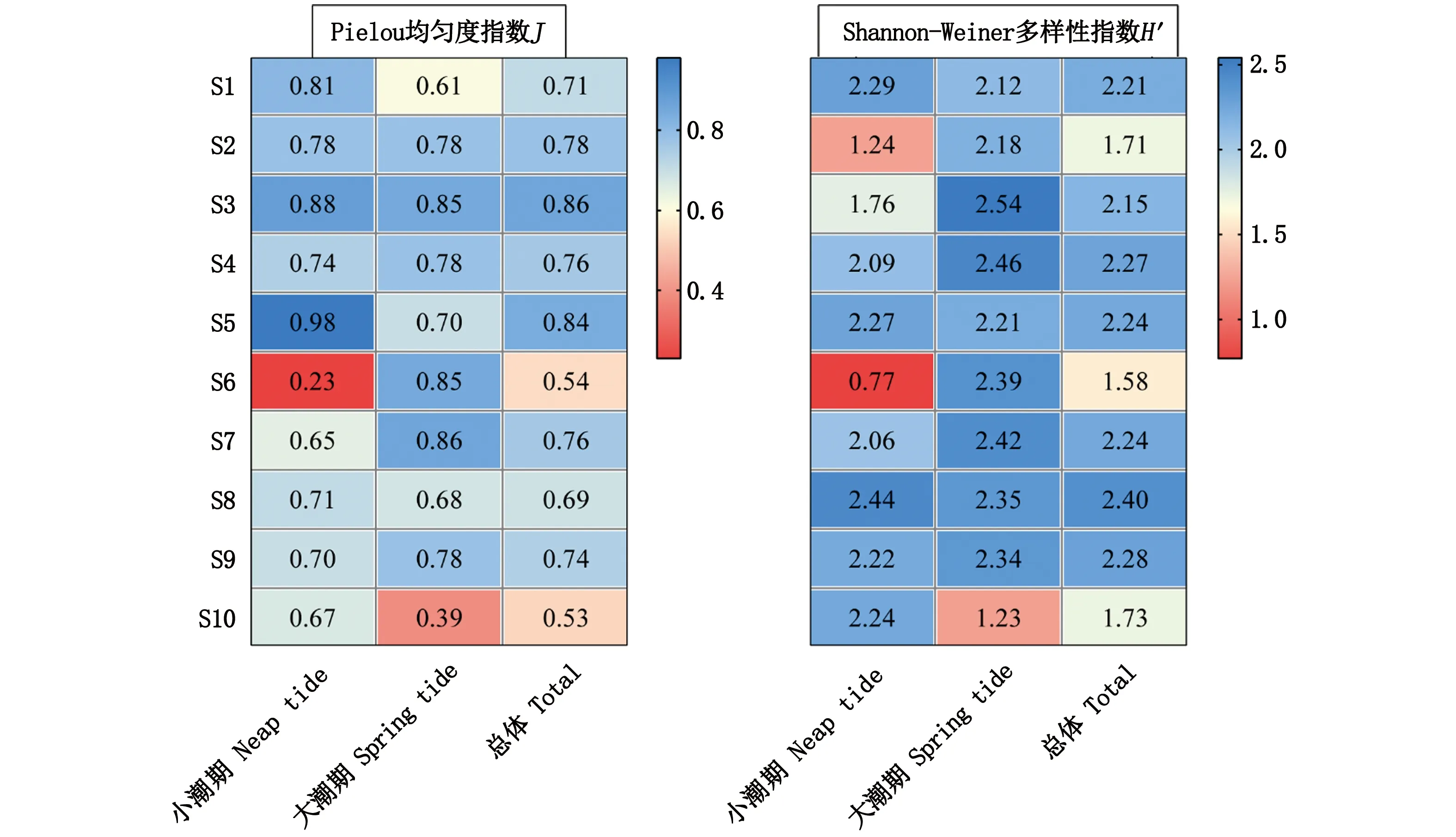
图4 珠江口伶仃洋海域鱼类多样性指数Fig.4 Fish diversity index in the Lingdingyang Estuary
2.5 鱼类群落与环境因子之间的关系
RDA冗余分析结果显示,鱼类群落结构与环境因子总体具有显著的相关性(P=0.05)。其中,水温(P=0.047)、悬浮固体(P=0.007)和叶绿素a(P=0.001)均与鱼类群落显著相关,是影响珠江口伶仃洋鱼类群落的主要环境因子(表3)。颈斑鲾、斑鰶、短吻红舌鳎(Cynoglossusjoyneri)、日本小沙丁鱼等鱼类的分布与水温、悬浮固体和叶绿素a均呈正相关,其他鱼类如小头栉孔虾虎鱼、斑尾刺虾虎鱼、半滑舌鳎(Cynoglossussemilaevis)、日本须鳎(Paraplagusiajaponica)等鱼类分布则与其呈负相关关系(图5)。
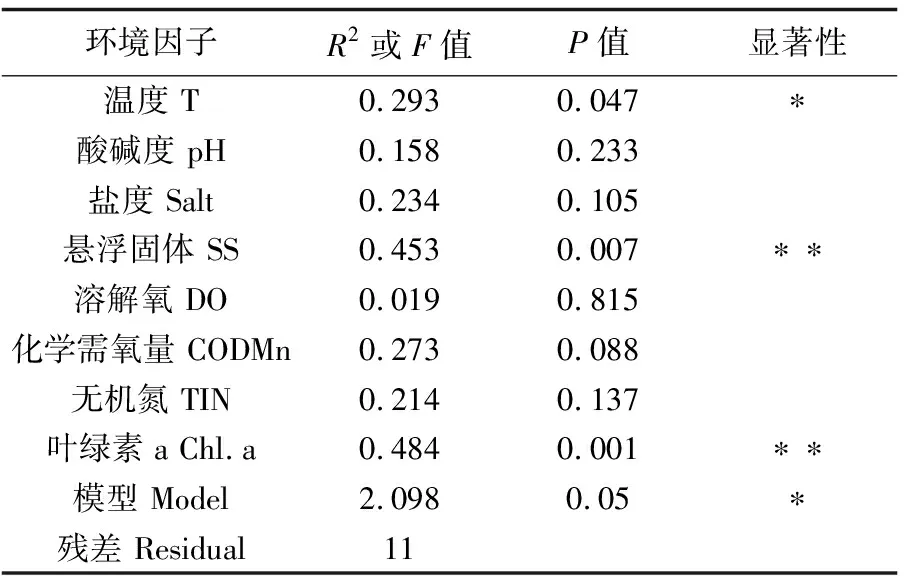
表3 环境因素对鱼类群落的影响Tab.3 Envfit test of environmental factors to fish community
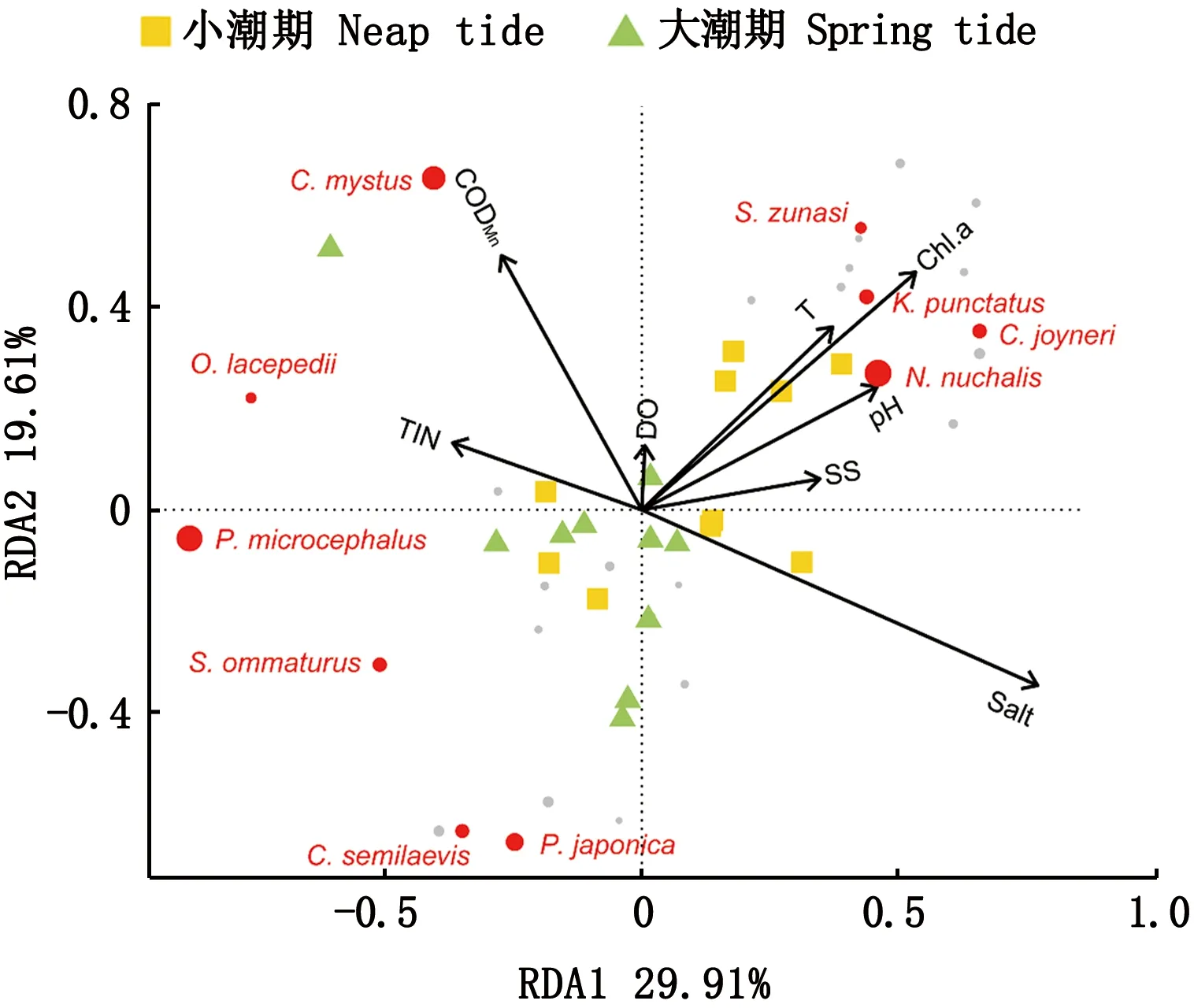
图5 珠江口伶仃洋鱼类组成与环境因子的冗余分析(RDA)Fig.5 RDA analysis between fish composition and environmental factors in the Lingdingyang Estuary注:图中,每个圆点代表一个物种;圆点面积代表物种丰度,面积越大,丰度越高;红色圆点表示丰度为前10的物种Note:In the figure, the circle represents the species, and the size of the circle represents the abundance of the species. Larger circle means higher abundance. Red circles represent the top ten species in abundance
3 讨论
3.1 大小潮对鱼类群落结构的影响
珠江口作为中国三大河口之一,具有“三江汇流、八口入海”的独特自然属性,终年受珠江径流和南海陆架水的综合作用,咸淡水交汇现象明显,水动力复杂多变[21],通常河口潮流和径流的强交互作用对于鱼类的行为迁徙会产生一定的影响。本次调查捕获的鱼类以虾虎鱼目和鲱形目鱼类物种数最多,尤其是虾虎鱼科(Gobiidae)鱼类,从鱼类生态类型上看,底层肉食性鱼类居多,中上层和中下层鱼类相对较少,这与部分学者在同纬度的一般河口区鱼类资源调查结果一致[22-24]。2个潮水期捕获的鱼类在物种数量上无明显差异,但在具体物种上具有一定的不同。如在小潮期仅出现斑鰶、日本小沙丁鱼、褐篮子鱼等滤食性及杂食性小型鱼类,兼有部分如六丝钝尾虾虎鱼、线纹鳗鲶等肉食性鱼类;在大潮期仅捕获到鳊等杂食性小型鱼类和大鳞鮻、长鳍凡鲻等较大型杂食性鱼类,以及日本发光鲷、棘头梅童鱼、鳞鳍叫姑鱼等肉食性鱼类。黄欣[25]在广西南流江河口鱼类群落结构的研究中指出,鱼类群落结构在大小潮间存在显著差异,但物种数在大小潮间差异不显著。此外,潮位的变化会促进水底沉积物的再悬浮[26],有利于底泥中的可溶性无机氮和磷酸盐进入上层水体[11],此时浮游生物大量繁殖,并通过上行效应吸引更高营养级的消费者到来。本次调查结果显示大潮期肉食性鱼类组成相对较高,这可能是由于大潮期带来更多的营养物质,吸引肉食性鱼类到此觅食所致。
相对重要性指数统计结果表明,颈斑鲾为大潮期第一优势种,小头栉孔虾虎鱼为小潮期的第一优势种,而凤鲚为大小潮期的共有第二优势种。黄吉万等[4]在对珠江口的拖网调查中发现,第一优势种为棘头梅童鱼,其次为凤鲚,这与本研究具有一定的相似性,优势种组成均有凤鲚,且二者调查时间均为3月,但调查年份有所不同,随着时间的推移以及受到河口复杂生境的影响,部分优势种的优势度会发生变化,鱼类群落结构较以往发生改变。
3.2 鱼类多样性的时空分布
不同的鱼类由于对诸多环境因子的偏好与耐受性有所不同,从而造成鱼类分布对环境的选择性[27]。本研究发现,S6站点的Pielou均匀度指数和Shannon-Weiner多样性指数受大小潮影响较大,小潮期低于大潮期,提示这个站点的多数种类偏好大潮期的环境;Shannon-Weiner多样性指数除S1、S8、S10站点小潮期高于大潮期外,其他站点均为大潮期高于小潮期。TZENG和WANG[28]对中国台湾淡水河口的研究中发现,涨潮时食物增多,同时鱼类也增多,与其研究相验证的是KRUMME和SAINT-PAUL[29]发现鱼类的丰度、饱食度和食物摄取在大潮期明显高于小潮期,因此,推测这是造成本研究Shannon-Weiner多样性指数大潮期普遍较高的原因之一。
3.3 鱼类群落与环境因子的关系
鱼类群落结构通常受水体中环境变化的影响[27],例如,水温会影响鱼类的洄游及生长发育[8-9],叶绿素a能反映水体中浮游植物的浓度与初级生产力情况,并通过上行效应影响浮游动物、鱼类的聚集与分布[10-11],高浊度环境则有利于众多鱼类躲避捕食者的捕食,从而影响鱼类种群密度[7]。本研究中,水温、悬浮固体和叶绿素a是影响鱼类群落结构的主要环境因子,其中颈斑鲾、斑鰶、短吻红舌鳎的分布与其呈正相关(斑鰶为滤食性鱼类,其分布主要受叶绿素a主导);其他鱼类如小头栉孔虾虎鱼、半滑舌鳎、日本须鳎等分布与其呈负相关(3种均为底层肉食性鱼类,主要分布于河口北部沿岸,主要受水深主导),表明不同鱼类对相同环境因子的偏好有所差异。芮银等[30]在对大陈洋保护区鱼类群落结构的研究中发现,鱼类在盐度、温度和水深等环境维度上存在生态分化现象,棘头梅童鱼和尖头黄鳍牙鱼或(Chrysochiraureus)等受水温影响较大;赤鼻棱鳀(Thryssakammalensis)等近岸中上层小型鱼类受叶绿素a的影响较大;鮸(Miichthysmiiuy)和刺鲳(Psenopsisanomala)等底层鱼类偏好盐度较高的区域;虾虎鱼科等近岸小型底栖鱼类受水深影响较大。短期监测得到的数据可能存在着偏差,河口区受咸淡水交汇及潮位的影响,主导鱼类分布的影响因素较为复杂[10-12, 31],想要得到更为科学可靠的数据还需要进行长期的监测。
4 结论
珠江口伶仃洋大小潮期主要鱼类优势种有颈斑鲾、凤鲚、小头栉孔虾虎鱼、鳄鲬,其中凤鲚和小头栉孔虾虎鱼为大小潮期的共有优势种;时间分布上,大小潮期的鱼类种类组成及其生物多样性均无显著差异,但大潮期捕获的种类略高于小潮期。伶仃洋春季鱼类群落结构与环境因子具有显著的相关性,影响鱼类分布的主要环境因素为水温、悬浮固体和叶绿素a。然而,短期监测得到的数据可能并不能全面反映该海域的鱼类群落结构及其与环境因子相互作用的机制,因此今后需定期对该海域的水环境与鱼类组成进行监测或调查。
