广西乐业野生春兰iPBS遗传多样性分析与指纹图谱构建
2023-03-31曾艳华何荆洲龙蔷宇范继征李秀玲卜朝阳
曾艳华,何荆洲,龙蔷宇,范继征,李秀玲,卜朝阳
(广西农业科学院花卉研究所,南宁 530007)
【研究意义】春兰(Cymbidiumgoeringii)是我国传统兰花中栽培历史最悠久、分布最广泛的一个种,具有较高的观赏价值,广西各地山区均有分布。广西乐业县位于桂西北,地处云贵高原东南麓,年均气温16.3 ℃,属亚热带湿润气候区,特别适合兰科植物生长,被称为中国野生兰花之乡,目前已发现有兰科植物52属156种[1],其中兰属植物16种,以春兰品种数量居多。春兰多为异花授粉,频繁的种间杂交产生了广泛的遗传变异。与其他兰科植物相比,春兰在叶形、叶色、花形、花香和花色上具有更丰富的遗传多样性及良好的个体选择机会,但也给其种质分类、鉴定和育种工作带来困难。引物结合位点扩增(iPBS)是以长末端重复序列(LTRs)类反转录转座子保守位点设计引物进行扩增的一种新型分子标记技术[2],与传统的分子标记技术相比,其操作简单有效,无需预先获知相关的LTR序列,在品种鉴别和遗传多样性分析方面更具优势。目前,该技术在国内外已成功应用于石斛兰[3]、金花茶[4]、枸杞[5]、葡萄[6-8]、牡丹[9]、番石榴[10]等多种园艺植物及农作物[11-13]的种质分类、遗传分析和DNA指纹图谱构建。因此,应用iPBS分子标记分析春兰种质的遗传多样性并构建DNA指纹图谱,对广西地区春兰种质的分类、保护及利用具有重要意义。【前人研究进展】分子标记是进行物种遗传研究的有力工具,在种质资源鉴定评价和遗传育种中发挥着重要作用,已广泛应用于兰属植物种质资源研究[14]。近年来,应用于春兰野生资源和栽培品种遗传多样性分析及DNA指纹图谱构建的分子标记有ISSR、SRAP、RAPD、AFLP和ITS,其中以ISSR和SSR分子标记应用最多。已有研究表明,利用SSR分子标记可对不同地域春兰资源进行遗传多样性和群体遗传结构分析[15-17];Huang等[18]开发出15个春兰的SSR分子标记并成功应用于春兰遗传研究;Lee等[19]研究表明,SSR分子标记可对春兰杂交后代进行种间鉴定;彭德镇[20]利用ISSR分子标记分析了江西境内350个野生春兰的遗传多样性,以14条筛选出的引物对样品进行扩增,得到139条谱带,其中多样性条带118条;高丽和杨波[21]利用ISSR分子标记对湖北11个野生春兰居群的遗传多样性水平进行研究,11条引物共检测到127个位点,其中112个为多态位点;孙叶等[22]利用ISSR分子标记分析了96份中、日春兰资源遗传多样性;谢慧敏等[23]利用ISSR分子标记分析江西主要山脉21个春兰野生居群的遗传结构与分化特征,并依据研究结果提出资源保护策略;Hui等[24]利用ISSR分子标记分析春兰栽培品种遗传多样性及亲缘关系。SRAP分子标记在基因组中分布均匀,但易受反应条件和扩增程序变化影响。牛田等[25]利用SRAP分子标记对42个春兰商品种进行遗传多样性分析,筛选出的15对引物组合共扩增出185个位点,其中多态性位点184个,可较好地揭示供试材料的亲缘关系。RAPD分子标记因费用低廉和易于分析,也发展成为传统的DNA分析方法。季祥彪等[26]采用RAPD分子标记分析贵州野生春兰的遗传多样性,结果发现野生春兰具有丰富的遗传变异,同时说明RAPD分子标记可有效用于春兰野生资源遗传多样性评价;孙彩云等[27]等利用RAPD分子标记分析中国兰属的50个材料,结果表明春兰品种的多样性高于其他测试品种,即春兰的遗传多样性整体水平较高;单丽丽等[28]进行春兰基因组DNA提取及对RAPD-PCR反应体系中各影响因素进行优化,建立了适合春兰RAPD分子标记的最佳反应体系。AFLP也是一种检测DNA多态性的分子标记,具有高分辩率、高重复性、高灵敏度和共显性等特点。王晓英等[29]研究证实,AFLP分子标记能有效分析春兰商品种的遗传多样性;陈惠云等[30]利用AFLP分子标记对18种主要名贵春兰品种进行DNA指纹图谱分析,筛选出16对引物,可明确将供试的18个名贵春兰品种加以区分,为保护名贵春兰育种产权、登录新品种、鉴定和检测品种真伪提供技术依据。陈雯[31]基于ITS分子标记分析江西11个野生春兰居群100个春兰种质的遗传结构,揭示了不同地理区域春兰的遗传多样性差异、遗传分化、居群遗传结构和基因流特点。【本研究切入点】迄今,应用iPBS分子标记对野生春兰种质资源进行遗传多样性分析的研究鲜见报道。【拟解决的关键问题】本研究以iPBS分子标记对收集于广西乐业县的81份野生春兰种质和4份春兰栽培品种进行遗传多样性和遗传关系分析评估,构建野生春兰DNA数字指纹图谱,为野生春兰种质资源的分类和早期鉴定评价、种质创新和新品种选育提供参考依据。
1 材料与方法
1.1 试验材料
供试春兰种质共85份(表1),其中,81份野生春兰种质收集于广西乐业县,保存于广西农业科学院花卉研究所兰花种质资源圃;以4份春兰传统商品种(国兰传统名品栽培品种)为对照,分别为余蝴蝶(C.goeringii‘Yu Hu Die’)、宋梅(C.goeringii‘Song Mei’)、笑蝶(C.goeringii‘Xiao Die’)和环球荷鼎(C.goeringii‘Huan Qiu He Din’),均采集于广西农业科学院花卉研究所兰花种质资源圃。取样时选取每份种质3株植株的健康嫩叶,分别用自封袋装好,冰盒包装送至实验室,液氮速冻后置于-80 ℃超低温冰箱保存备用。
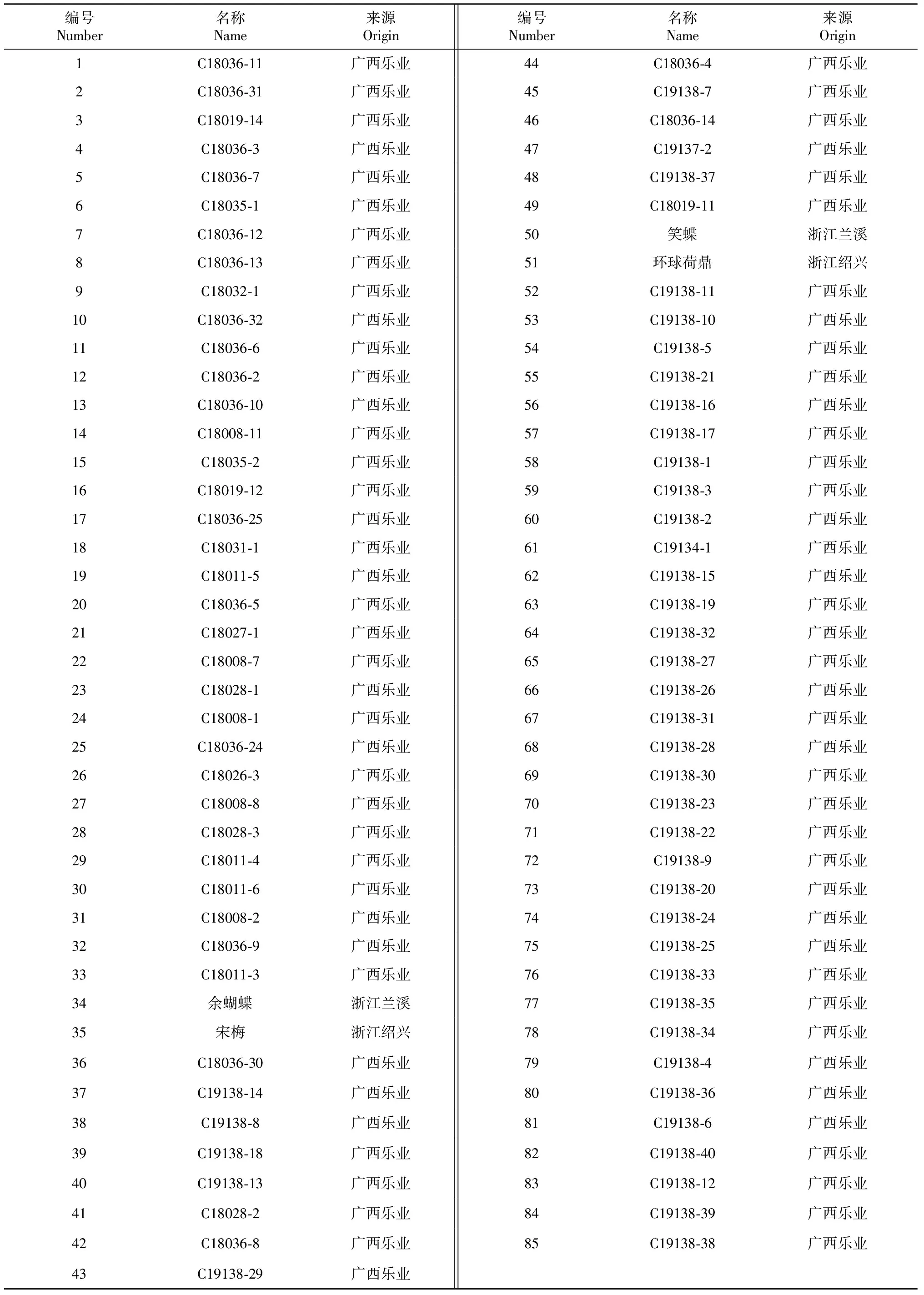
表1 85份春兰种质的名称和来源
1.2 试验方法
1.2.1 试验设计 2015—2019年在广西乐业县陆续收集供试材料,栽培于广西农业科学院花卉研究所兰花种质资源圃,2021年3月选取各性状趋于稳定的每份种质3株,每株采集2枚健康幼叶,分别用封闭的塑料袋包装,保存于冰盒,及时送回实验室液氮速冻,置于-80 ℃超低温冰箱保存备用。
1.2.2 基因组DNA提取 春兰基因组DNA采用Easy Pure Genomic DNA Kit(北京全式金生物技术有限公司)试剂盒提取,每个样本选取3株植株的叶片等量混合,用1.0%琼脂糖凝胶电泳检测DNA完整性,采用紫外分光光度计检测其浓度和纯度,将提取的DNA以TE缓冲液稀释至20.0 ng/μL,置于-20 ℃冰箱保存备用。
1.2.3 引物筛选及iPBS-PCR扩增 采用Kalendar等[2]发表的83个iPBS引物序列对供试材料DNA进行扩增,引物委托生工生物工程(上海)股份有限公司合成。从合成的引物中筛选出扩增条带清晰、多态性高、稳定性强的引物对供试样品进行PCR扩增。PCR反应体系20.0 μL:2×EasyTaqPCR Super Mix 10.0 μL,模板DNA 1.0 μL,引物1.0 μL,无菌水8.0 μL。扩增程序:94 ℃预变性4 min;94 ℃ 1 min,50 ℃(不同引物退火温度见表2)45 s,72 ℃ 1 min,进行35个循环;72 ℃延伸7 min。扩增产物置于4 ℃冰箱保存,取10.0 μL 用1.0%琼脂糖凝胶进行电泳检测。
1.3 统计分析
根据扩增产物的电泳图谱,同一迁移位置上有条带的记为“1”,无条带的记为“0”。以记录的扩增引物条带构建“0,1”矩阵,以NTSYS-pc 2.1进行数据分析,采用UPGMA方法进行聚类并构建树状图。利用POPGEN 1.32计算等位基因数(Na)、有效等位基因数(Ne)、Nei’s遗传多样性指数(He)和Shannon信息多样性指数(I)。根据聚类分析结果,筛选出可鉴别供试材料的引物,根据引物组合的“0,1”矩阵,构建野生春兰的DNA数字指纹图谱。
2 结果与分析
2.1 iPBS引物筛选与种质遗传多态性分析
利用83个iPBS引物序列对85份春兰种质进行DNA扩增,筛选出6条多态性好、重复性高、条带清晰的引物(分别为2081、2076、2225、2251、2249和2232)。由表2可知,6条引物共扩增条带数105条,平均每条引物扩增条带17.5条。其中,多态性条带95条,多态性比率达90.48%;以引物2249的扩增效果最好,扩增的条带数最多(20条),多态性位点比率(PPL)达100.00%。图1为引物2249对样本编号1~40春兰种质的扩增图谱。

M为标准分子量;1~40为春兰种质编号(与表1相同)。
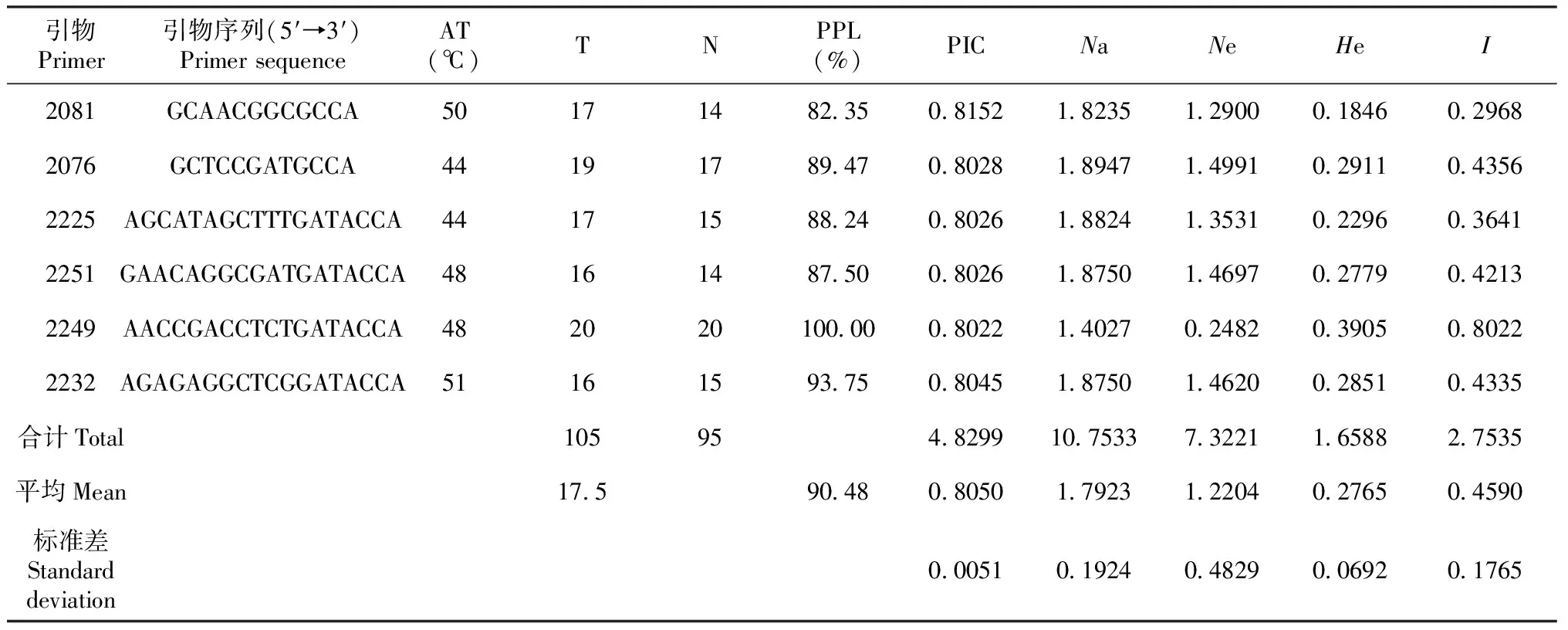
表2 6条引物对85份春兰种质的PCR扩增结果及85份春兰资源的遗传多样性指数
各引物的多态性信息含量(PIC)变辐很小,为0.8022~0.8152,平均为0.8050,标准差为0.0051,说明6条引物在85份春兰种质间均具有较高的多态性;平均Na为1.7923,Ne最高为1.4991,平均为1.2204;平均Nei’s和I分别为0.2765和0.4590,其中,引物2249扩增的多态性位点I最高,达0.8022;各遗传多样性指数均较高,标准偏差均较小,说明供试春兰种质间遗传多样性丰富。
2.2 春兰种质资源间的遗传相似性分析
采用“0,1”矩阵对扩增条带进行遗传相似性分析,得到85份春兰种质资源两两间的遗传相似系数(GS)为0.690~0.981,平均为0.744,供试野生春兰资源在iPBS分子标记的水平上变幅不明显。其中,编号为75号(C19138-25)与74号(C19138-24)的遗传相似系数最大,其次是56号(C19138-16)与57号(C19138-17)及64号(C19138-32)与70号(C19138-23),其遗传相似系数均为0.971。说明这些春兰种质的亲缘关系非常接近,可认定为同一品种或相似品种;亲缘关系较远的是编号为21号(C18027-1)与58号(C19138-1)、35号宋梅与58号(C19138-1)、35号宋梅与54号(C19138-5),其次是16号(C18019-12)与59号(C19138-3)、16号(C18019-12)与60号(C19138-2)、21号(C18027-1)与63号(C19138-19)、35号宋梅与48号(C19138-37)、44号(C18036-4)与85号(C19138-38)及76号(C19138-33)与85号(C19138-38),这6组春兰种质的遗传相似系数均为0.690。
2.3 春兰种质资源的聚类分析
从图2可看出,在遗传相似系数为0.704处,可将85份春兰种质区分为4个类群。其中,第Ⅰ类群包含79份种质,第Ⅱ类群包含4个春兰商品种(编号为34、35、51和50),第Ⅲ和第Ⅳ类群均只有1份种质,分别是21号和36号;4个春兰栽培品种均聚在一起,与其他春兰种质遗传距离相对较远;遗传相似系数较大的75号与74号、56号与57号及64号与70号均聚在一起,而遗传相似系数较小的几组种质均聚到不同的组;除21号和36号种质外,其他广西乐业县野生春兰种质均来源于同一分支,说明21号和36号种质的亲缘关系与其他广西乐业县野生春兰种质相对较远,需进一步探究其遗传分化原因。
2.4 春兰种质资源数字指纹图谱构建
以6条引物对85份春兰种质扩增的条带图谱进行记录,由此构建广西乐业县85份春兰种质“0,1”数字指纹图谱,结果发现引物2249扩增的PPL最高,达100.00%,可区分67份种质;引物2081扩增的PPL为82.35%,可区分41份种质。说明利用引物2249和2076组合可将85份春兰种质资源全部区分出来(表3)。

表3 引物2249和2076构建的85份春兰种质资源DNA指纹图谱
3 讨 论
种质资源评价是资源在作物改良中得以利用的前提条件,通过分子标记辅助确定种质资源的遗传结构,鉴定、管理与重要农艺性状有关的等位基因和种质,可弥补表型评价的不足。随着新一代大规模测序技术的迅速发展,我国兰花的基因组研究取得了突破性进展,完成了多种兰属植物[32-33]的高质量参考基因组测序和组装,为今后功能基因的挖掘与利用打下了基础。植物遗传多样性研究与种质资源收集、保存和更新密切关联,是种质创新和品种改良的基础。通过遗传多样性分析,可检测种质的遗传变异程度和遗传结构,进而评价种质资源的遗传潜力,对种质资源遗传完整性的保存、核心种质筛选及种质资源分类等具有重要参考价值[34],张德全和杨永平[35]对不同类型植物的遗传多样性进行统计,结果显示单子叶植物的I为0.2570,按照生活型划分,草本植物的I为0.2380,按照繁育系统划分,虫媒植物的I为0.2620;高丽和杨波[21]、谢慧敏等[22]研究表明,利用ISSR分子标记分析江西野生春兰和湖北野生春兰的I分别为0.3613和0.4037。本研究中,广西乐业县85份春兰种质的I平均值为0.4590,遗传多样性处于较高水平,与兰科中亲缘关系较近的建兰[36]、墨兰[37]和寒兰[38]相比,其多样性水平仍属最高,说明广西乐业县的生境非常复杂。此外,春兰为多年生草本植物,是兰属的广布种,这也是春兰物种多样性水平较高的主要原因之一,同时说明iPBS分子标记可成功运用于春兰种质资源鉴别及遗传多样性分析,能检测到较ISSR分子标记更高的遗传多态。
iPBS分子标记的每一引物组合检出的多态性条带数量多、重复性好,适用于不同种质间的遗传关系研究和系谱分析。已有研究证实,利用iPBS分子标记分析物种的亲缘关系时,基本都能按地域分布聚类[5-6,39]。本研究结果表明,来源于同一产地的春兰栽培品种均聚在一起,反映出春兰栽培品种与野生种质的遗传差异较大,而4个栽培品种间的遗传差异较小,是长期人为定向选择的结果[40];81份野生春兰种质中有79份具有相同起源,说明广西乐业县不同居群的春兰间变异不明显,总体遗传分化程度很小,其中遗传相似系数较大的种质均聚在一起;两两之间遗传相似系数最大的种质为C19138-25与C19138-24、C19138-16与C19138-17及C19138-32与C19138-23,结合其表型性状来看,两两之间的花色花形非常相似,可判定为近似品种;种质C18027-1和C18036-30的遗传距离与广西乐业种质较远,从花的表型性状来看,C18027-1有卷曲的侧萼片,C18036-30的花瓣质地较厚,花梗更高,花形较独特,可能是由于变异导致遗传分化,但具体原因需进一步研究确认。
4 结 论
iPBS分子标记可用于春兰种质的有效鉴别和遗传多态性分析。81份广西乐业野生春兰种质表现出丰富的遗传多样性,两两间的遗传相似系数变幅不明显,基因交流较频繁,遗传背景较相似;4个春兰栽培品种与81份野生春兰种质间遗传相似系数较小,亲缘关系较远,从分子水平上验证了产于广西乐业的野生春兰与栽培品种在遗传背景上存在差异。
