天津海域海月水母分布特征及其与环境因子的关系
2023-04-06张博伦郑德斌徐晓甫曾祥茜
郭 彪,王 硕,张博伦,郑德斌,徐晓甫,曾祥茜
(1.天津市水产研究所,天津,300457;2.天津市海洋牧场技术工程中心,天津,300457)
海月水母(Aurelia aurita)是全球广布种,隶属于钵水母纲(Scyphozoa)、旗口水母目(Semaeoslomeae)、洋须水母科(Ulmaridae)、海月水母属(Aurelia),广泛分布在70°N~40°S 的沿岸海域[1]。近年来,海月水母在我国渤海沿岸频有暴发[2-4]。海月水母作为东亚海域常见的致灾水母种类[5-7],其暴发严重影响海洋生态系统和社会经济发展[4]。一方面,水母摄食浮游动物、鱼卵仔稚鱼,可以改变浮游动物群落结构及生态系统营养结构,水母暴发会影响多个营养级,下行控制初级生产力[8-10]。另一方面,水母暴发会直接破坏渔业捕捞网具[11]、污染养殖设施[12]、危害增养殖生物[13]、造成基础工业设施管道堵塞[14]等。
海月水母生活史复杂,具有有性世代和无性世代的世代交替[2]。性成熟海月水母通过有性生殖释放浮浪幼虫,经一段浮游生活后进行附着,形成螅状体[2,15]。螅状体能够通过出芽、足囊、匍匐茎等多种无性繁殖方式进行螅状体的扩充[2,15]。在适宜的环境条件下,螅状体通过横裂释放营浮游生活的碟状体[2,15]。碟状体在适宜的温度、饵料供给条件下迅速生长发育为性成熟海月水母[2,15]。
天津位于渤海西岸,其近岸海域营养盐丰富,历史上是鱼虾的重要索饵场[16]。近年来,天津海域海月水母持续暴发,渔民生产网具破坏严重,渔民无法进行正常生产作业,许多渔船不得不放弃秋季对虾捕捞生产作业,2021 年天津对虾海捕统计产量为零[17]。目前,关于天津海域海月水母的分布特征未见报道,虽然王朋鹏等[3]于2018 年6 月份调查了整个渤海水母的分布特征,但缺少天津海域的调查站位。
全面了解海月水母在天津海域的分布特征,一方面有助于人们了解天津海域海月水母的暴发源头及其变动规律;另一方面有助于人们制定海蜇等生物的增殖策略,避免海月水母的危害[13],尽可能提高海蜇的增殖效益。为此,本研究在2021 年1—12 月期间系统性调查了天津海域海月水母分布特征,分析海月水母种群分布特征与环境因子的关系,以期为水母防控和海蜇增殖提供科学参考。
1 材料与方法
1.1 调查时间和地点
依据海月水母的生活习性[2,15],2021 年1 月下旬从不同区域(北疆电厂、中心渔港航道、北塘、东疆、临港、南港)海岸线开始向远离海岸线方向推进调查,直至无海月水母碟状体为止;同时在海洋牧场人工鱼礁区设计3 个站位进行调查。2021 年2—12月,每个月下旬安排1 个航次调查海月水母分布特征。每个调查依航次据上个航次调查结果逐步扩大调查范围。1—2 月调查站位图见图1,其他月份调查站位参考结果部分图2 和图3。

图1 初始调查站位示意图
1.2 调查方法
(1)海月水母蝶状体调查:调查采用浅水Ⅰ型网由底到表垂直拖网采集,利用4%(V/V)福尔马林海水固定保存,带回实验室在解剖镜下进行全样品鉴定并计数。2021 年1—4 月调查航次采用该方法。
(2)海月水母成体调查:采用大型浮游生物网在海水表层进行水平拖拽,拖拽船速为2 km·h-1,拖拽时间为10 min,如果拖拽过程中发现浮游生物网网兜内海月水母已满,网兜有严重下沉情况需立即起网,并记录好拖拽时间。2021 年5—12 月调查航次采用该方法。
(3)环境因子调查:水温、盐度、溶解氧、pH 等水质指标采用美国YSI 多参数水质分析仪(型号YSI professional plus)测定;叶绿素a 按照《海洋监测规范》(GB 17378—2007)进行样品采集与测定;同时采用船载LOWRANCE HDS-7 声呐探测仪测定各站位水深。
1.3 数据处理
蝶状体密度按体积计算,其计算公式如下:
式中,Dd 为蝶状体密度;Nd为样品中蝶状体的总个数;V 为垂直拖网水体体积。
由于海月水母成体基本上均漂浮在海水表面,故资源密度采用扫海面积法进行计算。其计算公式如下:
式中,Dc为海月水母成体密度;Nc为采集到的海月水母总数量;S 为采样过程中扫海的总面积。
所获的不同区域和不同时间海月水母资源量数据采用SPSS19.0 进行单因素方差分析,分析结果采用SigmaPlot 软件进行制图,不同站位的资源密度采用ArcGIS 软件作图。运用SPSS 19.0 软件计算了海月水母丰度与海水温度、盐度、溶解氧含量、pH 和叶绿素a 之间的Pearson 相关性系数,并用Origin 2018 软件绘制图形。
2 结果与分析
2.1 天津近岸海域海月水母蝶状体空间分布特征
2021 年1—4 月在天津近岸海域完成4 次海月水母蝶状体调查,其中1 月和2 月未发现海月水母碟状体,3 月和4 月发现海月水母蝶状体,其空间分布特征见图2。

图2 海月水母蝶状体分布示意图
海月水母蝶状体在天津近岸海域出现于3月底,主要分布在近岸线50 m 附近的海域,且岸线须有可供海月水母水螅体附着的礁石或人工岸线;海月水母碟状体的平均伞径长为(4.56±0.63)mm,蝶状体分布海域平均分布密度为(1 063.46±692.14)ind·m-3,电厂附近蝶状体密度最高,远高于其它调查海域(P<0.05)。在远离岸线的海洋牧场人工鱼礁区内未发现海月水母碟状体。
2021 年4 月底,海月水母蝶状体呈数量级增加,分布海域平均分布密度高达(9 165.89±8 758.09)ind·m-3,平均伞经长变化不明显,为(5.56±1.61)mm。蝶状体也由近岸线50 m 附近逐步扩大到近岸线5.556 km 范围。中心渔港附近海域海月水母碟状体的密度显著高于其他调查海域(P<0.05),其他调查海域蝶状体密度没有显著性差异(P>0.05)。
2.2 天津近岸海域海月水母成体空间分布特征
2021 年5—12 月在天津近岸海域完成8 个航次的海月水母成体调查,其中10—12 月调查未发现海月水母成体,其他月份海月水母成体空间分布特征见图3。

图3 海月水母成体分布示意图
2021 年5 月,海月水母继续扩散分布,平均伞径为(2.56±1.67)cm,表层水体平均分布密度为(4.48±3.81)ind·m-3。不同海域海月水母密度差异明显,其中中心渔港附近海域的密度显著高于其他调查海域(P<0.05),北塘和东疆附近海域的密度显著低于其他调查海域(P<0.05)。
2021 年6 月,海月水母在全天津海域均有分布,平均伞径为(8.23±3.18)cm,表层水体平均分布密度为(6.20±1.59)ind·m-3。不同海域海月水母密度差异正在逐步缩小,但中心渔港附近海域的密度仍然显著高于其他调查海域(P<0.05)。
2021 年7 月,受强降雨和各河口排放淡水的影响,天津沿岸海域海水呈明显褐黑色,水体中有大量死亡的藻类。在褐黑色水域中未发现海月水母分布。在远离岸线海域(海水颜色正常)发现海月水母,平均伞径为(13.23±6.11)cm,其表层水体平均分布密度为(8.63±1.75)ind·m-3。各调查海域海月水母的分布密度没有显著性差异(P>0.05)。
2021 年8 月,天津沿岸海域仍存在少量褐黑色海水分布区域,在此区域未发现海月水母。其他区域调查发现海月水母分布均匀,各区域海月水母密度没有显著性差异(P>0.05)。海月水母平均伞径为(14.41±7.77)cm,其表层水体平均分布密度为(8.63±1.36)ind·m-3。
2021 年9 月,较大个体的海月水母消失,海月水母平均伞径为(5.03±2.71)cm。海月水母在天津海域分布均匀,各区域海月水母密度没有显著性差异(P>0.05),其表层水体平均分布密度为(4.26±1.38)ind·m-3。
2.3 天津近岸海域海月水母的时间分布特征
海月水母蝶状体密度在时间序列上波动较大,2021 年3 月底开始出现,4 月底显著增加,4 月份海月水母蝶状体密度显著高于3 月份(P<0.05)(图4)。

图4 海月水母蝶状体丰度变化
将2021 年5—9 月海月水母成体资源密度进单因素方差分析,结果显示7 月和8 月海月水母成体资源密度均值显著高于5 月和9 月(P<0.05)。海月水母成体资源密度的均值在5 月、6 月和9 月之间无显著性差异(P>0.05),6 月、7 月和8 月间无显著性差异(P>0.05)(图5)。

图5 海月水母成体丰度变化
2021 年3—9 月采集到海月水母伞径均值和最大值见图6。由图6 可以看出,2021 年3—9 月海月水母伞径的均值和最大值均随着时间的推移有所增加。进入9 月份,由于大规格海月水母的消失,渔获水母伞径的均值和最大值均出现明显下降。

图6 海月水母伞径变化
2.4 海月水母分布与环境因子的关系
不同调查站位2021 年3 月和4 月海月水母蝶状体密度与环境因子之间的关系如图7 所示。海月水母蝶状体密度与温度、盐度均具有显著相关性(P<0.05),而与pH、溶解氧和叶绿素a 相关性不显著(P>0.05)。其中,蝶状体密度与温度呈显著正相关,相关性系数0.744;蝶状体密度与盐度呈显著负相关,其相关性系数-0.693。

图7 2021 年3—4 月海月水母蝶状体分布密度与环境因子(温度、盐度、pH、溶解氧和叶绿素a)相关性分析结果
不同调查站位2021 年5—9 月海月水母成体密度与环境因子间的关系如图8 所示。海月水母成体体密度与温度呈极显著正相关(P<0.01),其相关性系数为0.595。海月水母成体体密度与叶绿素a 同样呈极显著正相关(P<0.01),其相关性系数为0.506。而海月水母成体体密度与盐度、pH 和溶解氧均没有显著相关性(P>0.05)。
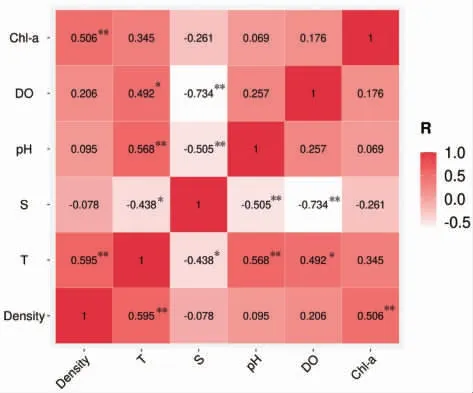
图8 2021 年5—9 月海月水母成体分布密度与环境因子(温度、盐度、pH、溶解氧和叶绿素a)相关性分析结果
3 讨论与结论
3.1 讨论
在自然海域,当环境条件适宜时海月水母螅状体发生横裂释放蝶状体[18],因此蝶状体可用于指示海域是否存在螅状体[4,19]。本研究中,海月水母蝶状体最初的出现海域为近岸50 m 有礁石或人工岸线的海域,其中中心渔港附近海域蝶状体数量较高。这表明,天津近岸海域海月水母螅状体栖息的主要海域为近岸有礁石或人工岸线的海域,其中中心渔港海域为主要栖息地。
2021 年3 月电厂附近海域蝶状体数量最高,其原因可能与该海域平均水深较浅有关。据调查发现,电厂海域的平均水深仅有不足2 m,远低于其他调查海域。较浅水深海域受太阳辐射等影响较大,其水温日变化幅度较大[20],而温度变化,尤其是较高的水温可能诱导海月水母螅状体横裂生殖产生蝶状体[21-22]。
海洋牧场海域存在大量人工鱼礁,理论上其可以为海月水母螅状体提供附着基[23]。但是,笔者的研究中并未发现海月水母蝶状体出现在海洋牧场人工鱼礁区。海洋牧场海域存在大量腹足纲生物[24],这些生物可能会大量摄食海月水母螅状体,抑制了该海域海月水母螅状体的栖息[15,25],需要进一步深入研究。
温度通过调节海月水母机体的酶活性等,实现对海月水母碟状体和水母体生长及代谢的影响[17]。相对温暖水温会刺激海月水母螅状体发生横裂生殖[22],10~17 ℃是胶州湾海月水母产生蝶状体的温度[26]。本研究中,海月水母蝶状体的出现基本符合上述规律,3 月底当水温接近于10 ℃时,海月水母蝶状体开展在近岸海域出现,4 月底当水温升至14 ℃左右时,海月水母蝶状体大量出现。对海月水母蝶状体密度和温度相关性分析也显示,在3 月和4 月期间,海月水母蝶状体密度与温度呈显著正相关,这一结果与其他人的研究结果基本一致[4,21]。本研究中,海月水母成体密度和温度呈极显著正相关,这种相关性与海月水母成体的季节变化趋势相吻合。Lucas等[27]研究发现,海月水母幼体一般在早春时期缓慢生长,春末夏初随着水温的升高呈指数增长,夏末种群数量和个体大小达到最大并停止生长,秋季随着性成熟和浮浪幼虫释放、胃丝排空、生长衰退,个体萎缩变小,随后死亡。本研究中,海月水母密度和伞径的季节性变化规律与其完全一致。
盐度是影响海月水母无性生殖过程的环境因子之一[28]。海月水母螅状体发生横裂生殖产生蝶状体的数量随盐度降低(34~27)而增加[12]。本研究中,虽然3 月和4 月不同区域之间海水盐度变化不大,但是海月水母蝶状体密度仍展现出与盐度显著负相关。这说明水体盐度与蝶状体丰度的相关性很强,薛力园等[4]在研究秦皇岛海域海月水母蝶状体分布与环境因子关系时也发现与本研究相一致的结论。
叶绿素a 能够反应水体中浮游植物的生物量[29],而海月水母的数量会随着浮游植物生物量的升高而增加[27],这可能与浮游植物生物量增加,提高了海月水母食物的可利用率有关[30]。本研究中,海月水母成体密度与叶绿素a 呈极显著正相关,说明浮游植物的生物量影响海月水母的分布。
3.2 结论
综合上述研究结果,总结如下结论和建议:(1)天津近岸海月水母蝶状体的来源地主要是以中心渔港为主的近岸有礁石和人工岸线海域,为本地生长型,其在每年4 月底会集中暴发。(2)海月水母成体种群数量和个体大小从5 月份开始逐步增大,到8月份达到峰值,随后二者开始下降,进入10 月底基本消失。(3)天津近岸海月水母蝶状体与海水温度和盐度显著相关,成体与海水温度和叶绿素a 极显著相关。(4)结合天津近岸海月水母的分布特征,建议海蜇增殖放流尽量在5 月初进行,放流地点应选择距海岸线11.112 km 以上海域,以降低海蜇苗种被海月水母捕食的机率,提高海蜇的存活率。
